Full Length Research Paper

ABSTRACT
Drilling activities usually discharge large quantities of fluids and drill cuttings, which are composed of fine particles, such as barite, that can physically affect Lophelia pertusa. An experiment to assess the effects of barite smothering on L. pertusa was conducted with a water recirculation system. The corals were collected in Santos Basin, Brazil. The experiment was conducted in 23 L cone-shaped aquaria, adapted with an individual water recirculation system. Each exposure to suspended barite particles (50 and 100 mg•L-1) plus a control group was maintained in triplicate. Three nubbins were accommodated in the aquaria and acclimated for 7 days. The experimental design simulated two cycles of discharge, followed by a recovery cycle of 7 days each, totaling 35 days. Nine polyps per treatment were monitored twice per day regarding their behavior levels. Time-lapse photographs were also taken during the recovery periods. Total polyp survival in the control was 100%, while those in the 50 and 100 mg•L-1 groups were 94.2 and 93.6%, respectively, with no significant difference between treatments. Polyp activity was different between treatments, with both exposed aquaria displaying higher activity than control. This was probably related to the natural cleaning behavior of L. pertusa.
Key words: Barite smothering, sedimentation, impacts of oil and gas industry, Santos Basin, Southeastern Brazil, South Atlantic.
INTRODUCTION
Brazilian offshore oil production is responsible for over 95% of the total oil production in the country, which, according to the Brazilian National Petroleum Agency (ANP), totaled 944,117 million barrels in 2018. The major offshore oil and gas (O&G) fields in the country are located in the southeastern Brazilian continental margin. This area, which comprises the Santos, Campos, and Espírito Santo basins, is responsible for 95% of the offshore oil production in the country (ANP, 2019). As in other parts of the world (Roberts, 2002), offshore O&G areas in Brazil may overlap with deep-sea coral ecosystems.
The distribution of deep-sea corals varies among these Brazilian offshore basins. In the Campos Basin, reef-forming species (Scleractinia) have been observed at depths of 500-1,200 m, predominantly occurring between 500-700 m with some shallower formations. In the Santos Basin, deep-sea corals occur from 200 m, such as Lophelia pertusa (Linnaeus 1758) reefs, although reef-forming species are present in depths up to 1,000 m. In the Espírito Santo Basin, deep-sea coral ecosystems have been found to occur at depths of 450-1,000 m (Cavalcanti et al., 2017, 2019). The species were found to be part of different communities: coral mounds, Lophelia reefs, and in association with authigenic carbonates. The majority of the available data on deep-sea corals in these areas is a result of oil industry efforts. A great deal of information is available regarding the Campos Basin, as major oil production fields have been concentrated in this area for several decades. In contrast, information on the Espírito Santo Basin is poor due to scarce and isolated records. These ecosystems are structurally complex environments that provide habitats for many invertebrate and fish species and offer ideal conditions for settlement, growth, and reproduction (Freiwald et al., 2004).
Among the threats posed to deep-sea coral ecosystems, the oil industry may have an impact, especially through physical and chemical interferences. The installation of subsea structures, such as pipelines and anchoring, is supposed to cause direct physical damage and particle loads through sediment resuspension (DNV, 2013). Deep-sea corals may also be exposed to ongoing discharge from oil production and drilling activities as well as to that of acute events like oil spills (Larsson and Purser, 2011; Purser and Thomsen, 2012; Allers et al., 2013; Järnegren et al., 2017).
In Brazilian offshore operations, the discharges from oil production platforms, such as produced water, are not of real concern for deep-sea coral environments because of the great depth of the water column and the large potential for dispersal. In contrast, drilling activities may pose multiple threats to nearby deep-sea coral communities, such as the potential impacts of the drill cuttings and drilling fluids discharges and anchoring operations that can cause burial, excessive particle loads, and exposure to toxic components (DNV, 2013). Drill cuttings and drilling fluids are directly discharged from the two upper sections of the well (top-hole section) onto the sea floor. After the riser is in place, the drill cuttings and drilling fluids return to the unit for separation. At this point, the drilling fluid can be reused or discharged, and the drill cuttings can be discharged at the sea surface, according to local legislation (DNV, 2013; IOGP, 2016).
To mitigate the risks to deep-sea coral communities, the Brazilian Environmental Agency (IBAMA) determined that the presence of deep-sea corals should be checked within a radius of 500 m from the location of a well, which should be set up at least 100 m away from any coral formation to avoid potential impacts due to drill cutting discharge from the top hole section. If the presence of deep-sea corals is confirmed during the drilling phase with a riser (when returned to the unit), the drill cuttings must be collected and discharged in a predetermined area at a minimum depth of 1,500 m, where the occurrence of deep-sea corals has not been noted in previous environmental characterization projects. In addition, anchor lines may not be positioned on coral formations or moved over them. As such, the use of dynamic positioning drilling rigs is recommended (IBAMA, 2019). The use and discharge of drilling fluids, drill cuttings, complementary fluids, and cements are determined by another IBAMA guideline that regulates their components and ecotoxicity, but there are currently no specific requirements for deep-sea corals.
Data generation and scientific knowledge are essential for guiding the O&G industry and environmental agencies in managing the potential risks to deep-sea corals to ensure the health of these ecosystems. However, there are fewer studies that have focused on deep-sea corals in the Southern Hemisphere compared to those in the Northern Hemisphere. Although studies have been conducted on the Brazilian coast (Viana et al., 1998; Le Goff-Vitry et al., 2004; Pires et al., 2004; Sumida et al., 2004; Castro et al., 2006; Kitahara, 2007; Pires, 2007; Arantes et al., 2009; Kitahara et al., 2009; Cordeiro et al., 2012; Bahr et al., 2020), most have focused on specific taxa or on a compilation of azooxanthellate stony corals, and no studies have focused on maintaining deep-sea corals in the laboratory and subsequent experimentation.
Since 2004, the Brazilian oil company PETROBRAS has been leading research efforts in the Brazilian continental margin and Southwest Atlantic, focusing exclusively on deep-sea corals. These research projects have mapped and characterized the deep-sea coral ecosystems from the Campos Basin by analyzing physical (geological and oceanographic) and biological (biodiversity) aspects (Cavalcanti et al., 2017, 2019). In addition, the company has acquired deep-sea coral data during its offshore operations. In 2016, to further the knowledge of these deep-sea coral communities and improve environmental management tools, the “Marine Sensitive Environments - SENSIMAR Project” was initiated. This project encompasses a broad five-year study that includes the mapping and characterization of benthic and demersal communities as well as an evaluation of the impacts of O&G offshore activities through laboratory and field experiments.
Within the context of the SENSIMAR Project, the present study developed maintenance and experimental systems to assess the impacts of offshore drilling activities on the deep-sea coral L. pertusa. This effort constituted an unprecedented initiative in Brazil, with major initial challenges regarding a lack of research institutes near sources of good quality seawater, which led to the adoption of a recirculating system instead of a flow-through system. L. pertusa was chosen as the test organism because it is the most widely distributed and well-studied structure-forming deep-sea coral, both in Brazil (Pires, 2007; Pires et al., 2015) and worldwide (Freiwald et al., 2004). These hard corals are sessile filter feeders that lacked symbiotic dinoflagellates (zooxanthellae) which live inside most scleractinian corals (Larsson and Purser, 2011). L. pertusa is generally associated to hard substrata and is sensitive to high temperatures, which naturally ranges from 4 to 14 °C, depending on both bathymetric and geographic location (Lunden et al., 2014a).
For this assessment, a focus on the physical impacts of drilling activities was adopted. Drill cuttings may vary due to geological formation characteristics and the drilling fluids used, thus barite (barium sulfate: BaSO4) was considered a good test substance to evaluate the physical impacts of drilling. In addition, as fine sediments tend to have greater effects on corals than do coarse sediments (Erftemeijer et al., 2012), barite was used to represent severe sedimentation conditions. Barite is used in drilling fluid formulations as a weighting agent to control hydrostatic pressure in the well and to prevent uncontrolled flow to the surface (Neff, 2008; 2010; IOGP, 2016). According to IOGP (2016), typical water-based drilling fluids and non-aqueous based drilling fluids may contain ~14 and ~33 wt% (0-50 wt%) of barite, respectively. Barite has low solubility in seawater and is considered to be a chemically inert substance, with its main effects attributed to physical impacts on organisms (Neff, 2008; Bakke et al., 2013). Barite grain size ranges from 6-75 µm (IOGP, 2016), with median grain sizes usually ranging between 10-20 µm (Neff 2008). Barite enters the environment as a result of the discharge of drilling muds or drill cuttings and tends to remain for a long time in the water column while dispersing over wide areas given that it settles slowly because of its small grain size (Lepland and Buhl-Mortensen, 2008; Ellis et al., 2012; Bakke et al., 2013).
Particle sedimentation can cause coral polyps to be either smothered or buried, which may result in both short- and long-term effects, such as tissue damage, decreased growth rates, enhanced mucus production, reduced food intake rates, increased polyp activity, increased respiration rates, and increased mortality under high sediment loads (Weber et al., 2006, 2012; Purser and Thomsen, 2012; Provan et al., 2016). Sediment loads may also adversely affect coral larvae and cause reductions in survival, settlement, and recruitment (Larsson et al., 2013; Järnegren et al., 2017). In a literature review, IOGP (2016) observed that burial effects appear to be less severe as the water depth increases (> 600 m), which allows for a greater dispersion of small particles and leaves thinner layers of drill cuttings near the well location. Thus, the discharge from a top hole section would be more critical for coral formation in Brazilian waters because of their occurrence in deep-water.
Numerical modeling studies of 62 wells located in the Campos and Espírito Santo Basins at different depths were conducted by PETROBRAS to evaluate the dispersion of drill cutting discharge from top hole sections, the results of which supported the regulation that establishes a 100-m no-drill distance from any coral formation. The results showed that a 1 mm thickness was always observed at a maximum distance of 60 m away from the well, with only one result reaching 88 m away from the well (PETROBRAS internal report). Another study in the Campos Basin, which was based on 33 drill cutting discharge numerical modeling studies, also observed reduced areas with deposition layers greater than or equal to 1 mm very close to the wells (Gallo et al. 2006).
The main objective of this study was to evaluate the response of L. pertusa to drilling discharge considering a 100 m distance from the top hole section. In this context, closed systems filled with natural and artificial seawater were tested to maintain L. pertusa in the laboratory, and experimental units with water recirculation were developed to assess the effects of barite smothering. The test concentrations (50 and 100 mg·L-1) considered a deposition layer greater than 1 mm, which took into account the established 100-m minimum distance between a drilling location and a coral formation, and also considered a deposition layer lower than 6.3 mm (Smit et al., 2008), which may result in damage to L. pertusa (Larsson and Purser, 2011). Therefore, this study considered that the highest barite concentration would reduce L. pertusa survival and cause variations in polyp activity and mucus production. This laboratory study, which evaluated both maintenance and experimentation components, on the deep-sea coral L. pertusa constitutes pioneering research in Brazil and was intended to improve the current scientific knowledge on the impacts of O&G offshore activities on these sensitive environments.
MATERIALS AND METHODS
Collection and onboard maintenance
Colonies of white L. pertusa were collected from the Santos Basin in southwestern Brazil using a remotely operated vehicle (ROV). Corals were collected during two different cruises that took place in February 2014 and March 2017 from reefs located between depths of 206-252 m and 207-245 m on the first and second cruises, respectively. Both cruises were carried out in the same location and on the same L. pertusa banks but at different times. The salinity and temperature recorded in the study area varied from 35.4-35.7 PSU and 12-15 °C, respectively, and a pH of 8.1 was recorded in both cruises. Deep-sea coral collections were made under ICMBio/MMA (Instituto Chico Mendes de Conservação da Biodiversidade/Ministério do Meio Ambiente) authorization (921/2018).
Colonies were identified with numbered tags to differentiate among banks/reefs. The colonies were maintained for approximately 2 days in a temporary system composed of three small 60-L tanks that were connected to each other and to a chiller (total volume of 180 L). The system was filled with surface seawater (35 PSU) that had been previously filtered through a 45-µm mesh, sterilized with an ultraviolet lamp, and cooled to sea bottom temperature (~14 °C). Water circulation through the system was driven by a submersible pump (SB1000, Sarlo Better, Brazil) at 1,000 L·h-1.
At the end of the cruise, the corals were taken to the laboratory in thermal boxes (40 and 80 L) that were provided with gentle aeration. Transportation from the ship to the laboratory facility took approximately 1 h. In the laboratory, the corals were gradually acclimatized by replacing 250 mL of field water with the system water. The temperature was stabilized every 30 min over the course of 4 h, as indicated by Lunden et al. (2014b). Colonies from the first cruise were used to establish the maintenance system in the laboratory with closed water circulation, while those from the second cruise were used only once in the barite experiment.
Long-term laboratory maintenance in a closed circulation system
System design
Three systems for L. pertusa maintenance were assembled following the basic principles defined in the procedures adopted for tropical organisms in aquaria. A 700-L system and a 300-L system were filled with natural filtered (0.45-µm mesh) seawater, and a 300-L system was filled with artificial seawater (Pro Reef Sea Salt, Tropic Marin®, Germany). All systems were set to 35 PSU. The system consisted of an upper water reservoir (40 L for the 700-L system and 60 L for the 300-L systems), two holding aquaria (150 L for the 700-L system and 60 L for the 300-L systems), a sump (60 L for the 700-L system and 60 L for the 300-L systems), a lower water container (300 L for the 700-L system and 60 L for the 300-L systems), and two chillers (Figure 1). Water circulation was performed using a submersible pump (4,000 L·h-1 for the 700-L system and 2,500 L·h-1 for the 300-L systems; Multi, SICCE, Italy), located inside the lower water box through PVC pipes. A sequence of 25-, 10-, and 5-µm filters and ultraviolet (UV) lamps (30 W for the large system, and 15 W for the small systems; Cubos, Brazil) were used to eliminate pathogenic bacteria and were installed between the lower water container and the upper tank.

The upper reservoir was made of transparent glass and was used to distribute the water through the two aquaria. The holding aquaria were also made of transparent glass to allow the corals to be visualized and to capture images. In addition, an adjustable power pump (Vortech MP10w, Ecotech, USA) was installed to generate an internal current and an air hose with a porous stone was included to promote constant and smooth aeration. The tanks were covered with white and transparent acrylic covers. Biological filtration was performed inside a sump, which had a division with transparent glass in two compartments. A protein skimmer (model RO-PS 3000Int for the 700-L system and 2000Int for the 300-L systems; Reef Octopus®, China) was used for organic matter removal. Aerobic media were also used for the removal of bacteria (Siporax®, Sera, Germany) and ammonia (Purigem®, Seachem®, USA), and a filter bag (50-200 µm) was placed inside the sump at the end of the drainage system to remove food debris.
A lower water container was used as a reservoir, in which two chillers (1 HP for the larger system and 1/3 HP for the other systems; Gelaqua, Brazil) were kept for temperature maintenance. One chiller circulated the water inside the tank with a submersible pump (SB2000 and SB1000, Sarlo Better, Brazil), and the second chiller was part of the main system circulation. Aragonite (3 kg) was placed inside the container as a substrate for anaerobic bacteria. The systems were maintained in a cold chamber at ~12 °C in darkness, which was equipped with back-up power generators. The stabilization of the nitrogen products (nitrate, nitrite, and ammonia) in water was achieved after a biological filter maturation period (approximately 1 month), which was stimulated by adding nitrifying and denitrifying bacteria (Biodigest, Prodibio, France) at the sump and a source of organic matter (Artemia sp. nauplii) in the holding aquaria.
Coral maintenance
The corals collected during the first cruise (February 2014) were first introduced into system 1 (700 L of natural seawater). After 10 months, they were transferred to system 2 (300 L of natural seawater) and maintained in this system for another two years. After which, they were transferred to an alternative system and maintained for another year. The corals collected in the second cruise (March 2017) were organized into three systems, including system 3 (300 L artificial seawater), and were maintained until December 2017, when the maintenance systems were discontinued. Fragments of the coral colonies were vertically attached to suction cups, and three fragments were selected to be monitored as sentinels for survival, integrity, and polyp behavior (Figure 1). Freshly hatched (< 24 h) Artemia sp. nauplii were provided three times a week at a rate of 300 nauplii per polyp per day (as recommended by Larsson et al., 2013; Provan et al., 2016). Aquarium bottom siphoning to remove food scraps was performed once per week with 10% water volume renewal to maintain water quality.
Water quality parameters were monitored and adjusted when necessary to keep them close to in situ conditions. Data of pH, salinity, dissolved oxygen, and conductivity were collected continuously from June 2015 to December 2016 in system 2 and from March to June 2017 in systems 1, 2, and 3 using a multiparameter probe (Professional Plus, YSI®, USA). Ammonia levels were monitored using an ISE/pH meter (Thermo Orion, USA), and nitrite and nitrate levels were measured using a colorimeter (Smart3, LaMotte, USA) and were assessed from November 2015 to February 2017 in system 2 and from March to June 2017 in all three systems. Other parameters, such as potassium, alkalinity, magnesium, and calcium content were monitored once a week with aquarium kits (Marine Care Multi Test Kit, Red Sea, USA), but these results are not presented in this paper. When necessary, adjustments were made in the sump to maintain the water parameters within the desirable levels. Salinity was adjusted with purified water by reverse osmosis (RO water), while pH adjustments were made with Kalkwasser (Two Little Fishies®, USA) and alkaline buffer ® (Seachem®, USA).
Exposure experiment with barite
Unit design
The study was designed to evaluate the effects of suspended and smothering barite particles on L. pertusa and on the recovery time. The experimental unit was based on the model proposed by Brooke et al. (2009) and Baussant et al. (2018), which was developed and refined through a series of preliminary tests. The unit was composed of a 20-L cone-shaped aquarium and a sump, with main closed circulation and auxiliary flow-through circulation. A drain was installed at the bottom of the cone, allowing the water to be drained, simulating vertical flow. The cone-shaped aquaria were made out of PVC and transparent acrylic to enable images to be captured (Figure 2). The sump was a 3-L glass vessel, which contained a submersible pump (SB1000, Sarlo Better, Brazil), and was responsible for the main circulation at a flow rate of 12 L·h-1. To buffer heat from the pump and maintain the desired temperature, the sump was maintained in a cooling bath system that was composed of two plastic containers filled with RO water that were connected to each other and to two chillers (1/3 HP).

Barite addition
The definition of the concentrations tested in the present study considered a deposition layer greater than 1 mm, which considered the established minimum 100-m distance between drilling activities and a coral formation, and lower than 6.3 mm (Smit et al. 2008). This may result in damage to L. pertusa, according to Larsson and Purser (2011). Therefore, concentrations of 50 and 100 mg·L-1 were chosen to represent the 3 and 5-mm barite layers, respectively, in the flat area of the experimental unit after one cycle of continuous exposure. Given the insoluble and highly dense nature of barite during the previous tests, a decay of approximately 20% of the initial concentration in 1 h was observed, and the continuous addition of barite was required to maintain the desired suspended concentrations. Barite was added continuously from a stock solution at the top of the aquarium using a 24-channel peristaltic pump (520s, Watson-Marlow, UK) at a flow rate of 0.18 L·h-1. The drained water was stored in 40-L reservoirs for later disposal. The barite concentrations in the reservoir were calculated considering losses along the system. A stock of clean water was also maintained for control and aquarium water exchange during the recovery period.
Experimental set-up
After 80 days of acclimatization in the maintenance system, the corals were used in the barite experiment. They were fragmented into nubbins with three to eight polyps each. Three L. pertusa fragments from the second cruise were vertically positioned in each exposure aquarium, approximately 40 cm below the water surface, and left to acclimatize to the experimental conditions for 7 days until barite addition. Treatments of 50 and 100 mg·L-1 and a control group with artificial seawater were established in triplicate (Figure 3). The amount of barite, defined in the previous tests for each concentration, was diluted in artificial seawater and added directly to the reservoir (40 L) of each experimental unit. A total of 1.22 and 2.7 g of barite were used for the 50 and 100 mg·L-1 concentrations, respectively.

The initial concentrations for the replacement stock solutions were 3.36 and 6.0 g·L-1 for 50 and 100 mg·L-1, respectively. Three different pumps were installed in each reservoir to keep the barite in suspension and ensure aeration of the solutions: 50 mg·L-1 (2,500 L·h-1; Multi, SICCE, Italy), 100 mg·L-1 (4,000 L·h-1; Multi, SICCE, Italy), and control (1,600 L·h-1 JP-025, Sunsun, China). During the exposure period, it was necessary to adjust the desired barite concentrations, which were made in 40-L reservoirs, by the addition of seawater or barite. For the control group, clean artificial seawater (35 PSU) was added to the reservoir (40 L).
A transition between the exposure and recovery periods was performed by exchanging the used hoses connected to the exposure aquaria with clean hoses connected to the clean water reservoir at the end of the last day of exposure. The renewal of clean water in the aquaria during the recovery periods was performed at a flow rate of 1.26 L·h-1 for 3 h per day, allowing 16.4% of the total water volume to be renewed in all exposure aquaria during the 7-day cycle. The experiment was kept in a cold chamber set to 12.5 °C in darkness. Artificial seawater at 35 PSU was used. The corals were fed newly hatched (< 24 h) Artemia sp. nauplii (300 nauplii·L-1·day-1) three times per week.
The total experimental duration was 35 days, with one acclimatization cycle, two exposure cycles, and two recovery cycles, each lasting 7 days (Figure 3). After 7 days of acclimatization in the exposure aquaria, activity was observed in all polyps, and the simulation with barite was initiated. This discharge attempted to mimic a drilling regime, although each drilling operation has specific characteristics according to the reservoir features and well design. At the end of the experiment, all fragments were transferred to a new aquarium and kept for observation.
The exposure system presented in this study allowed for a total volume of approximately 646 L of synthetic seawater to be used during the 35 days of the experiment. Circulation within one experimental unit required 23 L of synthetic seawater. During the acclimatization period, 16% of the total volume was manually changed once a week (34 L for the nine exposure replicates). The estimated volume of seawater used during the exposure period in each experimental unit, which was continuously pumped by the peristaltic pump, was 30.24 L, with a total of 272.16 L over 7 days. Considering the two exposure cycles (total of 14 days), the final volume was 544.32 L.
Water temperature, salinity, pH, and dissolved oxygen data were recorded daily using a digital thermometer (Instrutherm, Brazil), a digital refractometer (Atago, Japan), a pH meter (Orion Star A211, Thermo Scientific, USA), and a dissolved oxygen meter (Orion Star A213, Thermo Scientific, USA), respectively. Ammonia, nitrite, and nitrate concentrations were checked three times per week at the beginning, middle, and end of each exposure and recovery period. An electrode (Thermo Orion, USA) was used to measure the ammonia concentration, and a colorimeter (Smart 3, LaMotte, USA) was used to measure the nitrite and nitrate concentrations. The maintenance levels for water quality were used as a reference for the exposure system.
Three water samples were collected every day from each aquarium throughout the experiment to determine the barite concentrations using a turbidimeter (TD-300, Instrutherm, Brazil). The results of these readings, given in Nephelometric Turbidity Units (NTU), were converted to mg·L-1 using a calibration curve assembled from five preset barite concentrations. The average concentration was calculated. On the first day of exposure, samples were taken every hour to monitor the variations in the concentrations of each experimental unit to make any necessary adjustments. In the following days, analyses were performed three times per day (09:00, 13:00, and 17:00 h).
To evaluate the effects of barite smothering with the selected concentrations, three endpoints were observed: survival, mucus production, and polyp behavior. Polyp survival was verified at the end of the second recovery cycle, and mucus production was observed during the recovery cycles. To evaluate polyp behavior, time-lapse photos were taken at 30-min intervals from one replicate of each treatment. Two Nikon D90 cameras (Tokyo, Japan) coupled to two computers with Nikon Camera Control Pro 2 software were used, with one focused on the control and 50 mg·L-1 treatments and the other focused on the 100 mg·L-1 treatment. In addition to the time-lapse photos, polyp behavior was also analyzed by daily observations, similar to those performed by Larsson et al. (2013) and Orejas et al. (2016). Three polyps from each fragment (nine polyps per aquarium) were monitored throughout the experiment for activity. They were photographed twice per day at fixed times (10:00 and 15:00 h) and classified according to the opening of the polyps: (1) fully or half-opened/extended polyp or (0) fully closed/retracted polyp (Figure 4). Mucus production was observed but not quantified. The time-lapse photos were constrained to one replicate of each treatment because of physical space limitations given that the equipment (camera, tripod, and PC) requires sufficient room for optimal picture capture without compromising clarity. For daily observations, all replicates were analyzed without any such compromises.

Data analyses
To evaluate the variation in the barite concentration throughout the exposure cycles, a time series graph was constructed for each exposed treatment with all replicates. These graphs were made using Microsoft® Excel (USA). To test for differences in polyp survival and polyp activity by observing corals that had been exposed to different treatments, data based on a binary response (dead × alive, extended × retracted) were analyzed using a Kruskal-Wallis rank test because of the nonparametric nature of the data. To test for normality and homoscedasticity, Shapiro-Wilk and Bartlett tests were used, respectively. The data did not meet these assumptions, even after arc-sin transformation. All analyses were performed in R v. 3.5.2 (R Core Development Team, 2018). In the case of significant results, a post-hoc nonparametric multiple comparison analysis was used. These analyses were performed using the ‘nparcomp’ package in R (Konietschtke et al., 2015). For polyp activity, a box-plot of the proportion of extended polyps was created with the data from the entire experiment. In addition to the boxplot, time series graphs of the percentage of extended polyps were plotted for each replicate of all treatments. All plots were created using Microsoft® Excel.
Time-lapse photos were used for polyp activity analysis that analyzed pixel area variation (Baussant et al., 2018). Corel Paint Shop Pro X9 software was used to treat the photos, and ImageJ (Rasband, 2015) software was used to measure the pixel area. To process all photos, the batch process was used in both programs. These data were plotted in a time series graph for each fragment to evaluate variation throughout the experiment.
The time-lapse data were also analyzed to compare polyp behavior in both recovery cycles, despite being from only one replicate of each treatment. The time-lapse data displayed a nonparametric distribution, even after arc-sin transformation, as was found with observational polyp behavior data. A Kruskal-Wallis rank test was used, and in case of differences between treatments, nonparametric multiple comparisons with the R package ‘nparcomp’ were done. The differences between treatments in both recovery cycles were evaluated as a whole as well as in each recovery cycle, and the recovery differences between each exposed treatment were also evaluated. For comparisons between cycles for each exposure treatment, a Wilcoxon signed-rank test was conducted. All analyses were conducted considering a 95% confidence level. The differences between
RESULTS
Long- and short-term maintenance in a closed water circulation system
L. pertusa specimens were kept alive for 35 months in natural seawater systems with different volumes. The physical and chemical parameters were within the limits established for the species and for other corals, which reflected the stability of the maintenance system. The results presented in this study considered the YSI® parameters and nutrient contents acquired during the continuous monitoring periods. The 300-L natural seawater system, which was operational for the longest period (35 months), presented (mean ± SD) variations in temperature, salinity, pH, and dissolved oxygen of 10.9 ± 0.2 °C, 35.16 ± 0.53 PSU, 8.09 ± 0.23, and 7.69 ± 0.54 mg·L-1, respectively (Table 1). The nutrient (ammonia, nitrite, and nitrate) concentrations were below critical values (< 0.1 mg·L-1 for ammonia and nitrite and < 2.0 mg·L-1 for nitrate; Table 1).

Three systems, 700 and 300 L of natural seawater and 300 L of artificial seawater were used for the short-term maintenance of corals during the 80-day acclimatization period (March-June 2017). The YSI® records resulted in 1,716, 1,672, and 1,743 data points for each parameter for systems 1, 2, and 3, respectively, with the temperature, salinity, dissolved oxygen, and pH ranging from 10.6-13.10 °C, 34.2-37.0 PSU, 3.59-8.04, and 7.53-8.67, respectively. The average nitrite, nitrate, and ammonia values in the three maintenance systems were within the reference limits for seawater aquaria (nitrite < 2 ppm; nitrate < 0.2 ppm; ammonia < 0.1 ppm; Table 2). The values of the physical and chemical parameters (Table 2) also remained within the limits for coral survival, with no differences between the natural and artificial seawater systems (systems 2 and 3, respectively). Survivorship was 93, 80, and 83% in systems 1, 2, and 3, respectively. The high polyp survival rate (≥ 80 %) allowed for the polyps to be used in the experiment.

Barite exposure experiment
Suspended barite concentrations
The mean barite values for the 50 mg·L-1 treatment replicates (R) during the first cycle were 46.76 ± 16.26 (R1), 50.77 ± 19.19 (R2), and 47.51 ± 20.97 mg·L-1 (R3). During the second cycle, the concentrations were 59.29 ± 37.78 (R1), 61.12 ± 31.85 (R2), and 60.07 ± 37.11 mg·L-1 (R3; Figure 5). The second cycle showed greater variability in barite concentrations. In aquaria receiving 100 mg·L-1 barite, the mean concentrations were 109.98 ± 37.08 (R1), 92.56 ± 34.76 (R2), and 127.46 ± 41.96 mg·L-1 (R3) during the first exposure cycle. During the second cycle the mean concentrations were 192.2 ± 121.58 mg·L-1 (R1), 172.92 ± 76.88 mg·L-1 (R2), and 170.53 ± 78.26 mg·L-1 (R3; Figure 5). Greater variation during the second exposure cycle was also observed. The mean barite concentrations were 7% above the expected concentration of 50 mg·L-1 treatment and 40% above the 100 mg·L-1 treatment, with coefficients of variation of 52 and 55%, respectively.

Water quality parameters
Stable water conditions were maintained in the exposure aquaria. These parameters were also maintained within the maintenance levels (Figure 6). Total variation within the nine exposure units included temperature, salinity, dissolved oxygen, and pH values between 11.6-13.6 °C (12.46 ± 0.32°C), 34-36 PSU (35 ± 1 PSU), 4.27-8.52 mg·L-1 (6.51 ± 0.97 mg·L-1), and 7.48-8.60 (7.88 ± 0.21), respectively. Nutrient concentrations varied from 0.0012-0.35 mg·L-1 (0.0344 ± 0.0405 mg·L-1), 0-0.489 mg·L-1 (0.0489 ± 0.0721 mg·L-1), and 0.284-1.7850 mg·L-1 (0.4568 ± 0.2231 mg·L-1) for ammonia, nitrite, and nitrate, respectively, in the nine exposure units. Regarding the physical and chemical parameters, there were no differences among the control and 50 and 100 mg·L-1 treatments (Figure 6).

L. pertusa responses to barite
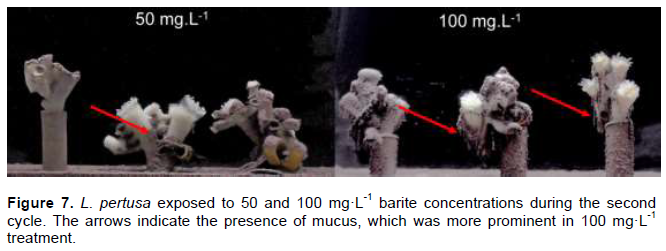
During the 7-day acclimatization period inside the exposure aquaria, no mortality was observed. Polyp survival in the control group was 100% after 35 days. Survival in the 50 and 100 mg·L-1 treatments was 94.23 and 93.62%, respectively, with no significant difference between the treatments and the control group (Kruskal-Wallis H = 0.3506, p = 0.84). The amount of settled barite varied among fragments in the same treatment because of the particular three-dimensionality of the L. pertusa fragments, and between treatments, as a higher barite concentration could imply a thicker layer. At the end of the experiment, all corals were transferred to clean seawater and monitored for 47 days, with no observed mortality. Greater mucus production was observed in the 100 mg·L-1 treatment, although this was not quantified (Figure 7).
Time-lapse results
A total of 1,571 time-lapse photos were analyzed for each treatment. The amount of suspended barite in the exposure aquaria interfered with the capture of some images. Thus, except for the control, the activity graphs were constructed considering only the acclimatization and recovery periods, during which the corals were exposed to clean seawater. In the control, polyp activity was observed throughout the experiment.
The time-lapse results included three fragments of only one replicate (R1) from the control and 50 and 100 mg·L-1 treatments. No records or pictures were taken during the exposure cycles because of the presence of barite in the water. The control fragments showed constant activity during the 35 days of the experiment, with some prominent peaks that were not necessarily related to feeding events. Fragments of the 50 mg·L-1 treatment, especially in systems 2 and 3, showed a considerable increase in activity after exposure compared to those in the acclimatization period. For these fragments, the activity after exposure was also greater than that of the control. Polyps from fragment 1 in the 100 mg·L-1 treatment presented low activity during the acclimatization period (lower than that of the control) and died during the first exposure cycle. Fragments 2 and 3 exposed to the 100 mg·L-1 treatment showed greater activity than those in the control and showed similar activity to that of the acclimatization period. Fragments from the 50 and 100 mg·L-1 treatments seemed to have similar increases in activity after exposure (Figure 8), with slightly higher activity in the 100 mg·L-1 treatment (Figure 9) compared to that of the control.


A comparison of both recovery cycles as a whole showed significant differences among treatments (Kruskal-Wallis chi-squared = 395.53, df = 2, p = 0), with the post hoc analysis indicating differences among all comparisons. For the first recovery cycle, significant differences were also present (Kruskal-Wallis chi-squared = 201.23, df = 2, p = 0). However, in this case, the post hoc analysis only indicated a difference when comparing the exposure treatments and the control; no difference was detected between the 50 and 100 mg·L-1 treatments. The second recovery cycle also displayed significant differences (Kruskal-Wallis chi-squared = 153.19, df = 2, p = 0), with all treatments being different from each other. The analysis for the 50 mg·L-1 treatment indicated a significant difference between cycles (V = 18,065, p = 0.0008537), with the second recovery cycle displaying higher polyp activity than that of the first (Figure 10). In contrast, the 100 mg·L-1 treatment displayed no differences between cycles (V = 22,420, p = 0.3024; Figure 10).

Direct observations
A total of 504 photos were analyzed for direct observation analysis. The direct observation results included three replicates (with three L. pertusa fragments each) of each treatment (control, 50 mg·L-1, and 100 mg·L-1), with records for the exposure cycles but not for the acclimatization period. Replicates of the control showed continuous activity, with some peaks that were not necessarily related to the feeding stimulus. Replicates of the 50 mg·L-1 treatment showed greater activity than those of the control. For R1, there was an evident increase in activity after the first exposure cycle, while for R2 and R3, activity remained high during both the exposure and recovery cycles. Replicates of the 100 mg·L-1 treatment also showed a greater activity than that of the control. Polyp activity increased after the first exposure cycle and remained high until the end of the test for R1 and slightly decreased in the second cycle for R2 and R3 (Figure 10). Polyp activity showed significant differences among treatments (Kruskal-Wallis H = 142.83, p = 0), with post hoc tests showing differences among all comparisons. The proportion of extended polyps varied between 0-0.33 (0.07 ± 0.1), 0-1 (0.32 ± 0.22), and 0-0.89 (0.23 ± 0.21) for the control, 50 and 100 mg·L-1 treatments, respectively, with greater activity observed in the 50 mg·L-1 treatment, and the lowest activity observed in the control. Feeding can stimulate polyp activity; however, a direct relationship between a stimulus (food) and response (activity) was not observed.
DISCUSSION
An evaluation of the tolerance limits of deep-sea corals to different substances allows for criteria and mitigation measures to be established by industries and agencies. However, an interpretation of the available data of the effects of suspended material on deep-sea corals is difficult because of the absence of standardized laboratory protocols. In addition, the high cost and complexity involved in deep-sea coral maintenance under artificial conditions and in conducting experiments with these organisms limits the number of laboratories that may be involved in this type of research. Given these factors, the data available in the literature are the results of experiments carried out at a base level and that have been adapted to the conditions of each laboratory. Therefore, these obstacles (particularly laboratory conditions as well as the absence of standardized protocols) prevent interlaboratory experiments to be conducted and limit comparisons among results. Some of the main achievements of this study were to test different maintenance conditions in closed water systems and to further define protocols for subsequent maintenance and exposure experiments.
In addition, access to seawater is another limiting factor that limits this type of research, especially for laboratories located far from the sea. This implies high costs for the purchase of natural seawater or artificial sea salt, which greatly interferes with the final cost of a research project. Thus, it is extremely important to identify systems that use the smallest possible amounts of seawater without interfering with the results of the experiments. This goal was likewise achieved with the maintenance and exposure systems designed in this study.
Long- and short-term maintenance in a closed water circulation system
Flow-through systems are best suited for the long-term maintenance of deep-sea corals. These systems do not require intense filtration processes. However, they still use physical filters (bags or sand filters) and UV sterilization (Orejas et al. 2019). Some systems may have hybrid flow, with the possibility of operating with both flow-through and recirculating flow, such as the aquaria system of the Instituto de Ciencias del Mar (ICM-CSIC), which was described by Olariaga et al. (2009), and that of the Instituto do Mar (IMAR)/University of Azores, which was described by Orejas et al. (2019). Despite this ability, continuous flow is the operational standard, with recirculating flow being used only in cases when the supply of seawater is interrupted.
The maintenance of deep-sea corals under optimum water quality conditions is not an easy task, especially in closed water systems that require a precise control of nitrogen products and micronutrient concentrations. Although temperature is also a critical parameter for these organisms, it can be controlled by keeping the systems connected to chillers and maintaining them in acclimated rooms equipped with back-up power generators. Recirculating systems are often used for short-term experiments (Hennige et al., 2015; Gori et al., 2016; Orejas et al., 2016), but for long-term maintenance, flow-through systems are preferable (Orejas et al., 2019).
Few studies have been performed on the long-term maintenance of deep-sea corals using closed water systems. Lunden et al. (2014b) presented a recirculation aquarium model that enabled the maintenance of L. pertusa for four years, which was composed of mechanical, chemical, and biological filtration. In contrast to the system presented in this paper, Lunden et al. (2014b) built a biological filter using the Jaubert plenum model with crushed coral and sand and introduced macroalgae to aid in nitrate removal. Despite these differences, the biological filters and procedures adopted in this study ensured that the monitored parameters were maintained within the desired limits for L. pertusa maintenance in the systems that were operated with natural seawater, independent of the capacity (300 or 700 L), and with artificial water, when comparing both 300-L systems. In general, compared to open systems, recirculation systems always require constant maintenance and a small amount of water renewal to ensure low concentrations of nitrogen products and to maintain trace levels of metals and minerals.
The maintenance procedures in this study ensured water quality and stability in the systems, with minimum temperature variations. Polyps were active during the entire maintenance period in the systems with both natural (35 months) and synthetic (10 months) seawater. Polyp behavior showed that the colonies had acclimated to laboratory conditions. Despite the brief study period, the use of synthetic water showed positive results with L. pertusa, and the recirculating system proved to be adequate for maintaining L. pertusa in the laboratory, as seen in Lunden et al. (2014b).
Barite exposure experiment
The cone-shaped aquaria used herein were adapted from models proposed by Brooke et al. (2009) to evaluate the effects of different sediment concentrations on two L. pertusa morphotypes and were also used by Baussant et al. (2018) to test the effects of drill cuttings on L. pertusa in pulsed and continuous exposure scenarios. As the authors used a continuous flow system, it was necessary to adapt the recirculating system to achieve continuous exposure without excessive water utilization. Thus, the addition of secondary flow-through circulation, at a much lower flow rate than that of the main circulation, was necessary to add barite to the exposure unit. In addition, routines employed for the maintenance systems were also necessary to ensure adequate water quality.
Barite loss occurred throughout the exposure system, mainly due to adherence to hoses and sedimentation. However, barite suddenly returned to the system when it came off the hoses, causing an increase in concentration, which was probably one of the factors that led to the large variation observed around the desired concentrations (50 and 100 mg·L-1), especially during the second exposure cycle. In addition, the positioning and distance of the hoses in the system and the efficiency of the peristaltic pump may also have contributed to this variation. Despite the high variation, the recirculated system designed for this experiment was adequate for barite exposure.
The effect of sediment coverage on deep-sea corals is not well understood (Allers et al., 2013). In laboratory experiments, fragments of L. pertusa were relatively tolerant to high sediment concentrations (Brooke et al. 2009; Larsson and Purser, 2011; Allers et al., 2013), low oxygen levels (Dodds et al., 2007; Allers et al. 2013), and burial (Brooke et al., 2009; Allers et al., 2013). In a laboratory study on L. pertusa from the Gulf of Mexico, Brooke et al. (2009) tested the tolerance of two morphotypes to different sediment loads as well as the response to complete short-term sediment coverage. The authors showed that L. pertusa could tolerate a fairly high short-term sediment coverage of 2-4 days before coral mortality set in and suggested that mortality was most likely due to oxygen deficiency. Sub-lethal effects on L. pertusa from exposure to both benthic sediments and drill cuttings, on the other hand, included tissue loss (Larsson and Purser 2011), reduced skeletal growth, and increased larval mortality, which depended on the concentration used and larval age (Larsson et al., 2013; Järnegren et al., 2017).
Larsson and Purser (2011) found that L. pertusa could survive repeated, slight smothering by natural sediments and drill cuttings, but death occurred when the polyps were wholly covered by the particles. The response to cuttings and natural sedimentation did not differ. It was concluded that the current effect levels from the non-toxic burial of 6.3 mm proposed by Smit et al. (2008) may result in damage to L. pertusa colonies. Allers et al. (2013) showed that the complete burial of coral branches for > 24 h in reef sediments resulted in suffocation, and that L. pertusa fragments suffocated only under 20× coverage (1,320 mg·cm-2) and not among tested nominal sediment coverages of 1× (66 mg·cm-2), 3× (198 mg·cm-2), or 7× (462 mg·cm-2). These experimental exposures were of short duration (12 h to 4 days) and only assessed tissue damage and survival. A longer experiment (12 weeks) that exposed L. pertusa to chronic and moderate (25 ppm) levels of natural sediments and drill cuttings resulted in high survival but reduced growth rates, particularly with suspended drill cuttings (Larsson et al. 2013). In addition, Baussant et al. (2018) tested different scenarios with drill cuttings that consisted of a long-term pulsed exposure (12 weeks, mean concentrations: 1-25 mg·L-1) and a short-term continuous exposure (2.5 weeks, mean concentrations: 4-42 mg·L-1), followed by 14 and 4 weeks of recovery, respectively. They observed a significant increase in skeleton growth at 4 mg·L-1 and a significant increase in mucus particulate organic carbon at 25 mg·L-1 at the end of the exposure time. In situ observations have also shown that drilling operations have little direct impacts on L. pertusa reefs exposed to low levels (< 25 ppm) of material (Buhl-Mortensen et al., 2015; Purser, 2015). Video monitoring carried out during discharge episodes in Norway revealed no significant behavioral differences between exposed and unexposed L. pertusa polyps (Buhl-Mortensen et al., 2010). Polyp retraction responded more systematically to changes in current velocity and direction than to cutting plumes.
Regarding the evaluation of polyp behavior, direct observations and time-lapse photography approaches seem to be complementary, which each approach presenting advantages and disadvantages. For instance, direct observation allows for continuous records, including during barite exposure, but interpretation can vary according to the observer, whereas time-lapse photography generates more comparable results since the analysis can be automated but does not allow for records to be taken during barite exposure cycles due to water turbidity. With both methods, higher polyp activity has been observed in the exposed aquaria, which is probably related to the natural cleaning behavior of L. pertusa.
Baussant et al. (2018) also observed that drill cuttings produced an increase in coral polyp activity during exposure. They noticed a return to pre-exposure conditions after the cessation of treatment, while the coenosarc was smothered as a result of exposure, even after 4 weeks of recovery. The authors noted these changes at a concentration of 10 mg·L-1, which seems to represent a threshold at which the coral status is modified compared to that of the natural environment. This concentration also corresponds to the tolerance threshold reported for dredging particles in shallow-water corals (Rogers 1990; Browne et al., 2015) and to the current threshold level of suspended particles used in general risk assessment models (Larsson et al., 2013). In this study, the amount of settled barite varied among fragments in the same treatment and between treatments, which may have caused the smothering and death of polyps. Nevertheless, polyp survival in this study demonstrated the tolerance of L. pertusa to barite concentrations greater than those observed in other studies, including for sessile sediment species in the environment (0.2 mg·L-1; Smit et al., 2008) and for the threshold of suspension particles in risk assessment models (10 mg·L-1; Rye et al., 2011). Therefore, barite had no lethal effect on L. pertusa under these test conditions, as polyp survival was more than 90% after 35 days of the experiment.
Even though there were no time-lapse records after 35 days, the polyps remained alive and active for more than 45 days after the end of the barite test. Therefore, L. pertusa showed tolerance to barite sedimentation at concentrations of 50 and 100 mg·L-1. Although long-term exposure to natural sediment is known to have energetic costs for corals and negative consequences, such as coenosarc damage and slower skeletal growth (Rogers, 1990; Weber, 2009; Erftemeijer et al., 2012; Baussant et al., 2018), these endpoints were not assessed in this study. Although it is not regularly quantified, mucus production was observed after the first discharge cycle in both treatments in this study and was not observed in the control. This may also have been possibly associated with the coral cleaning process and has already been reported by other authors (Larsson and Purser, 2011; Allers et al., 2013; Larsson et al., 2013; Baussant et al., 2018).
CONCLUSION
The results of the present study indicate that the mortality of cold-water coral polyps due to discharge from drill cuttings at distances of at least 100 m from the location of a well is not expected. It is very important that clear boundaries be established for oil and gas activities in areas near coral banks. As such, other studies are being planned to test the effects of natural sediments and drilling waste on threshold levels and the environmental risks of discharge at sea in typical drilling scenarios and those of related activities. In addition, other endpoints must be monitored in future experiments to evaluate the tolerance of L. pertusa and other deep-sea coral species to drilling discharge considering short and long terms.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGMENTS
The authors are grateful to Thierry Baussant and Steinar Sanni from NORCE (Norway) for kindly helping to define the conditions of coral exposure to barite and for assisting in the use of programs for image processing. They thank Marcia J. C. Botelho for her help in defining the design of the maintenance and exposure systems, Rafaela Costa and Adriano R. Machado for technical assistance throughout the exposure experiment, Sônia L. R. de Mello for her thoughtful comments and suggestions on the experimental design, and Teresinha Silva for her comments on the manuscript. They are grateful to PETROBRAS for funding the study in the context of the SENSIMAR Project.
REFERENCES
|
Agência Nacional do Petróleo (ANP) (2019). Gás Natural e Biocombustíveis, Anuário estatístico brasileiro do petróleo, gás natural e biocombustíveis: |
|
|
Allers E, Abed RMM, Wehrmann LM, Wanga T, Larsson AI, Purser A, de Beer D (2013). Resistance of Lophelia pertusa to coverage by sediment and petroleum drill cuttings. Marine Pollution Bulletine 74:132-140. |
|
|
Arantes RCM, Castro CB, Pires DO, Seoane JCS (2009). Depth and water mass zonation and species associations of cold-water octocoral and stony coral communities in the southwestern Atlantic. Marine Ecology Progress Series 397:71-79. |
|
|
Bahr A, Doubrawa M, Titschack J, Austermann G, Nürnberg D, Albuquerque AL, Friedrich O, Raddatz J (2020). Monsoonal forcing controlled cold water coral growth off southeastern Brazil during the past 160 kyrs. Biogeosciences Discussions |
|
|
Bakke T, Klungsøyr J, Sanni S (2013). Environmental impacts of produced water and drilling waste discharges from the Norwegian offshore petroleum industry. Marine Environmental Research 92(2013):154-169. |
|
|
Baussant T, Nilsen M, Ravagnan E, Westerlund S, Ramanand S (2018). Effects of suspended drill cuttings on the coral Lophelia pertusa using pulsed and continuous exposure scenarios. journal of Toxicology and Environmental Health, Part A 10:361-382. |
|
|
Brooke SD, Holmes MW, Young CM (2009). Sediment tolerance of two different morphotypes of the deep-sea coral Lophelia pertusa from the Gulf of Mexico. Marine Ecology Progress Series 390:137-144. |
|
|
Browne NK, Tay J, Todd PA (2015). Recreating pulsed turbidity events to determine coral - sediment thresholds for active management. Journal of Experimental Marine Biology and Ecology 466:98-109. |
|
|
Buhl-Mortensen P, Klungsøyr J, Meier S, Purser A, Tenningen E, Thomsen L (2010). Environmental Monitoring Report, Morvin 2009-2010. Bergen, Norway: Institute of Marine Research. |
|
|
Buhl-Mortensen P, Tenningen E, Skar Tysseland AB (2015). Effects of water flow and drilling waste exposure on polyp behaviour in Lophelia pertusa. Marine Biology Research 11(7):725-737. |
|
|
Castro CB, Amorim LC, Calderon EN, Segal B (2006). Cobertura e recrutamento de corais recifais (Cnidaria: Scleractinia e Milleporidae) nos recifes Itacolomis, Brasil. Arquivos do Museu Nacional 64(1):29-40 |
|
|
Cavalcanti GH, Arantes RCM, Falcão APC, Curbelo-Fernandez MP, Silveira MAS, Politano AT, Viana AR, Hercos CM, Brasil ACS (2017). "Ecossistemas de corais de águas profundas da Bacia de Campos" in: Comunidades Demersais e Bioconstrutores: caracterização ambiental regional da Bacia de Campos, Atlântico Sudoeste, eds. M. P. Curbelo-Fernandez and A. C. Braga (Rio de Janeiro: Elsevier). Habitats 4:43-85. |
|
|
Cavalcanti GH, Curbelo-Fernandez MP, Arantes RCM, Loiola LL, Falcão APC, Lima AC (2019). Deep-sea coral ecosystems off southeastern and Southern Brazilian slope in Abstract Book, International Symposium on Deep-Sea Corals. |
|
|
Cordeiro RTS, Kitahara MV, Amaral FD (2012). New records and range extensions of azooxanthellate scleractinians (Cnidaria, Anthozoa) from Brazil. Marine Biodiversity Records 5:e35. |
|
|
Det Norske Veritas (DNV) (2013). Monitoring of drilling activities in areas with presence of cold water corals. DNV Report nr 012-1691 |
|
|
Dodds LA, Roberts JM, Taylor AC, Marubini F (2007). Metabolic tolerance of the cold-water coral Lophelia pertusa (Scleractinia) to temperature and dissolved oxygen change. Journal of Experimental Marine Biology and Ecology 349(2):205-214. |
|
|
Ellis JI, Fraser G, Russell J (2012). Discharged drilling waste from oil and gas platforms and its effects on benthic communities. Marine Ecology Progress Series 456:285-302. |
|
|
Erftemeijer PLA, Riegl B, Hoeksema BW, Todd PA (2012). Environmental impacts of dredging and other sediment disturbances on corals: A review. Marine Pollution Bulletin 64(9):1737-1765. |
|
|
Freiwald A, Fosså JH, Grehan A, Koslow T, Roberts JM (2004). Cold-water coral reefs: out of sights - no longer out of mind. Biodiversity Series 22. Cambridge: UNEP-WCMC. |
|
|
Gallo A, Rocha AC, Scofano AM, Grossmann G (2006). Computational Modeling of Drill Cuttings and Mud Released in the Sea from E&P Activities in Brazil. In 9th International Marine Environmental Modeling Seminar (IMEMS 2006), Rio de Janeiro, RJ. |
|
|
Gori A, Ferrier-Pagès C, Hennige SJ, Murray F, Rottier C, Wicks LC, Murray R (2016). Physiological response of the cold-water coral Desmophyllum dianthus to thermal stress and ocean acidification. Peer Journal 4:e1606. |
|
|
Hennige SJ, Wicks LC, Kamenos, NA, Perna G, Findlay HS, Roberts JM (2015). Hidden impacts of ocean acidification to live and dead coral framework. Proceedings of the Royal Society B: Biological Sciences 282:20150990. |
|
|
Instituto Brasileiro do Meio Ambiente (IBAMA) (2019). Nota Técnica nº 2/2019/COEXP/CGMAC/DILIC |
|
|
International Association of Oil & Gas Producers (IOGP) (2016). Environmental fate and effects of ocean discharge of drill cuttings and associated drilling fluids from offshore oil and gas operations. Technical Report n. 543. IOGP, London, UK 145 p. |
|
|
Järnegren J, Brooke S, Jensen H (2017). Effects of drill cuttings on larvae of the cold-water coral Lophelia pertusa. Deep Sea Res. II 137:454-462. |
|
|
Kitahara MV (2007). Species richness and distribution of azooxanthellate scleractinia in Brazil. Bulletin of Marine Science 81(3):497-518. |
|
|
Kitahara MV, Capítoli RR, Horn Filho NO (2009). Distribuição das espécies de corais azooxantelados na plataforma e talude continental superior do sul do Brasil. Série Zoologia 99(3):223-236. |
|
|
Konietschke F, Placzek M, Schaarschmidt F, Hothorn LA (2015). nparcomp: an R software package for nonparametric multiple comparisons and simultaneous confidence intervals. Journal of Statistics Software 64(9):1-17. |
|
|
Larsson AI, Purser A (2011). Sedimentation on the cold-water coral Lophelia pertusa: Cleaning efficiency from natural sediments and drill cuttings. Marine Pollution Bulletin 62(6):1159-1168. |
|
|
Larsson AI, Van Oevelen D, Purser A, Thomsen L (2013). Tolerance to long-term exposure of suspended benthic sediments and drill cuttings in the cold-water coral Lophelia pertusa. Marine Pollution Bulletin 70(1-2):176-188 |
|
|
Le Goff-Vitry MC, Rogers AD, Baglow D (2004). A deep-sea slant on the molecular phylogeny of the Sceractinia. Molecular Phylogenetics and Evolution 30:167-177. |
|
|
Lepland A, Buhl-Mortensen P (2008). Barite and barium in sediments and coral skeletons around the hydrocarbon exploration drilling site in the Træna Deep, Norwegian Sea. Environmental Geology 56(1):119-129. |
|
|
Lunden JJ, McNicholl CG, Sears CR, Morrison CL and Cordes EE (2014a). Acute survivorship of the deep-sea coral Lophelia pertusa from the Gulf of Mexico under acidification, warming, and deoxygenation. Marine Science 2(78):1-12. |
|
|
Lunden JJ, Turner JM, McNicholl CG, Glynn CK, Cordes EE (2014b). Design, development, and implementation of recirculating aquaria for maintenance and experimentation of deep-sea corals and associated fauna. Limnology and Oceanography: Methods 12:363-372. |
|
|
Neff JM (2008). Estimation of bioavailability of metals from drilling mud barite. Integrated Environmental Assessment and Management 4(2):184-193. |
|
|
Olariaga A, Gori A, Orejas C, Gili JM (2009). Development of an autonomous Aquarium system for maintaining deep corals. Oceanography 22:44-45. |
|
|
Orejas C, Gori A, Rad-Menéndez C, Last KS, Davies AJ, Beveridge CM, Sadd D, Kiriakoulakis K, Witte U, Roberts JM (2016). The effect of flow speed and food size on the capture efficiency and feeding behaviour of the cold-water coral Lophelia pertusa. Journal of Experimental Marine Biology and Ecology 481:34-40. |
|
|
Orejas C, Taviani M, Ambroso S, Andreou V, Bilan M, Bo M, Brooke S, Buhl-Mortensen P, Cordes E, Dominguez-Carrió C, Ferrier-Pagès C, Godinho A, Gori A, Grinyó J, Gutiérrez-Zárate C, Hennige S, Jiménez |
|
|
CE, Larsson AI, Lartaud F, Lunden J, Maier C, Maier SR, Movilla JI, Murray F, Peru E, Purser A, Rakka M, Reynaud S, Murray RJ, Siles P, Strömberg S, Thomsen L, Van Oevelen D, Veiga A, Carreiro-Silva M (2019). Cold-Water Coral in Aquaria: Advances and Challenges. A Focus on the Mediterranean. In: Coral Reefs of the World, eds. C. Orejas, and C. Jiménez (Switzerland: Springer). |
|
|
Pires DO, Castro CB, Medeiros MS, Thiago CM (2004). Classe Anthozoa. In: Biodiversidade bentônica da região Sudeste-Sul do Brasil: plataforma externa e talude superior, eds. A.C.Z. Amaral and C.L.B. Rossi-Wongtschowski (São Paulo: Universidade de São Paulo). Série Documentos Revizee, Score Sul pp. 71-73 |
|
|
Pires DO (2007). The azooxanthellate coral fauna of Brazil. Bulletin of Marine Science 81(3):265-272. |
|
|
Pires DO, Seabra NA, Silva JVC (2015). Recifes de coral de profundidade: corais construtores e sua distribuição no Brasil. Brazilian Journal of Aquatic Science and Technology 19(3):51-61 |
|
|
Provan F, Nilsen MM, Larssen E, Uleberg KE, Sydnes MO, Lyng E, Øysæd KB, Baussant T (2016). An evaluation of coral Lophelia pertusa mucus as an analytical matrix for environmental monitoring: a preliminary proteomic study. Journal of Toxicology and Environmental Health, Part A 79(13-15):647-657. |
|
|
Purser A (2015). A time series study of Lophelia pertusa and reef megafauna responses to drill cuttings exposure on the Norwegian margin. PlosOne 10(7):e0134076. |
|
|
Purser A, Thomsen L (2012). Monitoring strategies for drill cutting discharge in the vicinity of cold-water coral ecosystems. Marine Pollution Bulletin 64:2309-2316. |
|
|
Rasband WS (1997-2015) ImageJ. National Institutes of Health, Bethesda, Maryland, USA. |
|
|
Roberts JM (2002). The occurrence of the coral Lophelia pertusa and other conspicuous epifauna around an oil platform in the North Sea. Journal of the Society for Underwater Technology 25(2):83-91. |
|
|
Rogers CS (1990). Responses of coral reefs and reefs organisms to sedimentation. Marine Ecology Progress Series. Oldendorf 62(1):185-202. |
|
|
Rye H, Dietlevsen MK (2009). Spreading and deposition of discharges to sea from exploration drilling at the Trolla field in the Norwegian sea. In Abstract Book, International Conference on Health, Safety and Environment in Oil and Gas Exploration and Production, SINTEF Report F11754 |
|
|
Rye H, Reed M, Durgut I, Arzlanoglu Y, Brørs B, Ditlevsen MK, Smit MG, Frost TK (2011). Environmental risk assessment of discharges from onshore facilities to coastal environments. In Abstract Book, International Conference on Health, Safety and Environment in Oil and Gas Exploration and Production, SPE 140430 |
|
|
Smit MGD, Holthaus KIE, Trannum H, Neff, JM, Kjeilen-Eilertsen G, Jak RG, Singsaas I, Huijbregts MAJ, Hendriks J (2008). Species sensitivity distributions for suspended clays, sediment burial, and grain size change in the marine environment. Environmental Toxicology and Chemistry 27(4):1006-1012. |
|
|
Sumida PYG, Yoshinaga MY, Madureira LAS-P, Hovland M (2004). Seabed pockmarks associated with deep-water corals off SE Brazilian continental slope, Santos Basin. Marine Geology 207:159-167. |
|
|
Viana AR, Faugères JC, Kowsmann RO, Lima JAM, Caddah LFG, Rizzo JG (1998). Hydrology, morphology and sedimentology of the Campos continental margin, offshore Brazil. Sedimentary Geology 115:133-157. |
|
|
Weber M (2009). How sediment damages corals (Ph.D. thesis). University of Bremen, Bremen, Germany. |
|
|
Weber M, Lott C, Fabricius KE (2006). Sedimentation stress in a scleractinian coral exposed to terrestrial and marine sediments with contrasting physical, organic and geochemical properties. Journal of Experimental Marine Biology and Ecology 336(1):18-32. |
|
|
Weber M, de Beer D, Lott C, Polerecky L, Kohls K, Abed RMM, Ferdelman TG, Fabricius KE (2012). Mechanisms of damage to corals exposed to sedimentation. Proceedings of the National Academy of Sciences 109(24):E1558-E1567. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


