Full Length Research Paper

ABSTRACT
Sorghum is a C4 grass native in the semi-arid environments of the African sub-Saharan and consequently chilling stress can affect the performance of the crop, especially at the reproductive stages. Moreover, a significant delay of flowering and maturity was observed when sorghum grows under low temperatures regions, and consequently farmers in highland areas of Uganda face yield penalties. Forty genotypes were evaluated in 2017B and 2018A seasons under non-stress (Kabanyolo) and cold stress (Kachwekano and Zombo) field conditions. Data were recorded on: Days to 50% flowering, days to physiological maturity, culm height, panicle length, panicle weight, kernel weight per panicle, and thousand grain weight. Mean comparison of most agronomic traits recorded indicated high significant differences for season-by-genotype, location-by-genotypes, and the three-way interaction (GxLxS). This indicates that cold stress significantly affects yield components. Significant positive correlation was obtained between days to 50% flowering, days to maturity, and culm height, which suggested that simultaneous improvement of these traits is possible. Some genotypes (IESV 91003LT, IESV 91105LT and IS 29376) were best ranked in normal environment but poorly performed in cold environments, which indicates lack of adaptation in highland. BM6, Cytanobe, IESV 91018, IESV 91609, IS 25563 showed generally good performance and stability in all locations. Therefore, these genotypes can be used as parental lines for further breeding process.
Key words: Sorghum, cold stress, flowering, maturity, yield component.
INTRODUCTION
Sorghum (Sorghum bicolor L. Moench) is among the most important food and animal feed grain crop wordwide and can be considered as the best bioenergy source in this era of global climate change (Reddy et al., 2008), owing to various merits in terms of tolerance to abiotic stresses (Tari et al., 2013). As a C4 grass native in the As a C4 grass native in the semi-arid environments of the African Sub-Saharan regions, the crop is well adapted to hot and dry conditions. However, its gradual introduction into regions characterized by low temperatures has led to the evolution of adapted cold tolerant sorghum (Maulana and Tesso, 2013). Although some progress has been made, numerous abiotic stresses, including cold stress, continue to present challenges in most sorghum producing areas.
Cold stress is a major determinant in the rate of plant growth and development, as well as distribution of plant genotypes in various regions of the planet (Sharma and Solanke and Sharma, 2008; Ramankutty et al., 2008; Yadav, 2009). Sorghum genotypes differ in their growth and development at the threshold temperature of around 15°C (Singh, 1985; Maiti, 1996). Therefore, a genotype can mature faster in non-stress environment, while in cold environment maturity is delayed. This is because gene expression patterns responsible for growth and development are altered under cold environment, and therefore protein stability is compromised which impairs stem and leaf growth (Rymen et al., 2007; Janmohhamadi et al., 2015). Chilling stress was found to cause a significant decline in key cellular functions and photosynthetic activity (Allen and Ort, 2001; Rapacz et al., 2008).
The effect of cold stress on plant growth rate and days to flowering varies among sorghum genotypes (Maulana and Tesso, 2013). However, earliness was found to be affected by both genetic background, environmental conditions, or the interaction of both. Recent studies reported six maturity genes and 40 QTL with small additive effects on flowering time (Rooney and Aydin, 1999; Mace et al., 2013). Hence, development of early maturing sorghum genotypes is a paramount goal for numerous breeding programs due to the fact that harvest can be done before the new season of cold and rainy weather resumes, thereby allowing famers to increase productivity and as well reduce yield penalties.
In Uganda, sorghum is grown in almost all agricultural regions, including the higher altitude regions that cover 25% of the arable land with high population density compared to the national density (Kasozi et al., 2005). To avoid the effects of seasonal cold temperatures, farmers in highland regions of Uganda plant sorghum 4 to 6 weeks before the beginning of the cold period that usually start from February to July. However, farmers are still using unimproved varieties with a longer maturity period of about eight months. Therefore, farmers would benefit from having genotypes with reduced maturity period for production twice per annum in order to alleviate issues of malnutrition and food insecurity. The objective of this study was to evaluate the effect of cold temperatures on plant development, flowering, maturity and yield components of sorghum genotypes in order to identify genetic sources of early maturity under cold stress.
MATERIALS AND METHODS
Genetic plant materials
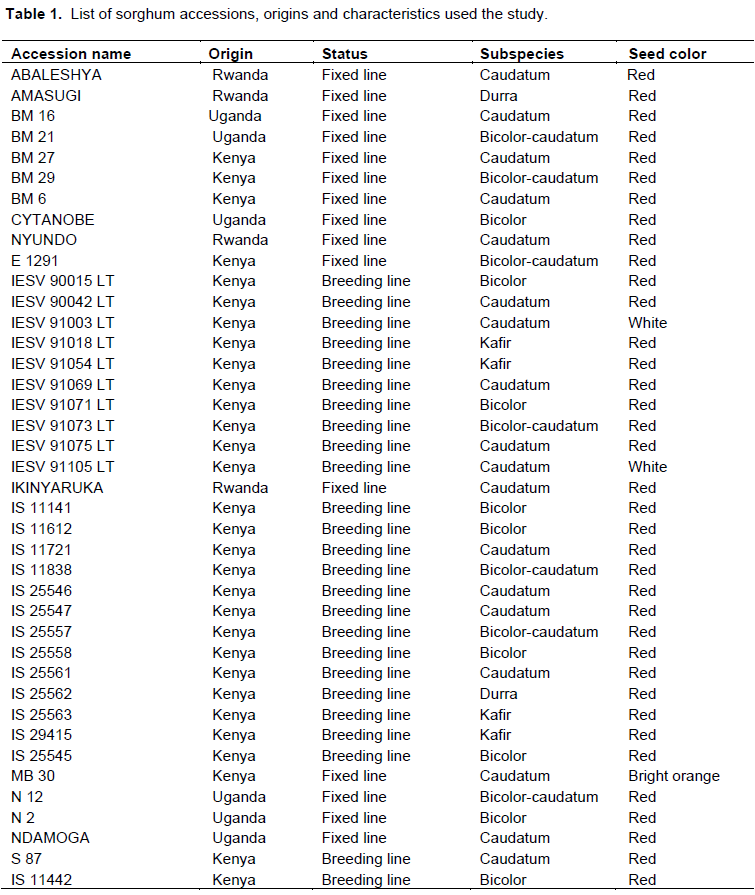
The forty highland sorghum genotypes used in this research study were acquired from International Crops Research Institute for Semi-Arid Tropics (ICRISAT, Nairobi – Kenya), including various breeding lines, released varieties and local landraces. The names, origins and characteristics of sorghum genotype used in the present study are given in Table 1.
Experimental design
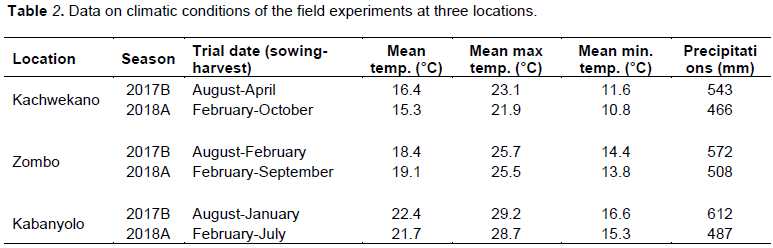
Sorghum genotypes were evaluated during two consecutive seasons (2017B and 2018A) at three locations: Kachwekano field farm (1° 15'S, 29° 57'E, 2,200 m.a.s.l.), located in the highland of South Western region of Uganda, is characterized by a bi–modal rainy season with an annual average rainfall of 1,300 mm, and has a sandy clay loam soil; Zombo (2° 40'S, 30° 54'E; 1,705 m.a.s.l.) situated in the northern region of the country, has heavy clay loam soil with an annual temperature and cooler temperature amplitude; while Kabanyolo (0° 32’N, 32°37’E, 1,240 m.a.s.l) is mid-altitude region characterized by relative optimum temperature ranges (19 - 28°C) for sorghum growth (Table 2).
A 4 × 10 alpha lattice design was used for this experiment with three replications. Plots of 3 m by 2.25 m were laid with spacing of 30 cm within row, and 75 cm between rows. Seeds were planted at 2 cm depth and agronomic practices were applied when necessary. Insecticides (Cypermethrin) were also applied regularly in order to control stem borer and shoot flies.
Data collection and statistical analysis
Data collection included: days to 50% flowering, days to physiological maturity, culm height, panicle length, number of leaves, panicle weight, kernel weight per panicle and thousand kernel weight. Days to 50% flowering were determined as the mean number of days from planting to half-bloom stage. Days to maturity was measured as the average number of days from planting to when the grains on the lower one-third section of the panicles have reached physiological maturity (formed black layer called aleuron). Culm height was recorded as the length of the plant from the ground to the beginning of the panicle, while the panicle length was measured as the length from the beginning to the tip of the panicle. After completion of physiological maturity, panicles were detached, dried and kernel threshed to measuring yield components. Panicle weight was measured as the weight of panicles from individual plants. Kernel weight per panicle was determined as the mean weight of kernels threshed from the individual panicle. A thousand kernel weight was measured and determined from each panicle.
A Restricted Maximum Likelihood (ReML) analysis was used to generate analysis of variance (ANOVA) for single site analysis, using Genstat 18th edition (VSN International, England). For multiple interactions (Genotype x Location x Season), data were analyzed as Randomized Complete Block Design (RCBD) in which replications, locations and seasons were considered as random and genotypes as fixed effects. Means were separated by Fisher’s protected least significance difference at 5% probability level. Pearson correlation was used to determine relationship among traits recorded in this experiment.
RESULTS
Growth and phenological parameters
The pooled analysis of variance including genotype, locations and seasons and their interactions, is presented in Table 3a and b. Genotypes, locations and seasons significantly affected culm height and panicle length. Although, the average reduction of culm height was about 12 cm and 17 cm at Zombo and Kachwekano compared to the optimal growth condition of Kabanyolo, respectively, there was marked variation among sorghum lines. Results showed that late maturing genotypes such as IS 11442, IS 25545, IS 11612 and IS 11721, recorded the tallest culm height (> 250 cm) across environments (Appendix 2). Generally, culm height was greatly affected by cold temperatures, since the non-stress environment of Kabanyolo (Mean: 180.9 cm; range: 100.8 to 323 cm) recorded the higher average compared to Zombo (Mean: 168.5 cm; range: 89.6 to 295.7 cm) and Kachwekano (Mean: 163.3 cm; range: 85.3 to 281.3 cm) (Table 4).
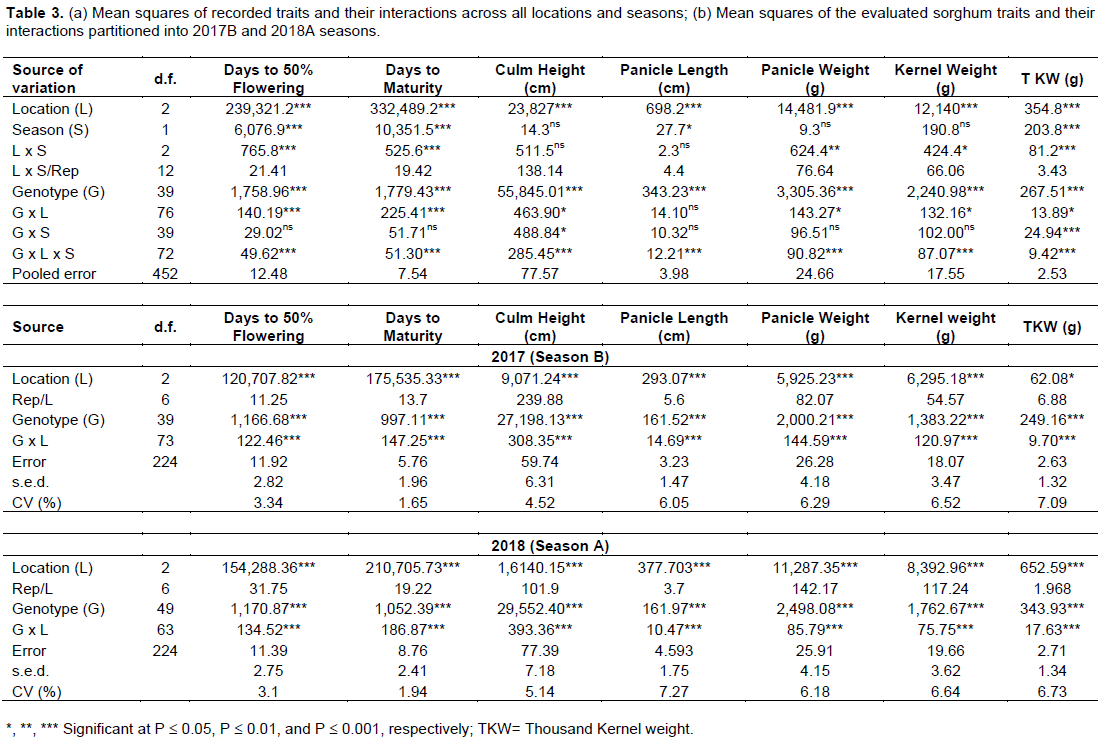
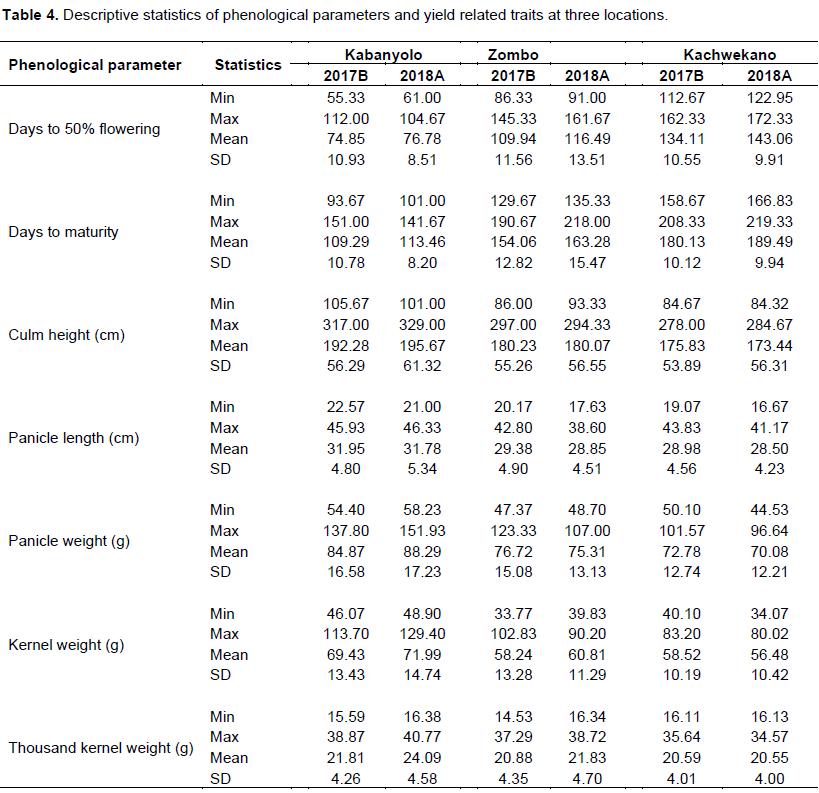
Days to 50% flowering and maturity period were both affected by cold stress, as indicated by the significant interaction of genotype x location x season (Table 3a). As expected, Kachwekano had the longest days to 50% flowering (Mean: 138.5 days; range: 117.8 to 167.3 days) followed by Zombo (Mean: 113.2 days; range: 88.6 to 153.5 days) while the non-stress environment of Kabanyolo recorded the shortest days to flowering (Mean: 75.8 days; range: 58.2 to 108.3 days). A significant genotype x season interaction indicated that flowering took slightly longer in the second season as compared to the first season. A similar trend was observed in days to physiological maturity, since additional 56 days at Zombo, and 73 days at Kachwekano were required to complete this stage, as compared to the non-cold stress environment of Kabanyolo. Generally, genotypes IESV 90015 LT, IESV 90042 LT and IESV 91003 LT flowered earlier than others across environments, while AMASUGI, IS 255545 and IS 11442 matured later (Appendix 2). Moreover, sorghum lines such as IESV 91054 LT, and IS 29415 failed to reach their reproductive stages due to their cold susceptibility under Kachwekano and Zombo environment.
Yield components
As expected, highly significant differences on all yield components evaluated in this study were observed and the genotypes and locations contributed significantly as sources of variation (Table 3a and b). Except at Kabanyolo, the season 2018A recorded relatively inferior yield components values because of extended periods of lower temperatures that occurred from March to August 2018 at Kachwekano and Zombo (Table 4). Overall, the cold weather of Kachwekano reduced 3 to 31.6% across sorghum genotypes, as compared to Kabanyolo. Except IESV 91105 LT that ranked first in the non-cold stress environment (Mean panicle: 144.8 g) and failed to reach maturity in the cold environments of Kachwekano and Zombo, results showed that AMASUGI, BM 6, CYTANOBE and IESV 91018 LT expressed higher panicle weight across environments, however, variation among other sorghum lines were marked.
Thousand kernel weight (TKW) and kernel weight per panicle averaged 25.2 and 73.2 g, respectively, at Kabanyolo, while it decreased at Zombo (TKW: 23.3 g; Kernel weight: 63.2 g) and Kachwekano (TKW: 22.6 g; Kernel weight: 60.8 g). As expected, highest kernel weight per panicle was recorded at MUARIK (IESV 91105 LT: 121.5 g), while the maximum at Zombo and KAZARDI was 102.3 g for IESV 91105 LT and 82.6 g for BM 6, respectively. Although there was marked variation in sorghum lines across locations and seasons (significant genotype x location x season), IESV 91105 LT recorded the highest kernel weight per panicle (Mean: 121.5 g), while the maximum at Zombo and Kachwekano was 102.3 g for IESV 91105 LT and 82.6 g for BM 6, respectively. Although there was marked variation in sorghum lines across locations and seasons (significant genotype x location x season), IESV 91003 LT and IESV 91105 LT expressed a higher TKW but were partially tolerant to cold, since they were unable to survive the weather conditions of Kachwekano in the season B. However, three sorghum genotypes recorded the lowest TKW, less than 17 g, at Kachwekano (BM21, IESV 91071 LT, IS 25561), Zombo were (BM16, IS 11721, IS 29376), while AMASUGI, IS 11612, and IS 11721 were ranked as the last at Kabanyolo.
Relationship among observed traits in the field trials
At Kachwekano, days to flowering was positive and highly significantly correlated to days to maturity (r = 0.95), culm height (r = 0.63), but negatively significant correlated to thousand kernel weight (r = -0.57) and slightly correlated to panicle weight (r = -0.29) (Table 5). Days to maturity was also highly significant correlated with culm height (r = 0.65) and thousand kernel weight (r = -0.54), and slightly correlated with panicle weight (r = -0.29) but non-significant with panicle length (r = 0.12) and kernel weight (r = -0.29). Moreover, panicle weight was also highly correlated to kernel weight (r = 0.96).
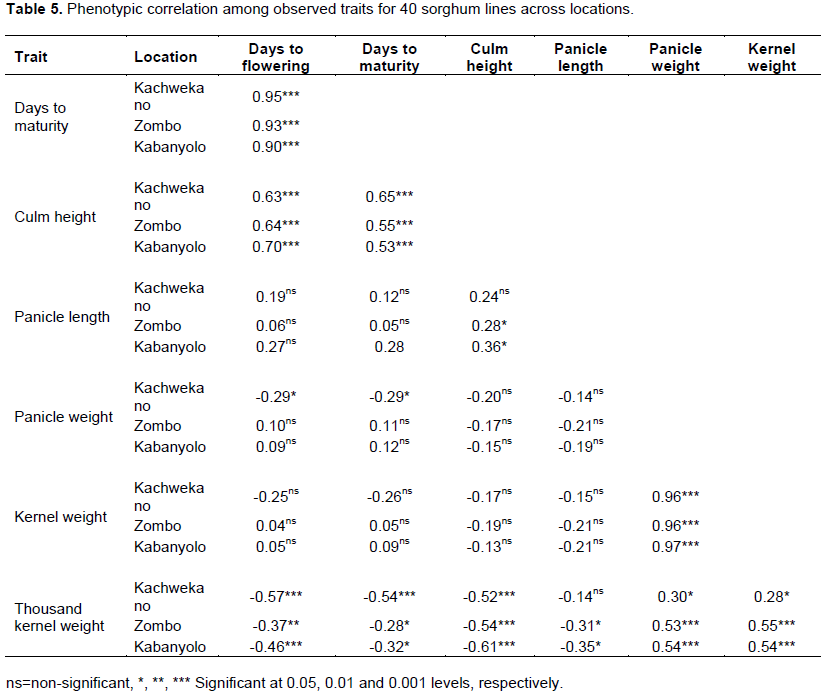
A similar trend was observed at both Kabanyolo and Zombo, where days to flowering was significantly correlated to days to maturity (r = 0.90 and r = 0.93, respectively), culm height (Kabanyolo: r = 0.53; Zombo: r = 0.56) and thousand kernel weight (Kabanyolo: r = -0.46, Zombo: r = -0.37). Days to maturity was significantly correlated to culm height (Kabanyolo: r = 0.53, Zombo: r = 0.56). Panicle length was only significant correlated to culm height (Zombo: r = 0.28; Kabanyolo: r= 0.36) and thousand kernel weight (Kabanyolo: r = -0.35; Zombo: r = -0.31).
Panicle weight was highly significant correlated to kernel weight (Zombo: r = 0.96; Kabanyolo: r= 0.97), and thousand kernel weight (Zombo: r = 0.53; Kabanyolo: r = 0.54), but non-significant to panicle length (Table 5). Moreover, kernel weight per panicle was significantly correlated to thousand kernel weight at Kabanyolo (r = 0.54) and Zombo (r = 0.55).
DISCUSSION
Effect of cold stress to flowering time
Being a C4 plant native in the tropical regions, sorghum is sensitive to temperature below 15°C at all growth and developmental stages (Solanke and Sharma, 2008). Delays in both flowering time and days to physiological maturity are the most frequent phenomena found in cool weather environments (Kapanigowda et al., 2013), especially in the African highland regions. In the present study, we noted that sorghum grown under cool weather (Kachwekano and Zombo) delayed significantly to reach days to 50% flowering and physiological maturity, even for the cold tolerant lines. This is because cold stress acts on key cellular functions, metabolism and photosynthetic activity (Rymen et al., 2007; Zhu et al., 2007; Liu et al., 2019). Therefore, plants responded by slowing the growth rate during the vegetative stage, except for susceptible sorghum lines that died at early developmental stages.
Flowering time and physiological maturity are characteristics controlled by the genetic make-up of the plant and other environmental factors, especially temperatures (Andres and Coupland, 2012). Therefore, different sorghum genotypes can show variable responses under different temperature regime. Since the beginning of 20th century, maturity has been an important trait and one of the main focus for sorghum breeding programs (Quinby et al., 1974). This focus is because knowledge about genetic mechanism that regulate flowering and environmental factors that affect this trait (Murphy et al., 2011), especially temperatures that are responsible for the plasticity in different environments (Marais et al., 2013), could play an important role in the optimization of sorghum production in the highland regions.
Kabanyolo (non-stress environment) was the best environment to identify early maturing sorghum lines, since both cold tolerant and susceptible genotypes were able to reach the final maturity stages. IESV 91003 LT, IESV 91054 LT, and IESV 91105 LT showed early maturity attributes, but showed partial tolerance to cold stress at Kachwekano. This indicates that those sorghum lines could possess recessive alleles for genes responsible for maturity, since they reduce days to flowering (Wang et al., 2015). However, this hypothesis needs to be tested through molecular and genetic analyses.
Generally, sorghum lines delayed by 37 and 63 days, at Zombo and Kachwekano, respectively, compared to non-stress weather conditions of Kabanyolo. Towards the end of the raining period, temperatures rose and cold stress was relieved, thus plants were able to reach their final plant height and complete maturity stage. Although all growth and phenological parameters decreased in all sorghum lines, cold-sensitive sorghum lines were seriously affected compared to tolerant variants. Moreover, physiological maturity was also affected since the grain filling period was longer in both cold environments, whereby the delay caused by this abiotic stress averaged 47 days at Zombo, and 73 days at Kachwekano.
Effects of cold stress on yield components
In tropical native plants like sorghum and maize, low temperature stress cause significant reduction in photosynthetic activity and biomass accumulation, which are the main sources of grain yield (Tari et al., 2013; Fiedler et al., 2014; Ortiz et al., 2017). In fact, cold stress negatively affects chlorophyll function, and consequently photosynthetic activities are significantly decreased (Allen and Ort, 2001; Tari et al., 2013). Furthermore, reproductive organs of plants grown cool environments can be seriously damaged and consequently cause reduction in the yield and yield components, when cold stress coincides with the grain filling period (Pereira da Cruz et al., 2006; Maulana and Tesso, 2013). In case where cold temperatures coincide with male and female organs formation, it may cause irreversible damages such as reduction anthesis rate, failed fertilization, reduced grain filling, which lead to the insufficient grain number per panicle and consequently low grain yield (Clarke and Siddique, 2004; Thakur et al., 2010; Maulana and Tesso, 2013).
The comparison between the non-stress (Kabanyolo) and cold stressed environments (Kachwekano and Zombo) indicated that yield related components were reduced for all evaluated genotypes, although some marked differences were identified whereby some genotypes could only yield a half of their actual performance as compared to Kabanyolo. However, cold tolerance is mainly determined by the levels of expression of cold tolerant responsive genes in the line per se (Janmohammadi et al., 2015). Cold tolerant genotypes have developed adaptation strategies to withstand cold stress through cold acclimation, whereby plants adjust to cold tolerance by exposing them to low but non-freezing temperatures (Thomashow, 1999, Chinnusamy et al., 2007). Genetic variability exists in sorghum adapted to high altitude areas of Africa, including the Eastern-African highland regions, which are considered as an important source of cold tolerant sorghum gene pool (Balota et al., 2010).
CONCLUSION
Low temperatures that coincide with vegetative period can affect various metabolic pathways, slowing growth rate and reduce photosynthetic activities. Consequently, susceptible plants would not survive, while tolerant genotypes with taller plant height can reach flowering and physiological maturity later. In the present study, sorghum genotypes with shorter stature coupled with tolerance to coldness were best ranked as early maturing in the cold environments of the highlands regions of Uganda, and thus can be used as parental lines for future breeding research based on line per se performance. Therefore, sorghum breeders need to constantly improve the genetic materials as far as flowering and grain filling period are concerned, as well as other agronomic traits based on farmer’s preferences, since this strategy would result in reducing yield penalty and contribute to enhancement of food security.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors are grateful to Makerere University for providing education and technical expertise, and ICRISAT (Kenya) for providing sorghum lines used in this research study. This research was funded by the Alliance for Green Revolution in Africa (AGRA).
REFERENCES
|
Allen DJ, Ort DR (2001). Impacts of chilling temperatures on warm-climate plants. Trends in Plant Science 6:36-42. |
|
|
Andres F, Coupland G (2012). The genetic basis of flowering responses to seasonal cues. Nature Reviews Genetics 13:627-639. |
|
|
Balota M, Payne WA, Veeragoni SK, Stewart BA, Rosenow DT (2010). Respiration and its relationship to germination, emergence, and early growth under cool temperatures in sorghum. Crop Science 50:1414-1422. |
|
|
Chinnusamy V, Zhu J, Zhu JK (2007). Cold stress regulation of gene expression in plants. Trends in Plant Science 12:444-451. |
|
|
Clarke HJ, Siddique KHM (2004). Response of chickpea to low temperatures stress during reproductive development. Field Crop Research 90:323-334. |
|
|
Fiedler K, Bekele WA, Duensing R, Gründig S, Snowdon R, Stützel H, Zacharias A, Uptmoor R (2014). Genetic dissection of temperature-dependent sorghum growth during juvenile development. Theoritical and Applied Genetics 127(9):1935-1948. |
|
|
Janmohhamadi M, Zolla L, Rinalducci S (2015). Low temperature tolerance in plants: Changes at the protein level. Phytochemistry 117:76-89. |
|
|
Kapanigowda MH, Perumal R, Aiken RM, Heral TJ, Bean SR, Little CR (2013). Analyses of sorghum [Sorghum bicolor (L.) Moench] lines and hybrids in response to early season planting and cool conditions. Canadian Journal of Plant Science 93:773-784. |
|
|
Kasozi SM, Bashaasha B, Ochwoh V (2005). Economics of sorghum production and soil fertility management in Kabale highlands, Uganda. Journal of Food, Agriculture and Environment 3:105-109. |
|
|
Liu L, Ji H, An J, Shi K, Ma J, Liu B, Tang L, Cao W, Zhu Y (2019). Responses of biomass accumulation in wheat to low-temperature at jointing and booting stages. Environmental and Experimental Botany 157:46-57. |
|
|
Mace ES, Hunt CH, Jordan DR (2013). Supermodels: sorghum and maize provide mutual insight into the genetics of flowering time. Theoretical and Applied Genetics 126:1377-1395. |
|
|
Maiti RK (1996). Germination and seedling establishment. In Maiti RK 1996. Sorghum science. Science publishers, Inc. Lebanon, NH. p. 41-98. |
|
|
Marais DL, Hernandez KM, Juenger TE and des Marais DL (2013). Genotype by-environment interaction and plasticity: exploring genomic responses of plants to the abiotic environment. Annual Review of Ecology, Evolution and Systematics 44:5-29. |
|
|
Maulana F, Tesso T (2013). Cold temperature episode at seedling and flowering stages reduces growth and yield components in sorghum. Crop Science 53:564-574. |
|
|
Murphy RL, Klein RR, Morishige DT, Brady JA, Rooney WL, Miller FR, Dugas DV, Klein PE, Mullet JE (2011). Coincident light and clock regulation of pseudoresponse regulator protein 37 (PRR37) controls photoperiodic flowering in sorghum: Proceedings of the National Academy of Sciences of the United States of America 108:16469-16474. |
|
|
Ortiz D, Hu J, Salas FMG (2017). Genetic architecture of photosynthesis in Sorghum bicolor under non-stress and cold stress conditions. Journal of Experimental Botany 68(16):4545-4557. |
|
|
Pereira da Cruz R, Milach SCK, Federizzi LC (2006). Rice cold tolerance at the reproductive stage in a controlled environment. Scientia Agricola 63(3):255-261. |
|
|
Quinby JR, Hesketh JD, Voigt RL (1973). Influence of temperature and photoperiod on floral initiation and leaf number in sorghum. Crop Science 13:243-246. |
|
|
Ramankutty N, Evan AT, Monfreda C, Foley JA (2008). Farming the planet: 1. Geographic distribution of global agricultural lands in the year 2000. Global Biogeochemical Cycles 22. |
|
|
Rapacz M, Tyrka M, Kaczmarek W, Gut M, Wolanin B, Mikulski W (2008). Photosynthetic acclimation to cold as a potential physiological marker of winter barley freezing tolerance assessed under variable winter environment. Journal of Agronomy and Crop Science 194(1):61-71. |
|
|
Reddy BVS, Ramesh S, Kumar AA, Wani SP, Ortiz R, Ceballos H, Sreedevi TK (2008). Bio-fuel crops research for energy security and rural development in developing countries. Bioenergy Research 1(3-4):248-258. |
|
|
Rooney WL, Aydin S (1999). Genetic control of a photoperiod-sensitive response in Sorghum bicolor (L.) Moench. Crop Science 39:397-400. |
|
|
Rymen B, Fiorani F, Kartal F, Vandepoele K, Inzé D, Beemster GTS (2007). Cold nights impair leaf growth and cell cycle progression in maize through transcriptional changes of cell cycle genes. Plant Physiology 143:1429-1438. |
|
|
Singh SP (1985). Sources of cold tolerance in grain sorghum. Canadian Journal of Plant Science 65:251-257. |
|
|
Solanke AU, Sharma AK (2008). Signal transduction during cold stress in plants. Physiology and Molecular biology of plants 14(1&2):69-79. |
|
|
Tari I, Laskay G, Takacs Z, Poor P (2013). Response of sorghum to abiotic stresses: a review. Journal of Agronomy and Crop Science 199:264-274. |
|
|
Thakur P, Kumar S, Malik JA, Berger JD, Nayyar H (2010). Cold stress effects on reproductive development in grain crops: An Overview. Environmental and Experimental Botany 67:429-443. |
|
|
Thomashow MF (1999). Plant cold acclimation: freezing tolerance genes and regulatory mechanisms. Annual Review Plant Physiology and Plant Molecular Biology 50:571-599. |
|
|
Wang Y, Tan L, Fu Y, Zhu Z, Liu F, Sun C (2015). Molecular Evolution of the Sorghum Maturity Gene Ma3. PLoS ONE 10(5):1-13. |
|
|
Yadav SK (2009). Cold stress tolerance mechanisms in plants: A review. Agronomy for Sustainable Development 30:515-527. |
|
|
Zhu J, Dong CH, Zhu JK (2007). Interplay between cold-responsive gene regulation, metabolism and RNA processing during plant cold acclimation. Current Opinion on Plant Biology 10:290-295. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


