Full Length Research Paper

ABSTRACT
Brown blotch disease caused by Colletotrichum capsici, is one of the most important disease affecting cowpea production in Burkna Faso. Every year the disease can cause yield losses as high as 42 to 100% on farmer’s fileds. This study aimed to understand the inheritance of its resistance in cowpea and also identify quantitative trait loci (QTL) associated with the resistance. Seedlings of parents and F1, F2 and BC1F1 progenies from crosses involving four susceptible and four resistant parents were screened in greenhouse conditions for brown blotch disease reaction. The inheritance studies indicated that different genes action were involved in the resistance of brown blotch resistance disease. In KN-1 variety, two or several genes associated to brown blotch were suggested. In Donsin local, a partially dominant gene, Rcc1, would be associated with the resistance. In Moussa local, a single major dominant gene, Rcc2, was identified. A single major recessive gene, rcc2, was found to control resistance with brown blotch in the cowpea resistant variety IT93K-503/46-13. The research of gene or QTL associated with brown blotch in cowpea variety KN-1 has been investigated. Polymorphic single nucleotide polymorphisms (SNP) markers and QTL associated to brown blotch resistance were discovered from 63 F2 individuals from a cross involving the susceptible variety Tiligre and the resistant KN-1. More than 11800 polymorphic SNP markers covered 1019.3 cM of the whole genome of cowpea were identified. A single dominant QTL, nomed qBBDR2 and 102 individual SNP markers tightly associated with the resistance of C. capsici were identified on chromosome 2 (Phytozome) of the cowpea genome.
Key word: Cowpea, Colletotrichum capsici, resistance, inheritance, single nucleotide polymorphisms (SNP) marker, QTL.
INTRODUCTION
Brown blotch disease caused by Colletotrichum capsici continues to cause enormous damage by seriously compromising development policies in terms of food security. However, the use of resistant varieties to brown
blotch disease, combined with appropriate agricultural practices, is a very effective and sustainable strategy to control the disease (Adebitan et al., 1992; Enyiukwu et al., 2014; Obi and Barriuso-vargas, 2014). Several varieties of cowpea, local and improved, have been identified as resistant and constitute real resources of exploitable genes. Some of these varieties have been investigated for resistance genes, including Striga gesnerioides (Ouédraogo et al., 2001; Boukar et al., 2004; Tignegre, 2010), Aphids (Boateng, 2015), Macrophomina phaseolina (Muchero et al., 2011; Pottorff, 2014), Fusarium oxysporum (Pottorff, 2014), and CABMV (Barro, 2016). However, information on C. capsici resistance genes is limited. This is because of the economic importance of brown blotch, the research of new sources of resistance is indispensable for identifying genes associated with the resistance. The breeding program will be considered a success when it takes into account the variability of the pathogen’s virulence. Indeed, the research conducted by Sérémé (1999) has highlighted the existence of different pathotypes of C. capsici. This high pathogenic variability of the C. capsici strains, makes selection much complex, a real challenge for cowpea breeders. The inheritance study of cowpea resistance to brown blotch has been undertaken. Abadassi et al. (1987) have identified a single recessive gene designed rcc controlling resistance to brown blotch in the resistant variety IT82E-16. Recently, Adetumbi et al. (2016) revealed a polygenic action of genes with quantitative effect to brown blotch in the resistant variety IT-95K-193-12. Similar studies conducted by Redden (1983) in cowpea seeds resistance to bruches (Callosobruchus maculatus) revealed a contribution of moderately resistant lines to susceptibility rather than resistance. Sérémé (1999) suggested a probable existence of quantitative effect genes involved in resistance to brown blotch in several genotypes. Recently, Thio et al. (2017) identified new sources of resistance in different cowpea genotypes. Among them, the cowpea genotype KN-1 showed stable resistance to the three isolates of C. capsici. Three of the genotypes, Moussa local, Donsin local and IT93K-503/46-13 showed interesting levels of resistance to C. capsici isolates and could be used in crosses with cowpea susceptible elite varieties.
In the present study, we described the nature of inheritance of resistance and detected QTL associated with brown blotch disease in cowpea.
MATERIALS AND METHODS
Plant
Crosses were made in greenhouse between four brown blotch susceptible cowpea varieties parents and four resistant cowpea varieties. The susceptible cowpea varieties group consists of the parents Tiligre (KVx775-33-2G), KVx61-1, KVx396-4-5-2D and Bambey-21. These susceptible genotypes have interesting agronomic performances and were used in the breeding program to increase their resistance to different biotic stresses. The resistant cowpea varieties include KN-1, resistant to all three isolates of C. capsici. Moussa local and Donsin local are both resistant to isolates C.cap-FA and C.cap-SA. The IT93K-503/46-13 variety, developed
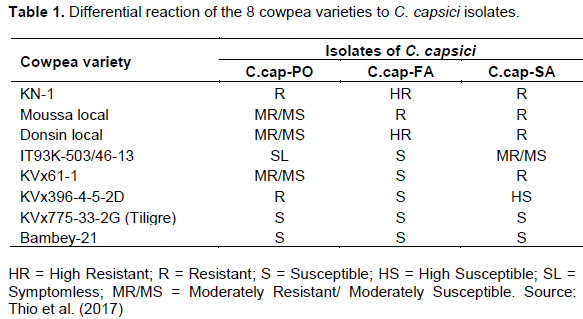
by IITA, is resistant only to C.cap-PO isolate with a higher level of resistance compared to the other three resistant varieties. Table 1 shows the differential reaction to the brown blotch disease for all the varieties (Thio et al., 2017).
Fungal material
The fungal material consists of the three isolates C.cap-SA, C.cap-FA and C.cap-PO of C. capsici identified as the most virulent in previous studies (Thio et al., 2017).
Population development
The four susceptible parents were used as female in crosses with the resistant parents to generate F1, F2 and BC1F1 population. Four to five seeds from each parent were planted in greenhouse in 10-L pots containing an unsterilized mixture of soil and sand to produce F1 plants. The F1 plants were advanced on one hand for selfing to produce F2 seeds (Figure 1), on the other hand, used as females with each of the two parents to produce BC1 backcross populations, BC1S and BC1R. Table 2 shows the different crosses made in this study.
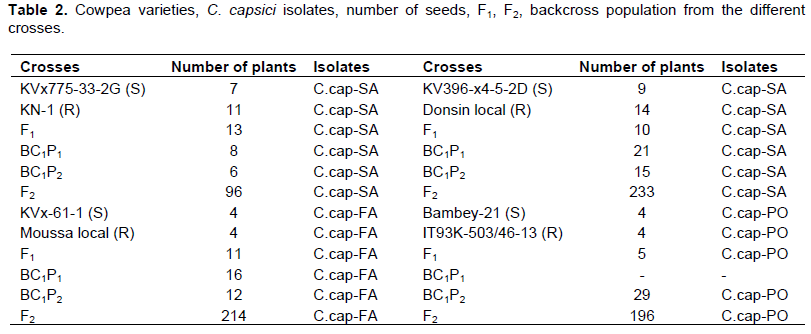
The different crosses along with their parents were evaluated in greenhouse condition from Mars 2016 to the end of May 2016. A mixture of soil, sand and organic fertilizer was autoclaved at 121°C for 30 mn and used to fill up 75% of the 2.5 L size pots. Seeds of the different populations and their parents were surface sterilized in 1% sodium hypochlorite for 5 min. Two seeds were sown in each pot.
Inoculation
The test materials (parents, F1, F2, BC1P1 and BC1P2) were evaluated in greenhouse condition for their reaction to brown blotch disease. The spore suspension from fresh fungi culture of 7 days was prepared and adjusted to a concentration of 106 spores/ml. The C. capsici suspension was applied by spraying over the entire surface of each population and their parents. Inoculated plants were kept under polyethylene plastic to maintain moisture overnight in order to allow better contamination (Figure 2).

Disease evaluation
Symptoms of brown blotch were scored at the 7th, 14th, 21st, 35th and 56th days after inoculation (DAI). The rating scale used ranges from 1 to 5 with score 1 = no symptoms, 2 = presence of small brown spots on the stem, 3 = coalescent spots on the stem, 4 = coalescent spots with presence of acervules on the stem, but not dead plants, and 5 = wilted stems, dead plants.
The disease severity index (severity) was estimated for all individuals screened genotypes. The disease severity index calculated from Allen et al. (1981) was applied to the F1, BC1 and F2 generations. For the evaluation of the different segregation reports, a method adopted by Voorrip et al. (2004) was used. This method uses both disease incidence and severity index as a criterion for classifying individuals into resistant and susceptible phenotypes. The severity index for segregated individuals was estimated by considering the individual score assigned to each genotype. Thus, individuals with scores 1 and 2 were considered resistant and those with scores 3 to 5 were considered susceptible.
Statistical analysis of the data
The analysis of the segregation data was done using Chi-square test. This test gives the calculated Chi-square values corresponding to the different phenotypic segregation ratios: 1R: 1S, 3R: 1S, 15R: 1S and 1R: 3S. These different ratios are related to the type of genes and the number of genes involved in the expression of resistance or susceptibility. The value of the Chi-square (c2) was calculated according to the formula:
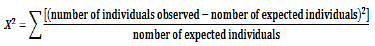
The null hypothesis (the segregation ratio follows Mendel’s law with different proportions) is accepted (value of Chi-square is not significant) at the threshold of 5% if c2 < 3.84 (ddl =1); P <0.05.
Illumina genotyping of F2 population and their parents
Leaves from parental lines Tiligre and KN-1 (3 samples each) and 90 F2 plants from cross between brown blotch-susceptible Tiligre and the multi-race-resistant variety KN-1 previously used were collected and dried at 35°C for 24 to 48 h. All the dried samples were transferred to Timko Laboratory for genotyping.
Amounts of 25 mg dry matter from each leave sample were used for genomic DNA extraction using a modified CTAB protocol (Ripoll et al., 2011). DNA from each individual was quantified using Nanodrop 2000/2000C and adjusted to the concentration of 50 ng/µl of DNA. The DNA samples were sent to the Center for Public Health Genomic at UVA for SNP genotyping using Illumina technology. Genotyping data were analyzed using GenomeStudio software version 2.0 (Illumina 2016).
Linkage map construction and QTL detection
A population of 63, semi-randomly selected from F2 individuals were genotyped with 11815 iSelect Consortium SNPs markers recovered all 11 cowpea chromosomes (Vigna unguiculata v1.0, NSF, UCR, USAID, DOE-JGI, http://phytozome.jgi.doe.gov/). Some of the 90 F2 individuals were excluded due to suspected derivation from incompletely inbred parental lines. Data from all SNP markers were used to construct linkage map using QTLIciMapping version 4.1 (Wang et al., 2011; Meng et al., 2015). The disease incidence was used as phenotypic value of the F2 population for marker-phenotype association analysis. Previously, the F2 population segregated into a 15 resistant: 1 susceptible ratio for both disease severity and incidence, suggested a polygenes action in KN-1 resistance to C. capsici. QTL mapping was performed using the ICIM add (Inclusive Composite Interval Mapping Additive) (Wang et al., 2011; Meng et al., 2015). Linkage association was done using logarithm of the odds (LOD). The value of the LOD score for which a QTL is significant was set to 3 (LOD score threshold). A largely positive LOD score (LOD score max > LOD score threshold P < 0.05) is synonymous with co-segregation between the marker and the phenotype and, conversely, a low LOD score means no linkage. The PVE values for which a QTL is declared major (PVE > 10) were determined according to the threshold of Collard et al. (2005).
RESULTS
Distribution of brown blotch in populations from crosses involving Tiligre and KN-1
The appearance of symptoms in the Tiligre variety used as a susceptible parent was observed from the 7th day after inoculation. From this date, the severity ratings of the disease remained almost constant until the 35th DAI where variations were observed. The assessments of the incidence and the severity of the disease at the 35 and 56th DAI within the different populations are presented in Table 3. The incidence of the disease in the susceptible parent Tiligre was significantly high, at 71.43%, at both 35 and 56th DAI. All the plants of resistant parent KN-1 were symptomless (0%) on both scoring dates.
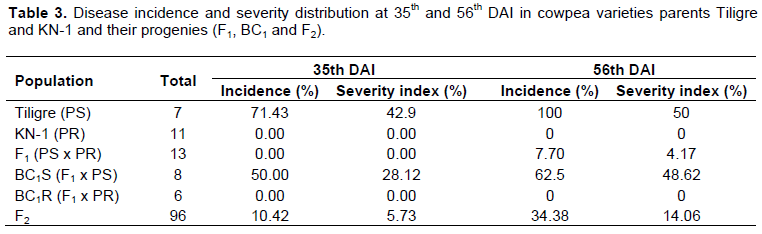
The severity of the disease in the susceptible parent Tiligre indicated 42.9 and 50%, respectively at the 35 and 56th DAI. All of the plants of resistant parent KN-1 were symptomless. The F1 hybrids of all the crosses involving Tiligre and KN-1 were uniformly resistant at 35th DAI. At the 56th DAI, the presence of symptoms in a single F1 hybrid was observed with an average of 4.17%. The distribution of the disease in terms of severity in the F2 population gives 5.73 and 14.06% respectively at 35 and 56th DAI. They are significantly lower than those obtained in the susceptible parent Tiligre.
Segregation pattern for resistance in brown blotch in KN-1
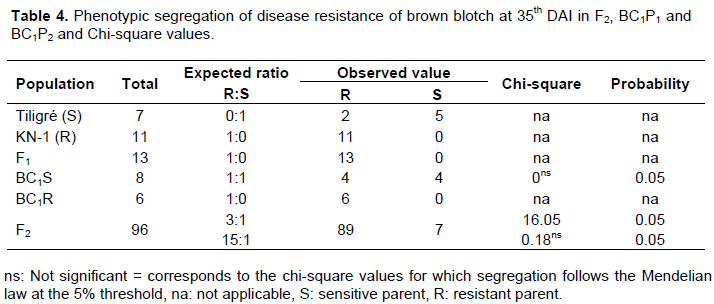
The data on genetic segregation in different populations derived from the crosses involving Tiligre and KN-1 at 35th DAI are shown in Table 4. The segregation pattern in the F2 population showed a good fit to 89R: 7S. The segregation pattern in the BC1S and BC1R population gives, respectively 4R: 4S and 6R: 0S at 35th DAI. These data indicated that resistance to C. capsici isolate C.cap-SA in the line KN-1 is controlled by two or several genes. It is a case of polygeny action.
Inheritance of resistance in Moussa local
As previously seen, brown blotch symptoms were observed on the susceptible variety KVx61-1 from the 7th day after inoculation.
The disease infection estimated by incidence and severity index varied throughout the experiment. All the plants of the susceptible parent KVx61-1 were severely affected by brown blotch, with 100 and 31.25%, respectively for incidence and severity for both 35 and 56th DAI (Table 5). Thus, all the plants of the resistant parent Moussa local were asymptomless (0% of incidence and severity) during the experimentation. Almost all the plants of F1 hybrid were completely free of brown blotch symptoms. Only one F1 hybrid showed symptoms of brown blotch with a score of 2 at the 56th DAI.The distribution of disease incidence in the F2 population was 30.84 and 43.92%, respectively at 35 and 56th DAI. The disease severity mean in the F2 population was 11.80% at 35th DAI., indicating the resistant status of this population. At the 56th DAI, the severity means in the F2 population was around 19.16%, indicating the F2 population was moderately resistant.
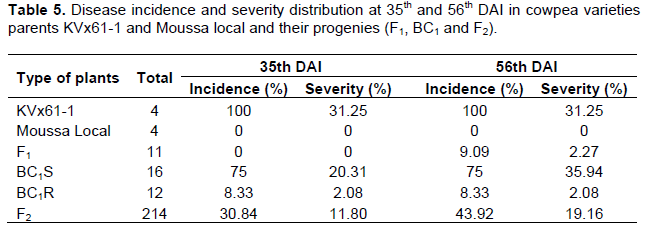
Figure 3 shows symptoms of brown blotch in the F2 individual plant at 35 DAI.

Segregation pattern for resistance in brown blotch in Moussa local
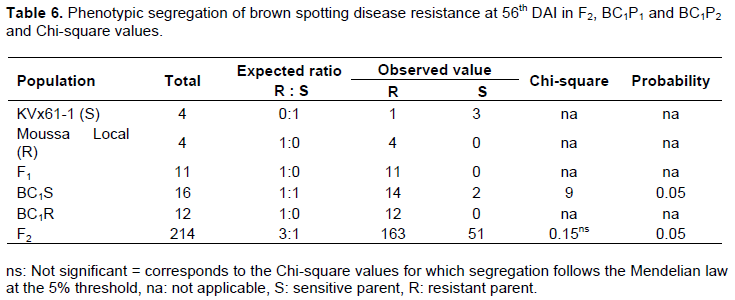
The segregation pattern in the F2 population showed a good fit to 183R: 31S and 163R: 51S, respectively at the 35 and 56th. The backcross to the susceptible parent BC1P1 showed a good fit to 15R: 1S and 14R: 2S, respectively at 35 and 56th DAI. The backross involving resistant parent BC1P2 was uniformly resistant as indicated of ratio 12R: 0S, at both the 35 and 56th DAI. The segregation pattern in the backcross population BC1P1 is not consistent with those expected. However, the segregation in the F1 hybrids and F2 population for resistance to brown blotch suggested that a major dominant gene is responsible for the resistance in Moussa local at the 56th DAI (Table 6).
Inheritance of resistance in Donsin local
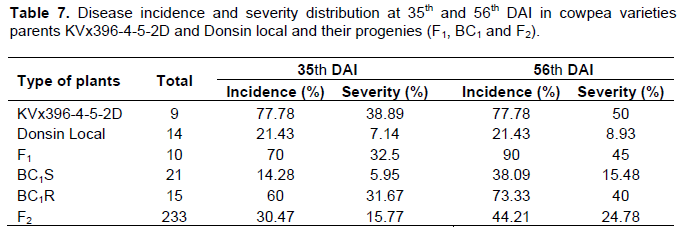
The responses of F1, F2, BC1 population and their two parents, inoculated with isolate C.cap-SA of C. Capsici is shown in Table 6. As previously seen in the first two tests, a progressive appearance of brown blotch symptoms was observed from the 7th DAI, especially in the susceptible parent KVx396-4-5-2D and the F2 population. The two parents, KVx396-4-5-2D and Donsin local showed symptoms of brown blotch but the disease severity in the susceptibe parent was very high with 38.89 and 50% respectively at 35 and 56th DAI.
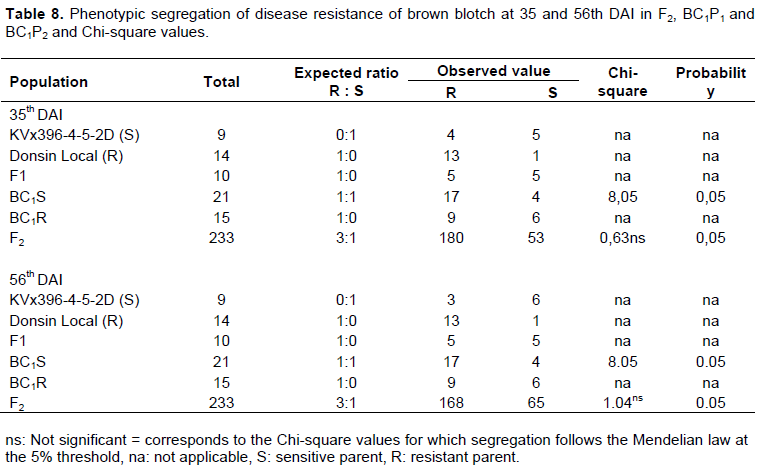
However, the disease incidence and severity in the resistant parent Donsin local was low compared to the susceptible parent (Table 7). The distribution of the disease incidence and severity in the F1 hybrids indicated a variation with a high proportion of susceptible individuals for both dates. In the F2 population, the disease incidence and severity index ranged from 30.47 to 44.21%, from 15.77 to 4.78%, respectively at 35 and 56th DAI. The segregation pattern in the backcross and F2 populations derived from the crosses are shown in Table 8. The results from the segregation pattern in the F2 population give a good fit to 180R: 53S and 168R: 65S, respectively at 35 and 56th DAI. These results suggest a single dominat gene governing resistance in Donson local variety. However, the F1 hybrids and the backcross resistant parent BC1P2 (BC1R) were not all resistant as expected (Table 8). This does not conform to the case of a total dominance.
Inheritance of resistance in IT93K-503/46-13
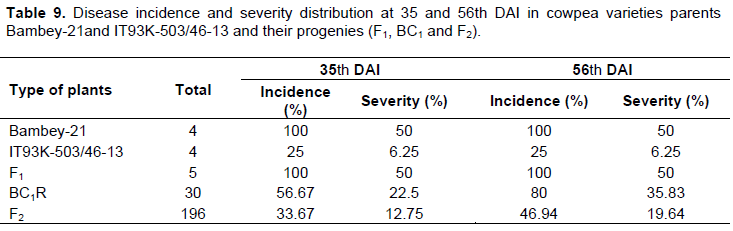
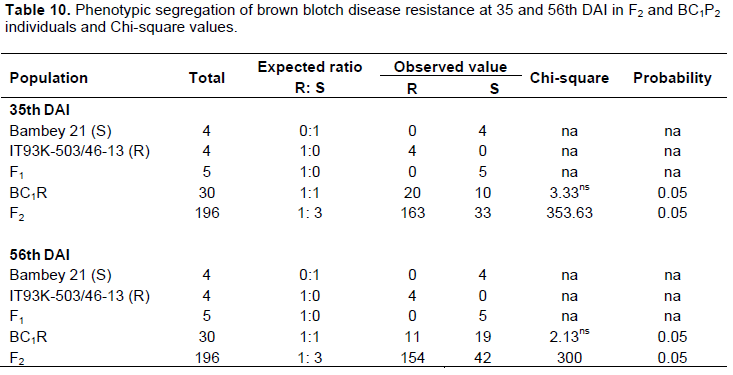
All the plants of the susceptible parent Bambey-21 showed symptoms of brown blotch disease (Figure 4). Almost all the plants of the resistant parent were resistant to brown blotch but one plant showed the symptoms of the disease with a score of 2 (as resistant). All F1 hybrids tested showed symptoms of brown blotch with a score of 3. The distribution of the disease incidence and severity in the F1 hybrids is in the order of 100 and 50% respectively, at the 35 and 56th DAI (Table 9). Symptoms of brown blotch were widely observed in backcross parent resistant and the F2 population. The results of the severity index in backross and F2 populations give 56.67 and 80% and 12.75 and 19.64%, respectively at the 35 and 56th DAI.The segregation patterns in the backross and F2 population are shown in Table 10. The backcross to the resistant parent BC1P2 (BC1R) fitted very closely to a 1R:1S ratio at both dates. The segregation pattern in the F2 population gives 163R: 33S and 154R: 42S, respectively at the 35 and 56th DAI. This report does not correspond to the expected ratio (1R: 3S). However, the segregation patterns in the backcross BC1R and the susceptibility of all F1 hybrids indicated that resistance to brown blotch in the IT93K-503/46-13 variety is monogenic recessive.

Genetic mapping based on SNP markers
A total of 11,813 polymorphic and codominant SNPs markers covering 1019.3 cM and averaged 92.66 cM between markers, indicating good overall coverage of the genome. Deviations related to Mendelian segregation were tested by the Chi-square test. Out of the 11813 polymorphic SNPs markers, only 7416 normally segregate (c2 < 6.64; P < 0.01). The distribution of all these markers on the 11 chromosome pairs of cowpea is shown in Table 11.
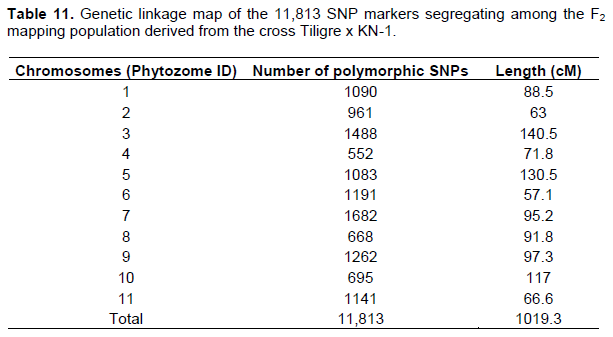
QTL detection associated to brown blotch in KN-1 variet
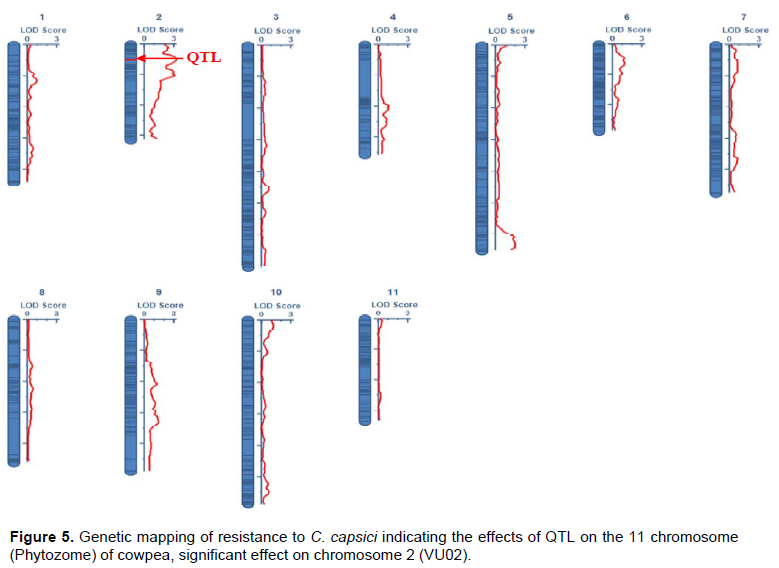
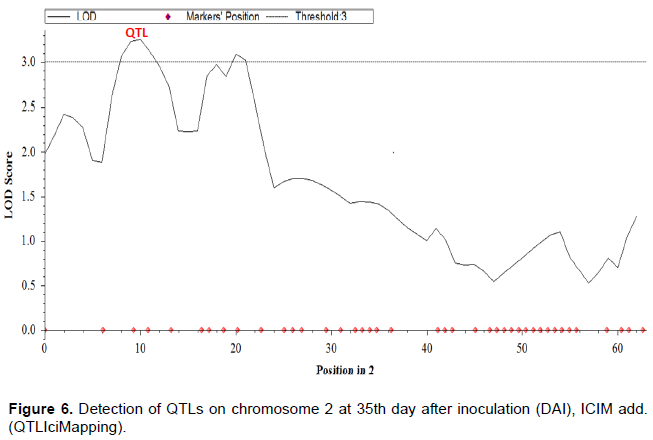
QTL analysis using the association test between the 11,813 polymorphic SNPs markers and the phenotype of disease incidence of the 63 F2 population showed the effects of probable QTLs on all 11 chromosomes of cowpea (Figure 4). A positive and significant effect of QTL (P < 0.05) was observed on chromosome 2 (Figure 5). The detected QTL explained 21.44% of the phenotypic variance (PVE), had an estimated of position 10 cM on the cowpea genome (Figure 6). The QTL detected showed an additive effect of 0.23 and associated to the alleles of the resistant parent KN-1 (Table 12). The LOD max score value for which the resistance QTL was declared significant in the order of 3.25 (> LOD threshold score, 3). The genomicregion flanking the resistanceQTL includes markers 2_21435 and 2_02435, respectively the left and right flanking markers. Additionaly, 102 individual SNPs markers (LOD score max > 3) associated with resistance to C. capsici were identified on chromosome Vu02.

DISCUSSION
Nature of genes involved in resistance in KN-1 and Donsin local
The distribution of resistant and susceptible phenotypes and Chi-square results from the crosses involving Tiligré and KN-1 revealed that resistance to C.cap-SA isolate of C. capsici in KN-1 was controlled by two or more genes with a quantitative effect (QTL). This resistance gene is believed to be from the resistant parent KN-1. Several genes with minor effects inducing different resistance mechanisms would be involved. These genes with minor effects occur at different stages of plant development and induce different mechanisms of resistance. Their effect contributes to the suppression of disease symptoms (Chun Ying et al., 2015).
Several studies on the inheritance of polygenic traits have been undertaken and have resulted in the identification of QTLs, some of which have been mapped. Chun Ying et al. (2015) identified several QTLs associated with resistance to Colletotrichum acutatum in the pepper. These QTLs were detected from F1 and BC1 individuals and would occur at different stages of plant development.
Contrary to the KN-1 variety, Donsin local variety tested with the same C.cap-SA isolate had a different reaction. Indeed, the behavior of F1 individuals at the 35 and 56th DAI did not conform to the expected results for a case of a totally dominant (1: 0) or totally recessive (0: 1) gene. Moreover, phenotypic segregation within backcross individuals (BC1P1 and BC1P2) also does not follow Mendelian law. This suggests a partial or incomplete dominance in Donsin local resistance to C.cap-SA isolate. It is most likely that the Donsin local variety is heterozygous. Heterozygous F2 individuals are thought to belong to the intermediate class (moderately resistant/moderately susceptible). This would explain the phenotypes of F1 and backcross individuals observed.
According to the concept of gene-for-gene developed by Flor (1942), the differential reaction observed in the local KN-1 and Donsin varieties, inoculated by the same isolate, suggests that these two varieties would have a different resistance gene. Considering the nomenclature proposed by Abadassi et al. (1987), the resistance gene identified in Donsin local variety was designated Rcc1 (resistance to C. capsici, dominant gene 1).
Nature of genes involved in resistance in Moussa local
The study of the inheritance of resistance of Moussa local variety to the isolate C.cap-FA, from the crosses involving KVx61-1 and local Moussa, suggests that a major dominant gene is responsible for this resistance. Previous studies by Sérémé (1999) had demonstrated resistance of the Moussa local variety to several C. capsici isolates and suggested a case of polygeny. It is not excluded that this variety has lost over time components involved in its resistance mechanisms or new virulence have raised.
The emergence of new C. capsici pathogens to which Moussa local variety was monogenic resistant is much more plausive. Several variants of the pathogens with different aggressiveness levels were identified (Thio et al., 2017). Mahasuk et al. (2009) identified a major dominant gene in chilli pepper resistance to anthracnose caused by C. acatatum. That gene was identified during the ripe fruit stage. Two new genes conferring resistance to the most aggressive pathotypes of C. acatatum in chili pepper were identified (Mahasuk et al., 2013).
The resistance gene identified in Moussa local variety was designated Rcc2 (Resistance to C. capsici dominant gene 2) as previously stated.
Nature of gene governing resistance in IT93K-503/46-13
It was appropriate to address the inheritance of resistance in IT93K-503/46-13, especially since this variety had demonstrated total resistance to C.cap-PO isolate. After analysis of the different phenotypic segregation ratios and disease distribution in the F2 population and BC1P2 (resistant parent backross) progeny, a major recessive gene responsible for resistance was identified.
Previous studies conducted by Abadassi et al. (1987) identified a recessive gene known as rcc (resistance to C. capsici) associated with brown blotch disease in IT82E-16 variety.
Kim et al. (2008) also identified a recessive gene governing chili pepper resistance to anthracnose disease caused by C. capsici in South Korea. A previous study conducted by Mahasuk et al. (2009) identified a recessive gene in chili pepper resistance to anthracnose. That gene was identified during the mature green fruit stage and is believed to be responsible for the spread of the disease. The resistance gene identified in IT93K-503/46-13 variety was designated rcc2 (resistance to recessive C. capsici gene 2).
QTL detection and markers associated to partial resistance of KN-1 variety to C. capsici
The polygenic inheritance of KN-1 variety has been prospected in marker-phenotype associations tests using QTL analysis. A dominant QTL located on chromosome 2 (Vu02) involved in partial resistance of cowpea to C. capsici was detected. The resistance QTL showed a PVE value of 21.4% and a positive additive effect of +0.2349, indicating that resistance is controlling by the favour alleles of the parent KN-1. Disease manifestation is due to the alleles of the susceptible parent Tiligré. The dominance effect of the resistance QTL was in the order of -0.28 indicating the superiority of the dominant parent KN-1 in terms of disease incidence and severity. However, the effects of the detected QTL were relatively low in terms of PVE value, additivity and dominance effect (Bratteler et al., 2006).
Kulkarni (2009) attributes this situation to unstable and unfavourable environmental conditions that do not promote disease symptoms developement and therefore better detection of QTL. Ferreira et al. (2006) demonstrated the need for a sufficient population, at least 200 F2 individuals, to detect QTL with a significant effect. Good saturation of the genetic map in polymorphic and codominant markers would promote better detection of QTL (Liu, 1998).
Working on brown blotch in Nigeria, Adetumbi et al. (2016) identified more than 130 SNPs markers in IT-95K-193-12 cowpea variety. Unlike the results of this study, they did not detect any genes or QTL. Several QTL associated with Colletotrichum lindemuthianum have been reported on chromosome 2 of Phaseolus vulgaris, a close relative of cowpea, suggesting that this linking group may play an important role in resistance to Colletotrichum species (Campa et al., 2014; Oblessuc et al., 2014; Silva, 2017). Darvishzadeh (2007) identified several QTLs and AFLPs markers associated with sunflower partial resistance to Phoma macdonaldii. These resistance QTLs were located primarily on linkage groups 5, 8 and 15.
The resistant QTL detected in this study could be named qBBDR2 (QTL for brown blotch disease resistance located on chromosome 2).
CONCLUSION
The inheritance study of brown blotch in cowpea indicated that genes of different nature are controlling the disease resistance. In KN-1, resistance is controlled by two or more genes with a quantitative effect. QTL analysis with SNP markers allowed to detection of a major dominant resistant QTL on chromosone 2 (Vu02) in KN-1 variety. In the Moussa local variety, resistance is monogenic and controlled by a dominant gene. This type of resistance is easy to handle and is frequently used in Marker-Assisted-Selection. In the Donsin local variety, resistance seem to be controlled by a partial-dominant gene.
A recessive gene is governing resistance in cowpea resistant variety IT93K-503/46-13.
Several thousand polymorphic SNPs markers were identified in KN-1 resistance and some of them were associated with brown blotch. These SNP markers could be converted into PCR based marker and used for population validation in Marker-Assisted-Selection.
The efficient exploration of converted SNP markers will identify among them those associated to C. capsici resistance genes in cowpea resistant varieties Moussa local and Donsin local.
CONFLICT OF INTERESTS
The author has not declared any conflict of interests.
REFERENCES
|
Abadassi JA, Singh BB, Ladeinde TOA, Shoyinka SA, Emechebe AM (1987). Inheritance of resistance to brown blotch, septoria leaf spot and scab in cowpea (Vigna unguiculata (L.) Walp.). Indian Journal of Genetics and Plant Breeding 47:299-303. |
|
|
Adebitan SA, Ikotun T, Dashiell KE, Singh, SR (1992). Use of three inoculation methods in screening cowpea genotypes for resistance to two Colletotrichum species. Plant Disease 76(10):1025-1028. |
|
|
Adetumbi JA, Akinyosoye ST, Olowolafe MO, Oloyede-Kamiyo QO, Agbeleye OA (2016). Genetic linkage map of cowpea (Vigna unguiculata (L.) Walp) using SNP markers. African Journal of Biotechnology 15(20):830-834. |
|
|
Allen DJ, Emechebe AM, Ndimande B (1981). Identification of resistance in cowpea to diseases of the African savannas. Tropical Agriculture 58(3):267-274. |
|
|
Barro A (2016). Etude génétique de la résistance du niébé [Vigna unguiculata (L.) Walp] au cowpea aphid- borne mosaic virus (CABMV) au Burkina Faso. Université Ouaga I Pr Joseph KI-ZERBO.75 p. |
|
|
Boateng AB (2015). Genetic studies of Aphids (Aphis craccivora Koch) resistance in cowpea. PhD Thesis, Kwame Nkrumah University of Science and Technology. 67p. |
|
|
Boukar O, Kong L, Singh BB, Murdock L, Ohm HW (2004). AFLP and AFLP-derived SCAR markers associated with Striga gesnerioides resistance in cowpea. Crop Science Society of America 44(4):1259-1264. |
|
|
Bratteler M, Baltisberger M, Widmer A (2006). QTL analysis of intraspecific differences between two Silene vulgaris ecotypes. Annals of Botany 98(2):411-19. |
|
|
Campa A, Rodríguez-Suárez C, Giraldez R, Ferreira JJ (2014). Genetic analysis of the response to eleven Colletotrichum lindemuthianum races in a RIL population of common bean (Phaseolus vulgaris L.). BMC Plant Biology 14(1):114-115. |
|
|
Chun Ying S, Sheng Li M, Zheng Hai Z, Alain P, Li Hao W, Bao Xi Z (2015). Resistances to anthracnose (Colletotrichum acutatum) of Capsicum mature green and ripe fruit are controlled by a major dominant cluster of QTLs on chromosome P5. Scientia Horticulturae 181:81-88. |
|
|
Collard BCY, Jahufer MZZ, Brouwer JB, Pang ECK (2005). An introduction to markers, quantitative trait loci (QTL) mapping and marker-assisted selection for crop improvement: The Basic Concepts. Euphytica 142(1-2):169-196. |
|
|
Darvishzadeh R (2007). Déterminisme génétique de la résistance du tournesol au Phoma. Thèse de Doctorat de l'Institut National Polytechnique de Toulouse. Institut National Polytechnique de Toulouse. 201p. |
|
|
Enyiukwu DN, Awurum AN, Ononuju CC, Nwaneri JA (2014). Biology and management strategies of cowpea anthracnose disease caused by Colletrotrichum species. Greener Journals of Biochemistry and Biotechnology 1(2):052-065. |
|
|
Ferreira A, da Silva MF, da Costa e Silva L, Cruz CD (2006). Estimating the effects of population size and type on the accuracy of genetic maps. Genetics and Molecular Biology 29(1):187-92. |
|
|
Flor HH (1942). Inheritance of pathogenicity in melampsora. Phytopathology 32:653-669. |
|
|
Illumina (2016). GenomeStudio ® Genotyping Module v2.0 Software Guide, 69p. |
|
|
Kim SH, Yoon JY, Do JW, Park HG (2008). A Major recessive gene associated with anthracnose resistance to Colletotrichum capsici in chili pepper (Capsicum annuum L.). Breeding Science 58(2):137-41. |
|
|
Kulkarni SA (2009). Epidemiology and integrated management. University of Agricultural Sciences, Dharwad, 168p. |
|
|
Liu BH (1998). Genomics, Linkage Mapping and QTL Analysis. CRC Press, Boca Raton, FL. |
|
|
Mahasuk P, Chinthaisong J, Mongkolporn O (2013). Differential resistances to anthracnose in Capsicum baccatum as responding to two Colletotrichum pathotypes and inoculation methods. Breeding Science 63(3):333-338. |
|
|
Mahasuk P, Taylor PWJ, Mongkolporn O (2009). Identification of two new genes conferring resistance to Colletotrichum acutatum in Capsicum baccatum. Phytopathology 99(9):1100-1104. |
|
|
Meng L, Li H, Zhang L, Wang J (2015). QTL IciMapping: Integrated software for genetic linkage map construction and Quantitative Trait Locus Mapping in Bi-parentale Populations. Crop Journal. Crop Science Society of China and Institute of Crop Science, CAAS 3(3):269-283. |
|
|
Muchero W, Ehlers JD, Close TJ, Roberts PA (2011). Genic SNP markers and legume synteny reveal candidate genes underlying QTL for Macrophomina phaseolina resistance and Maturity in Cowpea [Vigna unguiculata (L) Walp.]. BMC Genomics. BioMed Central Limited 12(8):14p. |
|
|
Obi VI, Barriuso-Vargas JJ (2014). Situation of biofungicides reconnaissance, a case of anthracnose disease of cowpea. American Journal of Plant Sciences 5:1202-1211. |
|
|
Oblessuc PR, Baroni RM, Pereira GDS, Chiorato AF, Carbonell SAM, Brinez B, Silva LDCE, Garcia AAF, Camargo LEA, Kelly JD, Benchimol-Reis LL (2014). Quantitative analysis of race-specific resistance to Colletotrichum lindemuthianum in common bean. Molecular Biology 34(3):1313-1329. |
|
|
Ouédraogo JT, Maheshwari V, Berner DK, St-Pierre C-A, Belzile F, Timko MP, (2001). Identification of AFLP markers linked to resistance of cowpea (Vigna unguiculata L.) to parasitism by Striga gesnerioides. Theoretical and Applied Genetics 102(6-7):1029-36. |
|
|
Pottorff MO (2014). Genetic mapping of Fusarium oxysporum F.sp. Tracheiphilum Race 3 and Race 4, Macrophomina phaseolina resistance and other traits in cowpea (Vigna unguiculata [L.] Walp). PhD Thesis, University of California Riverside, 304p. |
|
|
Redden R (1983). The inheritance of seed resistance to Callosobruchus maculatus F. in Cowpea (Vigna unguiculata L. Walp.). II* Analyses of percentage emergence and emergence periods of bruchids in F4 seed generations of two reciprocal crosses. Australian Journal of Agricultural Research 34(6):681-695. |
|
|
Ripoll J, Bon M-C, Walker J (2011). Optimalisation de l'extraction d'ADN génomique de la morelle jaune (Solanum elaeagnifolium Cav.), une plante invasive des milieux cultivés en région méditerranéenne. Biotechnology, Agronomy and Society and Environnement 15(1):95-100. |
|
|
Sérémé P (1999). La maladie des taches brunes du niébé (Vigna unguiculata) au Burkina Faso: connaissance des agents pathogènes impliqués et développement de méthodes de lutte. Thèse de Doctorat d'Etat ès-Sciences Naturelles, Université de Cocody, UFR Biosciences, Côte d'Ivoire. 222p. |
|
|
Silva LCD (2017). Linkage fine-mapping, Gwas and QTLs affecting morpho-agronomic traits of a common bean RIL. PhD Thesis., Universidade Federal de Viçosa. 118p. |
|
|
Thio IG, Zida PE, Néya BF, Batieno TBJ, Néya BJ, Sawadogo M, Sérémé P (2017). Differential Reaction of Cowpea Genotypes to Brown Blotch Disease (Colletotrichum Capsici) in Burkina Faso. African Journal of Agricultural Research 12(36):2773-2782. . |
|
|
Thio IG (2018). Etude génétique de la résistance du niébé (Vigna unguiculata (L.) Walp.) à la maladie des taches brunes causée par Colletotrichum capsici (Syd.) Butler et Bisby au Burkina Faso. |
|
|
Thèse de Doctorat Unique, Université Ouaga I Joseph KI-ZERBO, Burkina Faso. 211p. |
|
|
Tignegre J-BDLS (2010). Genetic study of cowpea (Vigna unguiculata (L.) Walp.) resistance to Striga gesnerioides (Willd.) Vatke in Burkina Faso. PhD Thesis, University KwaZumu-Natal, Republic of South Africa, pp. 1-170. |
|
|
Wang J, Li H, Zhang L, Li C, Meng L (2011). Users' manual of QTL IciMapping v3. 1. Institute of Crop Science Chinese Academy of Agricultural Sciences (CAAS) Beijing 100081, China and Crop Research Informatics Lab International Maize and Wheat Improvement Center (CIMMYT), 146p. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


