Full Length Research Paper

ABSTRACT
Cadmium produces a wide range of biological dysfunctions in human and laboratory animals where it chiefly affects the testes, whereas, Carpolobia lutea has been known to have antioxidant benefits. This study was intended to investigate the effects of Carpolobia lutea root extracts on testicular hormone function in cadmium-challenged male rats. 48 male Wistar rats (170-190 g) were divided into six groups, each containing eight rats. Experimental animals in control (Group 1) were given 0.2ml/kg body weight (BW) of 10% tween 80; Group two were administered 1mg/kg BW of cadmium (i.p); Group 3 were given 1mg/kg BW of cadmium (i.p) + 100mg/kg BW extract; Group four took 1mg/kg BW of cadmium (i.p) + 200mg/kg BW extract while Group five and six got 100mg/kg and 200mg/kg BW extract respectively. The administration of vehicle and extract was conducted orally for six weeks. Testicular activity of 17 beta-hydrosteroid dehydrogenase (17β-HSD) and serum testosterone, luteinizing and follicle stimulating hormone (LH, FSH) levels were evaluated. Findings indicated that cadmium statistically (p<0.05) lowered testicular 17β-HSD activity and serum testosterone, LH and FSH levels when compared with those of the control group animals. However, Carpolobia lutea and its co-administration notably (p<0.05) elevated the activity of testicular 17β-HSD and levels of serum testosterone, LH and FSH. The study suggests that Carpolobia lutea extract plays a protective function in ameliorating testicular damage caused by cadmium in rats. This is probably due to the extract’s potential in the management of testicular dysfunction and fecundity in animals.
Key words: Carpolobia lutea, polygalaceae, cadmium, 17 beta-hydrosteroid dehydrogenase, testicular function.
INTRODUCTION
Life is scheduled by nature from the least microbe to the highest whale to have one basic quest, which is to recreate. In addition to other required human activities, reproduction is its basic goal (Taflinger, 1996).
Reproduction is the biological process by which new organisms are produced from their parents. The irony of this is that some people still find it difficult to procreate as a result of infertility. Infertility is defined as the inability of sexually active persons to achieve conception despite regular unprotected sexual activity for a period of one year (Belsey, 1976; Yales et al., 1989; Bonde, 1996; Yesilli et al., 2005). Infertility in men is germane because they determine conception and sex (gender). One of the causes of male infertility is exposure to environmental toxicant. Others are chemotherapy, hypothalamic-pituitary factors, pre-testicular factors, isolated incidences of deficiency in gonadotropin, luteinizing hormone and follicle stimulating hormone as well as androgen excess, glucocorticoid excess and hyperprolactinemia (Reijo et al., 1996; Rosendahl et al., 2010; Olooto et al., 2012).
Cadmium (Cd) is one of the heavy metals and a potent environmental toxicant. Humans are exposed to Cd via contaminants found in drinking water and food; occupational exposition during mining and manufacturing of batteries and pigments that contain Cd; industrial activities such as smelting and refining of metals and municipal waste incineration which release Cd into the atmosphere as well as inhaling tobacco smoke (Blanco et al., 2007; Siu et al., 2009). Cd toxicity is associated with severe damage to testes of both human and animals (Fouad et al., 2009).
Rodent testes are especially sensitive to the deleterious effects of Cd exposure. This is because Cd impairs reproductive capacity by causing severe testicular degeneration, seminiferous tubule damage and necrosis in rats (Burukoglu and Baycu, 2008). The adverse consequences of exposure to Cd on the reproductive organs have been widely considered. Serum levels of testosterone, LH and FSH were notably lowered in the Cd exposed rats (Sadik, 2008).
The reduced activity or privation of male hormones to the testes by Cd has been reported to adversely affect the normal functioning ability of the testes and consequently male fertility (Gupta et al., 2004). Therefore, this growing evidence of toxicity to testes needs to be addressed by screening natural products with claimed androgenic, restorative or ameliorative potentials (Farnsworth, 1989; Eisner, 1990).
Carpolobia lutea G. Don (Family: Polygalaceae), a small tree growing up to 15ft in height (Akpan et al., 2012) is wide spread across the west and central areas of tropical Africa (Mitaine-offer et al., 2002; Nwidu et al., 2015). The common names include cattle stick (English); ikpafum, ndiyan, nyayanga (Ibibio); abekpok ibuhu (Eket); agba, angalagala (Igbo); egbo oshunshun (Yoruba); (Kayode and (Kayode and Omotoyinbo, 2008; Ogunwande et al., 2014). The plant has been scientifically validated to have the following medicinal effects: analgesic (Ajibesin et al., 2008, Jackson et al., 2011), protection of the GIT (Nwidu and Nwafor, 2009), anti-diarrheal (Nwidu et al., 2014), anti-diarrhoeal and treatment of ulcer (Nwafor and Bassey, 2007), anti-microbial (Ettebong and Nwafor, 2009), aphrodisiac (Mitaine-Offer et al., 2002), antimalarial (Okokon et al., 2011), contraceptive (Ettebong et al., 2011), antidiabetic and hypolipidemic (Akpan et al., 2012), acute and sub-acute toxicity (Nwidu et al., 2012), antiacetylcholinesterase activity and antioxidant.
Dare et al. (2015) reported that combined administration of C. lutea methanol root extract at 40 and 80 mg/kg body weight and sildenafil attenuated reproductive function impairment in male rabbit while Yakubu and Jimoh (2015) saw that 47, 94 and 141 mg/kg body weight of C. lutea aqueous root extract reinstated testicular function and sexual capacity in paroxetine-challenged male rats. Despite these streams of fertility findings, there is need to carry out scientific investigation on the male sexual ability of the plant at ethnobotanically acclaimed doses of 100 and 200 mg/kg body weight on toxicity caused by cadmium in rats. Therefore, this study seeks to evaluate the androgenic and testicular effect of methanol extract of C. lutea root on cadmium-challenged rats.
METHODOLOGY
Authentication of plant material
The roots of C. lutea were obtained from Ijare, a village at Akure, Ondo state and authenticated at Forestry Research Institute of Nigeria (FRIN), Ibadan, Nigeria where a Voucher number 109784 was designated to the specimen of C. lutea.
Animal model
Forty eight adult male Wistar rats were obtained from the Animal Care Unit of Igbinedion University, Okada, Edo State. Animals weighed between 170 and 190 g and were housed in a well-ventilated plastic cage and maintained under standard laboratory condition. They ate standard commercial rat pellets and drank clean water daily. Ethical guidelines for animal use were approved by the Institutional Animal Ethics Committee of the Igbinedion University, Okada, Edo State, Nigeria.
Extract preparation
The C. lutea root was air tried and pulverized. 2000 gram of the pulverized powder was soaked in 5 litres of methanol for 48 h and filtered. The filtrate was then concentrated using a freeze-drier. The extract was thereafter dried in a vacuum oven at 40° for 3 h to ensure the removal of leftover solvent. This yielded 50.29 g of semi-solid extract (2.51% yields) was then stored in a refrigerator for the study. The extract was re-diluted in 10% Tween 80.
Experimental design, animal grouping and treatment
The rats were acclimatized for two weeks. There are 6 groups of eight rats each and treated as follows (Table 1). The animals were treated with vehicle, cadmium (i.p single dose) and Carpolobia lutea methanol roots extract orally for 6 weeks. 24 h after the last administration, the animals were sacrificed; blood was collected from the animals via cardiac puncture for hormonal assay while the testes collected was weighed and used for the determination of 17 β-HSD activity.

Determination of serum testosterone
Serum testosterone (17b-hydroxyandrost-4-ene-3-one) level was determined by adopting the procedure described by Chen et al. (1991). Briefly, upon getting the desired number of coated wells, 10 μl of standards, specimens and controls was poured into appropriate wells. 100 μl of testosterone-horseradish peroxidase (Testosterone-HRP) Conjugate Reagent was added into each well. This was followed by topping each well with 50 μl of rabbit anti-Testosterone reagent and thoroughly mixed for 15 s. The mixture was warm at 37°C for 70 min after which the incubation mixture was removed by ficking plate contents into a waste container. The microtiter wells were rinsed and flicked 7 times with deionized water. The wells were then shaken sharply onto absorbent paper to remove all residual water droplets. Thereafter, 100 μl of tetramethylbenzidine (TMB) Reagent was pipetted into each well, mixed for 5 s and kept at room temperature for 15 min. 100 μl of Stop Solution was added to each well to stop the reaction. It was mixed gently for 25 s to allow blue color changes to yellow color completely. Absorbance was then read at 450 nm with a microtiter well reader within 10 min.
Computation of results obtained
The mean optical density value (OD450) was computed for each set of reference standard, control and sample. A standard curve was drawn by plotting the mean absorbance obtained for each reference standard against its concentration in ng/ml on a straight line graph paper; it has absorbance values on the vertical (y) axis, and concentrations on the horizontal (x) axis. Using the mean absorbance values for each sample, the corresponding concentration of testosterone in ng/ml was determined from the calibration curve. Values obtained for diluted samples were further converted by applying the appropriate dilution factor in the calculations.
Principle: The Testosterone ELISA follows the principle that competitive binding between testosterone in the test sample and testosterone-horseradish peroxidase (HRP) conjugates for a constant amount of rabbit anti-Testosterone. During the warming process, goat anti-rabbit IgG-coated wells are warmed with testosterone standards, controls, patient samples, testosterone-HRP conjugate reagent and rabbit anti-testosterone reagent for a period of 60 min. During the incubation, a fixed amount of HRP-labeled testosterone competes with the testosterone in the standard, sample, or quality control serum for a fixed number of binding sites of the specific testosterone antibody. Thus, the quantity of testosterone-HRP immunologically bound to the well gradually decreases as the concentration of testosterone in the rat sample increases. Unbound testosterone-peroxidase conjugate is then removed and the wells washed, followed by addition of TMB Reagent resulting in the development of blue color. The color development is stopped and the absorbance is measured spectrophotometrically at 450 nm. The intensity of the color produced is directly-related to the amount of enzyme present and is conversely-related to the amount of unlabeled testosterone in the sample. A calibration curve is obtained by plotting the concentration of the standard against the absorbance. The testosterone concentration of the test sample and controls run concurrently with the standards can be calculated from the standard curve.
Determination of serum luteinizing hormone
Serum luteinizing hormone was analysed by adopting the procedure described by Nielsen et al. (2001). Briefly, to the coated wells arranged in a rack, 50 μl of standards, test samples, and controls was placed rightly in labelled wells. 100 μl of Enzyme Conjugate Reagent was added into each well, mixed slowly for 20 s and left at room temperature (18-25°C) for 30 min. The incubation mixture was removed by shaking the plate contents into sink. The mixture was cleaned with water and the microtiter wells were stirred 5 times with distilled water. The wells were placed immediately onto paper towels to eliminate all residual water droplets. This was followed by adding 100 μl of tetramethylbenzidine (TMB) Reagent into each well and mixing gently for 10 s. The mixture was left at room temperature for 30 min after which 100 μl Stop Solution was added to each well to stop the reaction. It was slowly mixed for 20 s to ensure that all of the blue color changes completely to yellow color. Absorbance was thereafter read at 450 nm with a microtiter plate reader within 10 min.
Calculation of results
Mean absorbance value (A450) was computed for each set of reference standards, controls and test samples. The corresponding concentration of LH in mlU/ml from the standard curve was determined using the mean absorbance value for each test sample.
Principle: Enzyme immunoassay test follows a typical two-step approach or ‘sandwich’ type assay. The assay utilizes two highly specific monoclonal antibodies: A monoclonal antibody specific for LH is immobilized onto the microwell plate and another monoclonal antibody specific for a different region of LH is conjugated to horse radish peroxidase (HRP). LH from the sample and standards are allowed to bind to the plate, washed, and warmed thereafter with the HRP conjugate. After a second washing step, the enzyme substrate was added. The enzymatic reaction was stopped by addition of the stopping solution. The absorbance was measured on a microtiter plate reader. The intensity of the colour formed by the enzymatic reaction is directly proportional to the concentration of LH in the sample. A set of standards was used to plot a calibration curve from which the amount (concentration) of LH in the test samples and controls can be directly read.
Determination of serum follicle stimulating hormone
Serum follicle stimulating hormone was determined by using the protocol stipulated by Butt et al. (1973). Briefly, the followings were prepared: work solutions of the anti-FSH-HRP conjugate, wash buffer and the required number of well strips. 20 μl of each calibrator, control and test sample was added into respectively marked wells in duplicate followed by addition of 80 μL of assay buffer into each well. This was heated on a plate shaker (exactly 300 rpm) for 20 min at 25°C. The wells were washed 3 times with 300 μl of diluted wash buffer per well. This was followed by adding 90 μL of the conjugate work solution into each well after which it was warmed on a plate shaker (exactly 150 rpm) for 15 min at 25°C and the wells were washed again as earlier done. About 70 μl of TMB substrate was put into each well at 5 min interval. The mixture was heated on a plate shaker for 20 min at 25°C (or until calibrator F attains dark blue color for desired OD). About 30 μl of stop solution was added into each well at 5 min interval after which the plate was spectrophotometrically read on a microplate reader at 450 nm within 10 min after adding of the stop solution.
Calculations
The mean absorbance of each unknown duplicate was computed by minusing the average optical density value of the “0” calibrator from the mean absorbance values of the calibrators, controls and test serum samples. A calibrator curve was drawn on log-log sheet of paper with the average optical densities on the Y-axis and the calibrator concentrations on the X-axis. The unknown values were read directly off the calibrator curve at a standard wavelength.
Principle: The principle follows a typical two-step strategy or ‘sandwich’ type assay. The assay employs two highly potent monoclonal antibodies: a monoclonal antibody specific for FSH is immobilized onto the microplate and another monoclonal antibody active for a different region of FSH is conjugated to horse radish peroxidase (HRP). FSH from the test sample and standards are allowed to bind to the plate, washed, and thereafter warmed at room temperature with the HRP conjugate. After a second washing step, the enzyme substrate was added. Addition of the stop solution terminated the enzymatic reaction. The absorbance was measured on a microtiter plate reader. The degree of the color formed by the enzymatic reaction was directly-related to the concentration of FSH in the test sample. A set of standards was used to plot a calibration curve from which the amount of FSH in rat test samples and controls can be spectrophotometrically read.
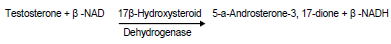
Determination of testicular 17 beta-hydrosteroid dehydrogenase (17β-HSD) [EC 1.1.1.51] activity
Harvested testes were homogenized in 2 ml of normal saline. The homogenate was centrifuged at 1,000 rpm for 15 min. The supernatant collected was employed for the estimation of 17β-HSD. The activity of testicular 17β-HSD was measured according to Jarabak et al. (1996). Briefly, 1 ml of the supernatant was mixed with 1 ml of 440 µmol sodium pyrophosphate buffer (PH 10.2), 40 µl of ethanol containing 0.3 µmol of testosterone (Sigma) and 960 µl of 25% BSA (Sigma), making the incubation mixture a total of 3 ml. The enzyme activity was measured after addition of 1:1 µmol NAD (Sigma) to the tissue supernatant mixture in a U2000 spectrophotometer cuvette at 340 nm against a blank (without NAD). 1 unit of enzyme activity is equivalent to a change in absorbance of 0.001/min at 340 nm.
Principle: The highly specific antibodies pre-coated 96-well microtiter plate containing samples, blanks and standards, incubated with a highly specific biotin-conjugated primary antibody and Avidin conjugated to Horseradish Peroxidase (HRP) and incubated for 10 minutes. After washing the plate and addition of the TMB (3,3’,5,5’-Tetramethylbenzidine) solution, the appearance of a blue colour should be detected due to an enzymatic reaction catalysed by HRP. Aspiration of Stop Solution terminates the HRP reaction causing a colour change from the blue to yellow with the signal intensity read spectrophotometrically at 450 nm on a plate reader. The amount of bound 17-beta-hydroxysteroid dehydrogenase is proportional to the signal generated by the reaction.
Data presentation and analysis
Findings derived from this work were expressed as mean ± SEM of eight determinations. Readings obtained were statistically analyzed using ANOVA and Duncan Multiple Range Test and difference between mean were taken to be substantive at p≤ 0.05. Significant difference was determined by analysis of variance and Duncan Multiple Range Test at 5% confidence level using SPSS 23.0 Software (Statistical Package for Social Sciences, Inc., Chicago, IL, USA).
RESULTS
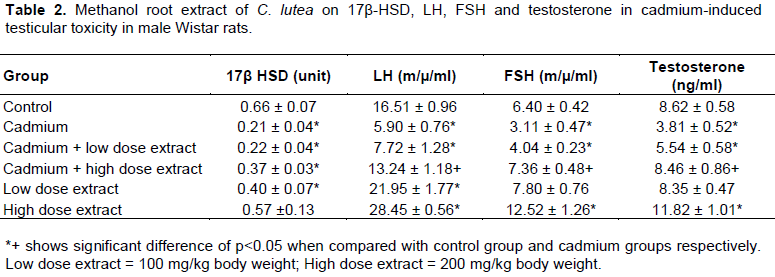
Testicular 17β-HSD activity was substantively (p<0.05) decreased in cadmium when placed with the distilled water treated control group, but was elevated in cadmium plus low dose extract, cadmium plus high dose extract and low dose extract treated rats when matched with cadmium treated animals (Table 2 and Figure 1).
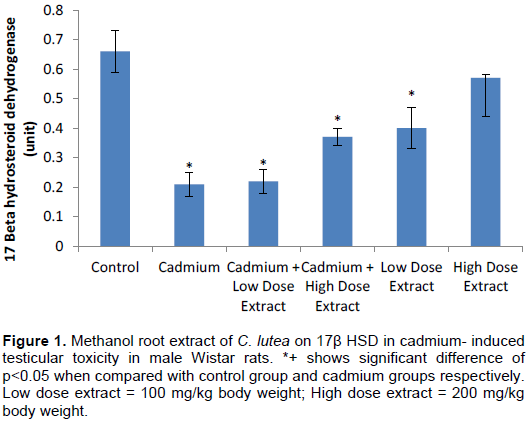
The result showed that LH was significantly (p<0.05) decreased in cadmium, cadmium plus low dose extract treated rats when compared with control group animals. Also, serum LH level was appreciably (p<0.05) elevated in high dose extract and low dose extract treated rats when placed side by side with control group animals (p<0.05). Serum LH concentration was notably (p<0.05) increased in cadmium plus high dose extract treated rats when weighed up with cadmium treated animals (Figure 2).
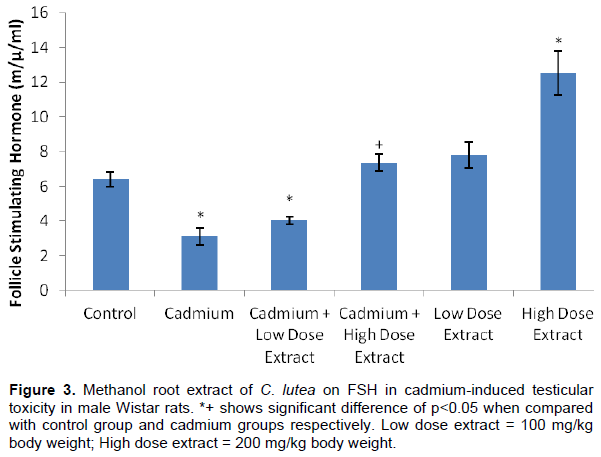
The result showed that serum FSH was sufficiently (p<0.05) decreased in cadmium, cadmium plus low dose treated rats when set side by side with the control group animals. Also, serum FSH was notably (p<0.05) increased in high dose extract treated rats when measured with control groups. Serum FSH was remarkably (p<0.05) increased in high dose extract treated rats when juxtaposed with cadmium treated group (Figure 3).
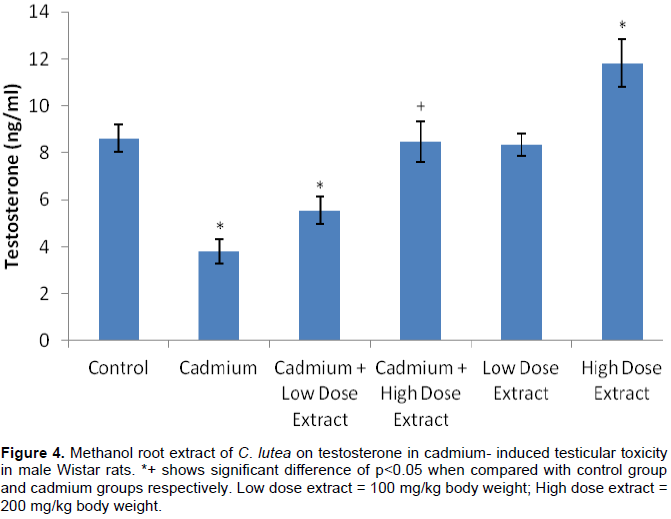
The figure shows that serum testosterone was glaringly (p<0.05) reduced in cadmium, cadmium plus low dose treated rats when liken with the distilled water treated control group animals. Also, serum testosterone was exceptionally (p<0.05) increased in high dose extract treated rats in comparison with rats in the control group (p<0.05). Serum testosterone was meaningfully (p<0.05) decreased in cadmium plus high dose extract treated rats when related with cadmium treated animals (Figure 4).
DISCUSSION
Cadmium produces a wide range of physiological damages in experimental animals and clinical models, affecting mainly the testes (Oteiza et al., 1999). The present study intends to examine the effects of methanol root extract of C. lutea on 17β-HSD, LH, FSH and testosterone in cadmium-induced damage on testes of Wistar rats.
Information gathered from the present work revealed that exposure to cadmium could affect male fertility. The levels of serum testosterone, luteinizing hormone and follicle stimulating hormone were meaningfully lowered in the cadmium treated rats as well as the activity of key androgenic enzyme 17β-hydrosteroid dehydrogenase. This is in agreement with the reports of (Sadik, 2008) who opined that serum testosterone, luteinizing hormone, follicle stimulating hormone, 17β-HSD and antioxidant profile in testes of the animals were significantly decreased as a result of oxidative stress.
Following various doses administered, cadmium notably altered hormonal and enzymatic status, to the extent that it challenged normal testicular function in rats by inducing oxidative stress, damaging testes and altering gonadotropin secretion (Waisberg et al., 2003; Martynowicz et al., 2005). This is in concord with the report of Waisberg et al. (2003) who maintained that via elevated free radical generation, cadmium could cause testicular distortion and impair male sexual function. It is also in agreement with the reports of Neeven et al. (2007); Bench et al. (1999) and Barbara et al. (2008), asserting that cadmium toxicity would alter sexual function, especially by its harmful action on normal male sexuality. Waalkes et al. (1997); and Lafuente et al. (2001) also reported that the decrease in the testicular function in cadmium toxicity could be adduced to its effect at the hypothalamic-pituitary-gonadal axis, suggesting that cadmium affects the function of the axis by acting at these three levels.
Testosterone is a vital biomarker of androgenicity (Walton et al., 1995). It is a major androgen secreted by the testes in nature. Approximately 8 mg of testosterone is generated daily; the major source (95%) is known as the interstitial cells of Leydig (Howell and Shalet, 2001). Testosterone is modified to give DHT by 5a-reductase (Howell and Shalet, 2001). The increase in serum testosterone concentration by methanol extract of C. lutea root at 100 and 200 mg/kg body weight is an indication that the extract has androgenic activity by presumably expediting some committed enzymatic step reactions in the testosterone generative pathway. The rescind in levels of testosterone values by the extract of C. lutea root suggests that the plant may be indicative of the presence of some androgenic substances which might have enhanced the productive architecture of testes of the animals to yield more testosterone (Walton et al., 1995).
Luteinizing hormone (LH) is released from cells in the anterior pituitary called gonadotrophs (Yamada et al., 1994) where it binds to Leydig cells in testes to encourage release of testosterone (Stoleru et al., 1993). The increase in serum LH levels by the extract at 100 and 200 mg/kg body weight may mean a stimulatory action by the extract on hypothalamic-pituitary axis of the male rats. Such elevation in LH level may account for the increased level of testosterone since the gonadotropin is responsible for the synthesis and secretion of testosterone (Pitteloud et al., 2008; Valdes-Socin et al., 2017).
Follicle stimulating hormone (FSH) is essential for gonadal enhancement and maturation at puberty as well as gamete synthesis during meiosis (Simoni and Nieschlag, 1995). FSH has a stimulating effect on the testicles where it is essential for normal spermatogenesis. It is also required for the onset and regulation of spermatogenic process. The significant increase in serum concentration of FSH by the extract at 100 and 200 mg/kg body weight may be attributed to stimulatory effect by the extract on the hypothalamic-pituitary axis or anterior pituitary since the release of FSH into the blood is regulated by the gonadotropic releasing hormone (GnRH) secreted by the hypothalamus (Simoni et al., 1999; Sharma et al., 2012).
17 Beta-Hydroxysteroid dehydrogenase (17β-HSD) catalyzes the enzymatic conversion between highly-active 17 beta-hydroxy- and low-active 17-keto-steroids where it regulates the biological activity of sex hormones (Miettinen et al., 1996). 17β-HSDs catalyzes the reduction of 17-ketosteroids and the dehydrogenation of 17β-hydroxysteroids in steroidogenesis. The major reactions catalyzed by 17β-HSD (e.g., the conversion of androstenedione to testosterone) are hydrogenation (reduction) rather than dehydrogenation (oxidation) reactions (Martel et al., 1992). The increase in activity of 17β-HSD in the testes of the animals by the extract at 100 and 200 mg/kg body weight may be an indication of androgenicity and would boost sexual drive, fertility and reproductive activities in male rats (Labrie et al., 1997).
CONCLUSION
The study shows that cadmium-induced oxidative stress altered testicular and gonadotropin secretion by decreasing androgenic hormones and steroidogenic enzyme activities. C. lutea however ameliorated the altered testicular indices. The study suggests C. lutea as a candidate in the management of penile erectile malfunction and impaired libido especially at 100 and 200 mg/kg body weight.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Ajibesin KK, Ekpo BA, Bala DN, Essien EE, Adesanya SA (2008). Ethnobotanical survey of Akwa Ibom State of Nigeria. Journal of Ethnopharmacology 115(3):387-408. |
|
|
Akpan MM, Okokon JE, Akpan JE (2012). Antidiabetic and hypolipidemic activities of ethanolic leaf extract and fractions of Carpolobia lutea. Molecular and Clinical Pharmacology 3:100-107. |
|
|
Barbara P, Lukaszyk A, Senczuk W (2008). The anti-reproductive effects of long term oral administration of cadmium on the adult male rat. International Journal of Andrology 2(1â€6):150-161. |
|
|
Belsey MA (1976). The epidemiology of infertility: A review with particular reference to sub-Saharan Africa. Bulletin of the World Health Organization 54(3):319. |
|
|
Bench G, Corzett MH, Martinelli R, Balhorn R (1999). Cadmium concentrations in the testes, sperm and spermatids of mice subjected to long-term Cadmium chloride exposure. Cytometry 35:30-36. |
|
|
Blanco A, Moyano R, Vivo J (2007). Quantitative changes in the testicular structure in mice exposed to low doses of cadmium. Environmental Toxicology and Pharmacology 23(1):96-101. |
|
|
Bonde JP (1996). Male fertility. In: Comhaire FM, editor. Chapman and Hall Medicals. New York: Chapman and Hall pp. 266-284. |
|
|
Burukoglu D, Baycu C (2008). Protective effects of zinc on testes of cadmium-treated rats. Bulletin of Environmental Contamination and Toxicology 81(6):521-524. |
|
|
Butt WR, Ryle M, Shirley A (1973). Kinetic studies with 125i-labelled follicle stimulating hormone. Journal of Endocrinology 58(2):275-287. |
|
|
Chen A, Bookstein JJ, Meldrum DR (1991). Diagnosis of a testosterone-secreting adrenal adenoma by selective venous catheterization. Fertility and Sterility 55(6):1202-1203. |
|
|
Dare A, Salami SA, Tolulope OK, Akindele OO, Raji Y (2015). Comparative evaluation of the aphrodisiac efficacy of sildenafil and Carpolobia lutea root extract in male rabbits. Journal of Intercultural Ethnopharmacology 4(4):302. |
|
|
Eisner T (1990). Chemical prospecting: A Call for action. In: Borman, F.E and S.R. Kellert (Eds), Ecology, Economic and Ethics: The Broken Circles. Yale University Press, New Haven, CT. |
|
|
Ettebong EO, Nwafor PA (2009). In vitro antimicrobial activities of extracts of Carpolobia lutea root. Pakistan Journal of Pharmaceutical Sciences 22(3). |
|
|
Ettebong EO, Nwafor PA, Ekpo M, Ajibesin KK (2011). Contraceptive, estrogenic and antiestrogenic potentials of methanolic root extract of Carpolobia lutea in rodents. Pakistan Journal of Pharmaceutical Sciences 24(4):445-449. |
|
|
Farnsworth NR (1989). Screning plants for new medicine. In: Wilson, E.O (Ed.), Biodiversity. PartII. Nattional Academy Press, Washinghton, pp. 88-97. |
|
|
Fouad AA, Qureshi HA, Al-Sultan AI, Yacoubi MT, Ali AA (2009). Protective effect of hemin against cadmium-induced testicular damage in rats. Toxicology 257:153-160. |
|
|
Gupta RS, Kachhawa JB, Chaudhary R (2004). Antifertility effects of methanolic pod extract of Albizzia lebbeck (L.) Benth in male rats. Group 243:6-89. |
|
|
Howell S, Shalet S (2001). Testosterone deficiency and replacement. Hormone Research in Paediatrics 56(Suppl. 1):86-92. |
|
|
Jackson C, Mbagwu H, Jackson I, Ekpe G, Etienam F (2011). Analgesic activities of ethanolic extract of the root of Carpolobia lutea. African Journal of Pharmacy and Pharmacology 5(3):367-370. |
|
|
Jarabak J, Adams JA, Williams-Ashman HG, Talalay P (1996). Purification of a 17β-hydroxysteroid dehydrogenase of human placenta and studies on its transhydrogenase function. Journal of Biological Chemistry 237(2):345-357. |
|
|
Kayode J, Omotoyinbo MA (2008). Cultural erosion and biodiversity: conserving chewing stick knowledge in Ekiti State, Nigeria. African Scientist 9(1):41-51. |
|
|
Labrie F, Luu-The V, Lin SX, Labrie C, Simard J, Breton R, Belanger A (1997). The key role of 17 beta-hydroxysteroid dehydrogenases in sex steroid biology. Steroids 62(1):148-58. |
|
|
Lafuente A, Marquez N, Perez-Lorenzo M, Pazo D, Esquifino AI (2001). Cadmium effects on hypothalamic-pituitary-testicular axis in male rats. Experimental Biology and Medicine 226(6):605-611. |
|
|
Martel C, Rheaume E, Takahashi M, Trudel C, Couet J, Luu-The V, Simard J, Labrie F (1992). Distribution of 17 beta-hydroxysteroid dehydrogenase gene expression and activity in rat and human tissues. The Journal of Steroid Biochemistry and Molecular Biology 41(3-8):597-603. |
|
|
Martynowicz H, Skoczynska A, Karczmarek-Wdowiak B, Andrzejak R (2005). Effect of cadmium on testis function. Medycyna Pracy 56(2):167-174 |
|
|
Miettinen MM, Mustonen MV, Poutanen MH, Isomaa VV, Vihko RK (1996). Human 17 beta-hydroxysteroid dehydrogenase type 1 and type 2 isoenzymes have opposite activities in cultured cells and characteristic cell- and tissue-specific expression. Biochemical Journal 314(3):839-845. |
|
|
Mitaine-Offer AC, Miyamoto T, Khan IA, Delaude C, Lacaille-Dubois MA (2002). Three new triterpene saponins from two species of Carpolobia. Journal of Natural Products 65(4):553-557. |
|
|
Neveen T, Elbetieha A, Darmani H, Khamas W (2007). Effects of long term exposure to cadmium chloride on fertility in adult male mice. Veterinary Research 1(2):40-48. |
|
|
Nielsen MS, Barton SD, Hatasaka HH, Stanford JB (2001). Comparison of several one step home urinary luteinizing hormone detection test kits to OvuQuick. Fertility and Sterility 76(2):384-7. |
|
|
Nwidu LL, Nwafor PA (2009). Gastroprotective effects of leaf extracts of Carpolobia lutea (Polygalaceae) G. Don. in rats. African Journal of Biotechnology 8(1):012-019. |
|
|
Nwidu LL, Nwafor PA, Vilegas W (2015). The aphrodisiac herb Carpolobia: A biopharmacological and phytochemical review. Harmacognosy Reviews 9(18):132. |
|
|
Nwidu LL, Oluwaseyi AS, Nwafor PA (2012). Acute and sub-acute toxicity profile of Carpolobia lutea leaf extract in rats. Journal of Pharmacology and Toxicology 7:140- 149. |
|
|
Nwidu LL, Ukiri OO, Rodrigues CM, Vilegas W (2014). Antidiarrheal mechanism and ionic profile of Carpolobia lutea ethanolic stem-bark extract in rats. African Journal of Traditional, Complementary and Alternative Medicines 11(2):257-263. |
|
|
Ogunwande IA, Flamini G, Avoseh NO, Banwo ID (2014). Essential oil of Carpolobia lutea. Chemistry of Natural Compounds 50(2):373-375. |
|
|
Okokon JE, Effiong IA, Effiong E (2011). In vivo antimalarial activities of ethanolic crude extracts and fractions of leaf and root of Carpolobia lutea. Pakistan Journal of Pharmaceutical Sciences 24(1):57-61. |
|
|
Olooto WE, Amballi AA, Banjo TA (2012). Male infertility. Journal of Microbiology and Biotechnology Research 2(3):379-385. |
|
|
Oteiza PI, Adonaylo VN, Keen CL (1999). Cadmium induced testes oxidative damage in rats can be influenced by Zinc intake. Toxicology 137(1):13-22. |
|
|
Pitteloud N, Dwyer AA, DeCruz S, Lee H, Boepple PA, Crowley WF, Hayes FJ (2008). Inhibition of luteinizing hormone secretion by testosterone in men requires aromatization for its pituitary but not its hypothalamic effects: evidence from the tandem study of normal and gonadotropin-releasing hormone-deficient men. The Journal of Clinical Endocrinology and Metabolism 93(3):784-91. |
|
|
Reijo R, Alagappan RK, Patrizio P, Page DC (1996). Severe oligozoospermia resulting from deletions of azoospermia factor gene on Y-chromosome. Lancet 347(9011):1290-3. |
|
|
Rosendahl MC, Andersen LN, Cour-Freiesleben SG, Juul AK, Andersen LA (2010). Causes of male infertility. Fertility and Sterility 94(1):156-166. |
|
|
Sadik NA (2008). Effects of diallyl sulfide and zinc on testicular steroidogenesis in cadmium-treated male rats. Journal of Biochemical and Molecular Toxicology 22(5):345-353. |
|
|
Sharma TP, Nett TM, Karsch FJ, Phillips DJ, Lee JS, Herkimer C, Padmanabhan V (2012). Neuroendocrine control of FSH secretion: iv. hypothalamic control of pituitary FSH-regulatory proteins and their relationship to changes in FSH synthesis and secretion. Biology of Reproduction 86(6):171. |
|
|
Simoni M, Nieschlag E (1995). FSH in therapy: Physiological basis, new preparations and clinical use. Reproductive Medicine Review 4(3):163-177. |
|
|
Simoni M, Weinbauer GF, Gromoll J, Nieschlag E (1999). Role of FSH in male gonadal function. Annales d'Endocrinologie 60(2). |
|
|
Siu ER, Mruk DD, Porto CS, Cheng CY (2009). Cadmium-induced testicular injury. Toxicology and Applied Pharmacology 238(3):240-249. |
|
|
Stoleru SG, Ennaji A, Cournot A, Spira A (1993). LH pulsatile secretion and testosterone blood levels are influenced by sexual arousal in human males. Psychoneuroendocrinology 18(3):205-218. |
|
|
Taflinger RF (1996). Biological Basis of sex appeal: Human reproduction, cost of sex, reproductive strategies, sexual desire. Springer Edition 2113-6. |
|
|
Valdes-Socin H, Daly AF, Beckers A (2017). Luteinizing Hormone deficiency: historical views and future perspectives. Austin Andrology 2(1):1015. |
|
|
Waalkes MP, Rehm S, Devor DE (1997). The effects of continuous testosterone exposure on spontaneous and cadmium-induced tumors in the male Fischer (F344/NCr) rat: loss of testicular response. Toxicology and Applied Pharmacology 142(1):40-46. |
|
|
Waisberg M, Joseph P, Hale B, Beyersmann D (2003). Molecular and cellular mechanism of cadmium carcinogenesis. Toxicology 192(23):95-117. |
|
|
Walton S, Cunliffe WJ, Kaczkes K, Early AS, McGarrigle HHG, Katz M (1995). Clinical, ultrasound and hormonal markers of androgenicity in Acne vulgaris. British Journal of Dermatology 133(2):249-253. |
|
|
Yakubu MT, Jimoh RO (2015). Aqueous extract of Carpolobia lutea root ameliorates paroxetine-induced anti-androgenic activity in male rats. Middle East Fertility Society Journal 20:192-197 |
|
|
Yales CA, Thomas C, Kovacs GT, De-Krester DM (1989). Androgen, male factor, infertility and IVF. In:Wood C, Trouson A, editors. Clinical in vitro fertilization. USA: Springer-Verlag pp. 95-111. |
|
|
Yamada T, Nakamura J, Murakami M, Okuno Y, Hosokawa S, Matsuo M, Yamada H (1994). The correlation of serum luteinizing hormone levels with induction of Leydig cell tumors in rats by oxolinic acid. Toxicology and Applied Pharmacology 129(1):146-154. |
|
|
Yesilli C, Mungan G, Seckiner I, Akduman B, Acikgoz S, Altan K. (2005). Effect of varicocelectomy on sperm creatine kinase, HspA2 chaperone protein (creatine kinase-M type), LDH, LDH-X, and lipid peroxidation product levels in infertile men with varicocele. Urology 66(3):610-615. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


