Full Length Research Paper

ABSTRACT
The development of resistance to antimalarial drugs is a major challenge for global malaria control. Full Plasmodium falciparum resistance to dihydroartemisinin–piperaquine treatment has been reported recently in Cambodia. These events were directly associated with increased copy number variations (CNVs) in the plasmepsin system, including the PfPlasmepsin 2 gene. Pfplasmepsin 2 copy number was the most significant molecular signature associated with dihydroartemisinin–piperaquine treatment failure. Even though the piperaquine resistance has not been observed in regions in which artemisinin resistance has not been documented, it is possible to find an amplification of the Pfplasmepsin 2 gene in these regions. In this present study, we investigate to do a surveillance of Pfplasmepsin 2 copy number variations in Senegal by qPCR. Pfplasmepsin 2 copy number was assessed in 120 P. falciparum positive patients, 60 from Dakar and 60 from Kedougou by qPCR and an amplification of the Pfplasmepsin 2 genes was measured by using five standards of mixed synthetic gene fragments. Using a copy number threshold of 1.7 and 1.73% carried a multiple copies of Pfplasmepsin 2, whilst one copy of the gene was found in 98.26% of the isolates. Our results show that the CNVs associated with resistance to piperaquine are probably already frequent in Senegal. Paradoxically, Pfplasmepsin 2 multi-copy is generally found in parts of Africa where dihydroartemisinin–piperaquine failures are rare and resistance to piperaquine has not yet been described. However, it is no evidence to confirm piperaquine resistance in Senegal.
Key words: PfPlasmepsin 2 copy number, Senegal, Piperaquine, resistance, drug resistance.
Abbreviation: CNV, Copy Number Variation; ACT, Artemisinin combination therapy; WHO, World Health Organization; NMCP, National Malaria Control Programme; AL, Arthmeter-Lumefantrine; DHA-PPQ, Dihydroartemisinin–Piperaquine; IPT, intermittent preventive treatment; SP, Sulfadoxine–Pyrimethamine; SMC, seasonal malaria chemoprevention; PfPM2, Plasmodium falciparum plasmepsin 2; DNA, deoxyribonucleic acid.INTRODUCTION
Artemisinin combination therapy (ACT), the use of a short acting artemisinin derivative and a long-acting partner drug, is recommended worldwide for the treatment of Plasmodium falciparum malaria (World Health Organization, 2018). In 2006, in line with World Health Organization (WHO) recommendation, the Senegalese National Malaria Control Programme (NMCP) introduced the ACT regimen with Arthmeter-Lumefantrine (AL) as first-line treatment of uncomplicated P. falciparum malaria and dihydroartemisinin–piperaquine (DHA-PPQ) combination was then recommended as a second-line treatment. Although DHA-PPQ, is not currently used as the first-line treatment against uncomplicated malaria, this antimalarial drug has been widely used to compensate for antimalarial drugs shortages in 2010 and 2011 (Thiam et al., 2012).
In addition, the NMCP recommended the intermittent preventive treatment IPT of pregnant women with sulfadoxine–pyrimethamine (SP) and seasonal malaria chemoprevention (SMC) for children with SP-amodiaquine (Bamba et al., 2013; Mbaye et al., 2017).
Unfortunately, the recent emergence of P. falciparum resistance to artemisinin derivatives in Southeast Asia challenges malaria control and elimination efforts. This situation is increasingly compromised by concurrent resistance to partner drugs in combination therapies, such as the piperaquine (Witkowski et al., 2017).
In 2014, the first report of DHA–PPQ treatment failures was published (Saunders et al., 2014). Therapeutic failures were estimated to reach 60% indicating a dramatic expansion of piperaquine resistance. Moreover, full P. falciparum resistance to DHA/PPQ treatment has been reported recently in Cambodia (Amato et al., 2017). These events were directly associated with increased copy number variations (CNVs) in the plasmepsin system, including the pfPM2 gene (PF3D7_1408000) coding for the food vacuole enzyme plasmepsin II (Sanogo et al., 2018). CNV is generally considered as emerging at relatively rapid mutation rates (Cheeseman et al., 2009).
The relevance of the Pfplasmepsin 2 (PfPM2) gene amplification is conferring any survival or fitness advantages in response to PPQ pressure. Piperaquine resistance is poorly characterized. It is currently identified by late clinical failures with amplification of the plasmepsin 2 copy number gene. Although several genetic variations have been associated with decreased piperaquine susceptibility: a single copy of the mdr1gene has been associated with dihydroartemisinin–piperaquine treatment failures in Cambodian patients (Witkowski et al., 2017).
However, PfPM2copy number was the most significant molecular signature associated with dihydroartemisinin–piperaquine treatment failure (Witkowski et al., 2017). The piperaquine resistance has not yet been observed in area where artemisinin resistance has not also been documented as Senegal. However, it is possible that plasmepsin 2 amplifications will be found in other regions where piperaquine has been used as a partner drug (Bopp et al., 2018). Thus the assessment of PfPM2 gene copy number to areas where piperaquine is being used in artemisinin-based combination therapies is important. In line with the NMCP strategy for the surveillance of antimalarial drug efficacy, the aim of this study was to do a surveillance of multiple copies of PfPM2 gene, potentially involved in piperaquine resistance.
MATERIALS AND METHODS
Study site and sample collection
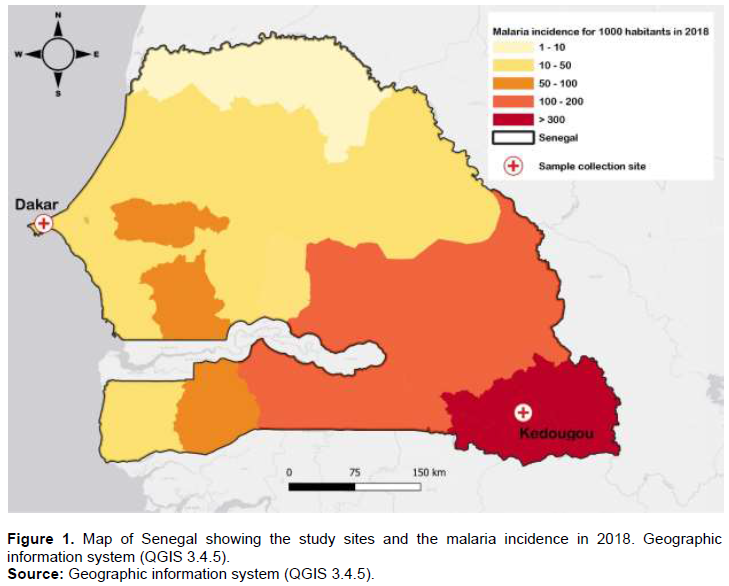
In total, 120 patient samples from two regions of Senegal were selected: Dakar (60), Kedougou (60), which present different levels of malaria transmission intensity (Figure 1). In Dakar malaria transmission is low and parasite prevalence is estimated at 1.3%. In the southeastern Kedougou region, the level of malaria transmission is high with an incidence higher than fifteen malaria cases per 1000 habitants (PNLP-Senegal, 2018). In Dakar samples were collected between October and November 2015 while in Kedougou the samples were collected in September 2016. These Samples were collected in the some months because these months corresponding to periods of high transmission in Senegal. For routine surveilliance in a region of low transmission such as Dakar, we estimate that a maximum sample size would be required to obtain a positive case compared to Kedougou with high transmission. Venous blood samples were collected in 5 ml vacutainer tubes and filter paper was made for molecular testing. All individuals in this study presented with uncomplicated malaria and parasite presence and species was confirmed by microscopy.
Ethical approval
The study protocol was approved by the National Ethics Committee for Health Research of Senegal (CNERS). Before participant recruitment and sample collections were initiated, written and informed consent was obtained from all participants.
Copy number variation assays
Parasite DNA was extracted from 120 samples using the QIAamp DNA Blood Mini kit (Qiagen) according to manufacturer instructions (QIAamp DNA Blood Mini Kit, Qiagen, Valencia,CA). The relative PfPM2 copy number were assessed by qPCR using real-time PCR machine (ABI 7500) as described by Witkowski with minor modifications in the Sequences of fragments synthetic genes (Witkowski et al., 2017). As a single copy endogenous gene control, we used the single copy β-tubulin gene and P. falciparum genomic DNA from strain 3D7 which has one copy of PfPM2 was included in each run as controls.
Briefly, quantitative PCR (qPCR) was carried out in 20 μl volumes in a 96-well plate containing 10 μl qPCR EvaGreen dye Supermix, 1 μl of each forward and reverse primer (Table 1), 3 μl H2O and 5 μl of template DNA. Amplifications were performed under the following conditions: 98°C for 3 min, followed by 40 cycles of 95°C for 10s and 58°C for 20 s and amplifications were run in triplicates.
Determination of the copy number
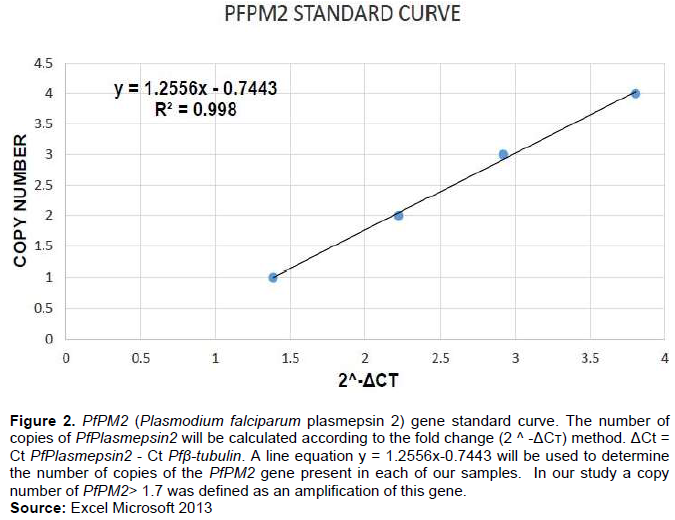
Amplification of the PfPM2 genes was measured by using five standards of mixed synthetic gene fragments (Table 2).The number of copies for each gene is determined relative to a standard curve. The standard curve is obtained from a mixture of fragments synthetic genes of Pfplasmepsin2 and Pfβtubulin by making five molar ratio following theses dilutions: standard 1 (1:1 PfPlasmepsin2-Pfβtubulin), standard 2 (2:1 PfPlasmepsin2-Pfβtubulin), standard 3 (3:1 PfPlasmepsin2-Pfβtubulin) and standard 4 (4:1 PfPlasmepsin2-Pfβtubulin)

The standards were also quantified according to the following Protocol: Quantitative PCR (qPCR) was carried out in 20 μl volumes in a 96-well plate containing 10 μl qPCR EvaGreen dye Supermix,1 μl of each forward and reverse primer (Plasmepsin2 primer for the Pfplasmepsin2 gene and β-tubulin primer for the Pfβ-tubulin gene), 6 μl H2O and 2 μl of template DNA. Amplifications were performed under the following conditions: 98°C for 3 min, followed by 40 cycles of 95°C for 10 s and 58°C for 20 s and amplifications were run in triplicates. The number of copies of PfPlasmepsin2 will be calculated according to the fold change (2 ^ -ΔC?) method.
ΔCt = Ct PfPlasmepsin2 - Ct Pfβ-tubulin and the Ct is deduced from the standard curve.
Once the 2 ^ -ΔC? is obtained for the five standards, we used it to make a standard curve with a line equation y = ax + b, it will be used to determine the number of copies of the PfPM2 gene present in each of our samples (Figure 2). In our study a copy number of PfPM2> 1.7 was defined as an amplification of this gene. Our CVN for the control (3D7) was 0.7.
RESULTS
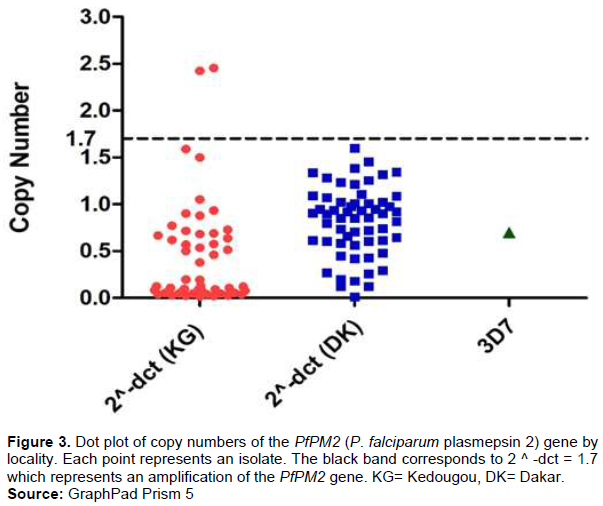
Using a copy number threshold of 1.7 the relative PfPM2 copy number was assessed in 120 P. falciparum positive patients, 60 in Dakar and 60 in Kedougou. From the samples with a valid copy number estimate (n = 115), 1.73% carried a multiple copies of PfPM2, whilst one copy of the gene were found in 98.26% of the isolates (Figure 3). A multiple copies were noted only in Kedougou with 3.33%. In Dakar all the samples presented a single copy.
DISCUSSION
DHA/PPQ has shown near-perfect efficacy levels in clinical trials conducted in Africa, the combination also has been proposed as second-line treatment of Plasmodium falciparum malaria in Senegal. Unfortunately, full P. falciparum resistance to DHA/PPQ treatment has been reported recently in Cambodia. These events were directly associated with increased copy number variations in the PfPM2 gene. Therefore, preexisting PfPM2 duplications in Cambodia might have been rapidly selected by DHA/PPQ, aided by a less effective protective action of the artemisinin derivative (Hastings et al., 2016). Such a scenario suggests that this copy number variation may already be present in Africa. Accurate and timely surveillance of drug resistance markers aids in maintaining and prolonging the efficacy of the limited selection of anti-malarial drugs available. Thus, it becomes opportune to follow this emergence and it is for this reason that we have chosen to study the number of copies of the PfPM2 gene in order to determine the number of copies circulating in each individual of P. falciparum in Dakar and in Kedougou. 1.73% carried a multiple copies of PfPM2, whilst one copy of the gene were found in 98.26% of the isolates from Kedougou (Figure 2).The copy number of PfPM2 genes was differ between isolates from different sites. An amplification of this gene was only reported in Kedougou. Kedougou is a particular site with presence of gold mining, more than 10 nationalities are represented in this region, with a strong representation of Malians, Burkinabés, and Guineans. This relative affluence favors the informal sale of antimalarial drugs and promotes abusive self-medication. This is one of the key factors in the emergence of drug resistance (Ministere de l’economie et des finances, 2018).This situation can explain the presence of multiple copies of PfPM2 gene in this area.
Our results show that the multiplications associated with resistance to piperaquine are probably already frequent in Senegal, which is of concern given the use of DHA / PPQ as a second line of treatment in Senegal. Recently complete resistance of P. falciparum following DHA / PPQ treatment has been reported in Cambodia and this resistance has been directly associated with an increase in the number of copies of the PfPM2 2 gene (Amato et al., 2017).
In Mali, in 65 out of the 96 samples it was confirmed the presence of 7 infections carrying 2 copies of PfPM2 (Sanogo et al., 2018). Previously a multiple copies of pfpm2 was also found in Mozambique with a frequency of 1.1% in 351 samples (Gupta et al., 2018).This same situation has already been found in a study carried out in high transmission areas of West Africa (Mali, Burkina Faso and Guinea) by the West African Network for Antimalarial Drugs (Sagara et al., 2018). Another study showed a strong presence of multiple copies of the PfPM2 gene in Uganda with a frequency of 33.9% or 38/112 and a low copy frequency in Gabon and Mozambique with respectively frequencies of 11.3% (8/71) and 12.5% ??(1/8) (Leroy et al., 2019).In Ethiopia and Cameroon only a single copy of PfPM2 was detected in two isolates after DHA-PPQ failures (Russo et al., 2018).Paradoxically, PfPM2 in multiple copies is generally found in parts of Africa where DHA-PPQ failures are rare and resistance to PPQ has not yet been described. However, there is no evidence of PPQ resistance in Senegal although in this study we found a presence of multiple copies of the PfPM2 gene.
Recently, the clinical and parasitological efficacy of DHA/PPQ has been estimated at 97.6% with only 2.5% of individuals experimenting late therapeutic failure (Diallo et al., 2020). The rare cases of failure of PPQ-based treatments described in Africa were not associated with amplification of the PfPM2 gene (Leroy et al., 2019).
However, a new study show that PfCRT Thr93Ser and Ile218Phe mutation have been shown to confer resistance to PPQ without the presence of pfpm2 duplications (Dhingra et al., 2019). Also multiple copies of PfPM2 and pfcrt-F145I were detected in 2014 (12.8%) and increased to 30.4% in 2015. Parasites containing either multiple PfPM2 copies with and without pfcrt-F145I or a single pfpm2 copy with pfcrt-F145I exhibited elevated IC90 values of piperaquine (Boonyalai et al., 2021).
In summary PfPM2 duplications correlates with PPQ resistance, but duplication is not essential for resistance. Therefore, plasmepsin copy number should not be used as a sole indicator of PPQ resistance. Additionally, some polymorphisms in pfcrt can confer resistance to PPQ in Dd2 parasites and the E415G pfexo mutation has been correlated with DHA-PPQ resistance in Cambodian isolates (Moss et al., 2022).
In other words, in Africa it is unclear whether the amplification of PfPM2 is necessary and/or sufficient for the development of resistance to PPQ. These observations suggest that, further molecular surveillance of PPQ resistance including multi-genes will be necessary.
CONCLUSION
The emergence of DHA–PPQ resistance greatly threatens the efficacy of the remaining ACTs worldwide. With the availability of PfPM2 copy number as molecular markers of piperaquine resistance, it is necessary to have robust assays that can be used to monitor the presence and frequency of these markers in contemporary parasite isolates across endemic regions. In this current study we found a presence of multiple copies of PfPM2 gene but the association between these amplifications and a clinical resistance to PPQ was not verifiable. However, an additional functional work is needed to better understand the mechanisms of PPQ resistance and to identify the association of increased piperaquine IC50 values with the copy-number variation. Also a continuous Surveillance for increased PfPM2 copy number could aid malaria control efforts by pinpointing areas where these drugs may be failing like in Senegal.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors appreciate the African Center of Excellence for Genomics of Infectious Disease (ACEGID), the International Centre for Excellence in Malaria Research (ICEMR) project and the Parasitology and Mycology Laboratory Le Dantec Hospital. They also appreciate Younouss Diedhiou, Lamine Ndiaye, Amadou MactarMbaye, Daba Zoumaro for their contribution to the sample collection and patients and staff for their contribution to this study.
REFERENCES
|
Amato R, Lim P, Miotto O, Amaratunga C, Dek D, Pearson RD, Almagro-Garcia J, Neal AT, Sreng S, Suon S, Drury E, Jyothi D, Stalker J, Kwiatkowski DP, Fairhurst RM (2017). Genetic markers associated with dihydroartemisinin-piperaquine failure in Plasmodium falciparum malaria in Cambodia: a genotype-phenotype association study. The Lancet Infectious Diseases 17(2):164-173. |
|
|
Bamba S, Séré A, Nikiéma R, Halidou T, Thiéba B, Dao B, Guiguemdé RT (2013). Traitement préventif intermittent à la sulfadoxine - pyriméthamine du paludisme chez les femmes enceintes: Efficacité et observance dans deux hôpitaux urbains du Burkina Faso. Pan African Medical Journal 14(1). |
|
|
Boonyalai N, Thamnurak C, Sai P, Ta W, Gosi P, Utainnam D, Rodkvamtook W, Kuntawunginn W, Vesely BA, Spring MD, Fukuda MM, Lanteri C, Walsh D, Saunders DL, Smith PL, Wojnarski M, Sirisopana N, Waters NC, Jongsakul K, Gaywee J (2021). Plasmodium falciparum phenotypic and genotypic resistance profile during the emergence of Piperaquine resistance in Northeastern Thailand. Scientific Reports 11(1):1-13. |
|
|
Bopp S, Magistrado P, Wong W, Schaffner SF, Mukherjee A, Lim P, Dhorda M, Amaratunga C, Woodrow CJ, Ashley EA, White NJ, Dondorp AM, Fairhurst RM, Ariey F, Menard D, Wirth DF, Volkman SK (2018). Plasmepsin II-III copy number accounts for bimodal piperaquine resistance among Cambodian Plasmodium falciparum. Nature communications 9(1):1-10. |
|
|
Cheeseman IH, Gomez-Escobar N, Carret CK, Ivens A, Stewart LB, Tetteh KKA, Conway DJ (2009). Gene copy number variation throughout the Plasmodium falciparum genome. BMC Genomics 10(1):1-11. |
|
|
Dhingra SK, Small-saunders JL, Ménard D, Fidock DA (2019). Plasmodium falciparum resistance to piperaquine driven by PfCRT The Lancet Infectious Diseases 19(11):1168-1169. |
|
|
Diallo MA, Yade MS, Ndiaye YD, Diallo I, Diongue K, Sy SA, Ndiaye D (2020). Efficacy and safety of artemisinin-based combination therapy and the implications of Pfkelch13 and Pfcoronin molecular markers in treatment failure in Senegal. Scientific Reports 10(1):1-8. |
|
|
Gupta H, Macete E, Bulo H, Salvador C, Warsame M, Carvalho E, Ménard D, Ringwald P, Bassat Q, Enosse S, Mayor A (2018). Drug-resistant polymorphisms and copy numbers in Plasmodium falciparum, Mozambique, 2015. Emerging infectious diseases 24(1):40. |
|
|
Hastings IM, Hodel EM, Kay K (2016). Quantifying the pharmacology of antimalarial drug combination therapy. Scientific Reports 6(1):1-7. |
|
|
Leroy D, Macintyre F, Adoke Y, Ouoba S, Barry A, Mombo-Ngoma G, Ndong Ngomo JM, Varo R, Dossou Y, Tshefu AK, Duong TT, Phuc BQ, Laurijssens B, Klopper R, Khim N, Legrand E, Ménard D (2019). African isolates show a high proportion of multiple copies of the Plasmodium falciparum plasmepsin-2 gene, a piperaquine resistance marker. Malaria Journal 18(1):1-11. |
|
|
Mbaye A, Gaye A, Dieye B, Ndiaye YD, Bei AK, Affara M, Deme AB, Yade MS, Diongue K, Ndiaye IM, Ndiaye T, Sy M, Sy N, Koita O, Krogstad DJ, Volkman S, Nwakanma D, Ndiaye D (2017). (Plasmepsin II-III copy number accounts for bimodal piperaquine resistance amon. Malaria Journal 16:1-7. |
|
|
Ministere de l'economie et des finances (2018). Rapport De L ' Etude Monographique Sur L ' Orpaillage Au. |
|
|
Moss S, Ma?ko E, Krishna S, Campino S, Clark TG, Last A (2022). How has mass drug administration with dihydroartemisinin ? piperaquine impacted molecular markers of drug resistance?? A systematic review. Malaria Journal pp. 1-23. |
|
|
PNLP-Senegal (2018). Bulletin Epidemiologique Annuel 2017 Du Paludisme Au Senegal Mars 40. |
|
|
Russo G, L'Episcopia M, Menegon M, Souza SS, Dongho BGD, Vullo V, Lucchi NW, Severini C (2018). Dihydroartemisinin-piperaquine treatment failure in uncomplicated Plasmodium falciparum malaria case imported from Ethiopia. Infection 46:867-870. |
|
|
Sagara I, Beavogui AH, Zongo I, Soulama I, Borghini-Fuhrer I, Fofana B, Traore A, Diallo N, Diakite H, Togo AH, Koumare S, Keita M, Camara D, Somé AF, Coulibaly AS, Traore OB, Dama S, Goita S, Djimde M, Bamadio A, Dara N, Maiga H, Sidibe B, Dao F, Coulibaly M, Alhousseini ML, Niangaly H, Sangare B, Diarra M, Coumare S, Kabore MJT, Ouattara SM, Barry A, Kargougou D, Diarra A, Henry N, Soré H, Bougouma EC, Thera I, Compaore YD, Sutherland CJ, Sylla MM, Nikiema F, Diallo MS, Dicko A, Picot S, Borrmann S, Duparc S, Miller RM, Doumbo OK, Shin J, Gil JP, Björkman A, Ouedraogo JB, Sirima SB, Djimde AA (2018). Pyronaridine-artesunate or dihydroartemisinin-piperaquine versus current first-line therapies for repeated treatment of uncomplicated malaria: a randomised, multicentre, open-label, longitudinal, controlled, phase 3b/4 trial. Lancet 391:1378-1390. |
|
|
Sanogo K, Mårtensson A, Sagara I, Björkman A, Veiga MI, Ferreira, PE, Djimde A, Gil JP (2018). Plasmodium falciparum Plasmepsin 2 Duplications, West Africa 24:1591-1593. |
|
|
Saunders DL, Vanachayangkul P, Lon C (2014). Dihydroartemisinin-Piperaquine Failure in Cambodia. New England Journal of Medicine 371(5):484-485. |
|
|
Thiam S, Thwing J, Diallo I, Fall FB, Diouf MB, Perry R, Ndiop M, Diouf, ML, Cisse MM, Diaw MM, Thior M (2012). Scale-up of home-based management of malaria based on rapid diagnostic tests and artemisinin-based combination therapy in a resource-poor country?: results in Senegal. Malaria Journal 11:1. |
|
|
Witkowski B, Duru V, Khim N, Ross LS, Saintpierre B, Beghain J, Chy S, Kim S, Ke S, Kloeung N, Eam R, Khean C, Ken M, Loch K, Bouillon A, Domergue A, Ma L, Bouchier C, Leang R, Huy R, Nuel G, Barale JC, Legrand E, Ringwald P, Fidock DA, Mercereau-Puijalon O, Ariey F, Ménard D (2017). A surrogate marker of piperaquine-resistant Plasmodium falciparum malaria: a phenotype-genotype association study. Lancet Infectious Diseases 17(2):174-183. |
|
|
World Health Organization (WHO) (2018). Artemisinin resistance and artemisinin-based combination therapy efficacy (Status report -- August 2018). |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


