Full Length Research Paper

ABSTRACT
Fasciolosis is a major problem to livestock production and is considered an emerging health hazard to humans and other animals. An increase in the prevalence of anthelmintic-resistant helminth strains, drug residues in animal products poses a challenge in the control of the disease necessitating research for alternatives. Here, we investigated the presence of anthelmintic phytochemicals in the aqueous and ethanolic extracts of Leonotis nepetifolia dry leaves and their activity on Fasciola gigantica flukes. The flukes were isolated from the infected liver collected from the abattoir maintained in normal saline solution at 37°C. Cold maceration and infusion methods were used for ethanolic and aqueous extractions, respectively. Phytochemical screening and quantification of target phytochemicals were done following standard methods. The percentage mortality caused by each treatment at varying concentrations (2, 6 and 10 mg/ml) was determined and observations were made at three 2 h intervals, and compared with negative control. The lethal concentrations and lethal time for 50% (LC50 and LT50) and 90% (LC90 and LT90) of the flukes were estimated for each treatment. Ethanolic and aqueous extracts of the plant contained anthelmintic phytochemicals. Tannins had a higher concentration in the aqueous extract (0.2362(0.002) mg/ml; p < 0.01) than in the ethanolic extract (0.005(0.001) mg/ml). LC50 and LC90 decreased time of exposure, the aqueous extract showing the lowest LC50 of 0.698 mg/ml and LC90 of 4.435 mg/ml compared to the ethanolic extract with LC50 of 2.521 mg/ml and LC90 of 10.264 mg/ml after 6 h of exposure. LT50 and LT90 decreased with the dosage of the treatments, the aqueous extract had lower LT50 of 2.474 h and LT90 of 5.025 h than that of the ethanolic extract at LT50 of 2.673 h and LT90 of 7.282 h at 10 mg/ml dosage. The anthelmintic activity of L. nepetifolia extracts is concentration and time of exposure dependent. The aqueous extract of L. nepetifolia has higher activity against F. gigantica than the ethanolic extract.
Key words: Anthelmintic, Fasciola gigantica, fascioliasis, Leonotis nepetifolia.
INTRODUCTION
Fasciola species, also known as liver flukes, are among the most important trematode species transmitted mainly by consuming vegetation or water containing metacercariae (Sumbria and Singla, 2020). Specific freshwater snail species are important in the life cycle of the Fasciola flukes. These parasites are an important cause of fascioliasis that infects both livestock and wild animals and has a worldwide distribution (World Health Organization (WHO), 2010). The WHO recognises fascioliasis as an emerging human health threat with 2.4 to 17 million people infected and more than 90 million people at risk of infection (Anuracpreeda et al., 2017). Fascioliasis is also recognised as a human neglected tropical disease (WHO, 2010). It is estimated that worldwide, more than 300 million cattle are exposed to Fasciola spp. causing an estimated loss of over US$3.2 billion per annum in animal productivity (Olaechea, 2004; Joan et al., 2015). Fascioliasis leads to a decrease in weight gain, milk yield and fertility as well as the high condemnation of infected livers at slaughter areas (Malatji et al., 2020).
In East Africa, Fasciola gigantica is the main cause of fascioliasis (Malatji et al., 2020; Walker et al., 2008). However, F. hepatica was reported in cattle from highland areas within the region (Walker et al., 2008). Fasciola hepatica has also been reported to coexist with F. gigantica in Ethiopia and Tanzania (Malatji et al., 2020). In Uganda, reports show the prevalence of Fasciola spp. in highland areas at 78% and lowland areas at 65% (Joan et al., 2015b). The study further reports a prevalence of 73.3% F. gigantica and 5.68% F. hepatica in Kampala abattoir causing an estimated loss of over 231 billion Uganda shillings (92.5 US Dollars) annually on the condemned liver at the abattoir.
Control of fascioliasis is largely dependent on chemotherapy with triclabendazole, closantel, clorsulon and albendazole being the drug of choice (Alvarez-Sanchez et al, 2006; Fairweather et al., 2020). The drug triclabendazole has been reported to be the most effective therapeutic agent for Fasciola spp. control on the market, capable of killing all life stages of the liver fluke (Castro-Hermida et al., 2021). Due to this factor, triclabendazole has been heavily used which has led to the increased spread of resistant fluke populations (Kelley et al., 2016; Fairweather et al., 2020). Albendazole and clorsulon have high efficacy against mature flukes though low efficacy by albendazole and moderate efficacy by clorsulon against immature forms (Alvarez-Sanchez et al, 2006; Rehbein and Visser, 1999; Castro-Hermida et al., 2021). Albendazole and rafoxanide-resistant fluke strains have also been reported in fluke populations isolated from cattle in Egypt (Shokier et al., 2013). Though clorsulon-resistant fluke strains have not been reported, a reduction in the efficacy of the drug against triclabendazole-resistant strains has been reported (Alvarez-Sanchez et al, 2006; Coles and Stafford, 2001). It is predicted that if eradication of anthelmintic resistance is difficult for gastrointestinal nematode parasites, for liver flukes it will even be more difficult (Borgsteede et al., 2005; Kaur et al., 2015; Pawar et al., 2019). These synthetic anthelmintic drug residues have also been reported in livestock products. The high cost of purchasing synthetic drugs makes them unaffordable to low-income farmers in developing countries (Waller, 1997). Research into reliable and cost-effective alternative therapeutics that are less toxic to humans is critically needed (Eguale et al., 2007). There is growing interest in medicinal plants as safer and sustainable alternative sources of anthelmintic agents (Tariq et al., 2009). Compared to synthetic medicines, plant herbs are often considered to be harmless and served as a rich source of numerous novel biologically active compounds (Nasma et al., 2018; Goel et al., 2020 a, b).
The use of herbal remedies has been highly adopted by African countries with about 80% of the population relying on them for their primary health care (Boadu and Asase, 2017; Simoben et al., 2018). The WHO reported that about 60% of Ugandans rely on the use of herbal medicine (WHO 2013). Leonotis nepetifolia (lion’s ear) is a wild herbaceous plant belonging to the mint family (Family Lamiaceae) (Imran et al., 2012). An ethno-botanical survey carried out in Rukungiri district, Southwestern Uganda revealed L. nepetifolia among the most commonly used herbal plant in the treatment of common diseases (Gumisiriza et al., 2021). L. nepetifolia aqueous and ethanolic anthelmintic activity was reported against Pheretima posthuman (Indian earthworm) (Vasuki et al., 2017). While P. posthuman is an annelid and not a helminth sensu stricto, the bioactivity of an extract against these organisms gives preliminary information about the possibility of their anthelmintic activity. L. nepetifolia was also been reported to reduce gastrointestinal egg counts in faecal samples of goats by 78% in combination with Vernonia amygdalina (Mbanzamihigo et al., 2013). L. nepetifolia is a promising anthelmintic alternative to the synthetic anthelmintic therapeutics on market. In this paper, we evaluate the in vitro anthelmintic activity of L. nepetifolia against F. gigantica.
This study investigated the presence of the anthelmintic phytochemicals in the aqueous and ethanolic extracts of L. nepetifolia dry leaves and their activity on F. gigantica, and estimated the lethal dosage for killing 50 and 90% of the helminths by plant extracts.
MATERIALS AND METHODS
Collection of plant materials
L. nepetifolia were collected from the riverbanks of Rwaminyuri stream at grid reference N00°54.595’E031°46.434’ (GPS Essentials Version 4.4.64) in Kiboga District, Central Uganda. The plant was identified using morphological characteristics (Giang et al., 2021). The plant leaves were collected from 50 L. nepetifolia plants by cutting the leaf stalks using a knife in the evening and samples were collected from only the plants at the flowering stage due to the highest composition of secondary metabolites (Ahmad et al., 2006).
Preparation of aqueous extracts
The leaves of L. nepetifolia were dried under shade and crushed in a mortar to form a coarse powder. L. nepetifolia aqueous extract was prepared by infusion method where 500 g of the coarse powder of L. nepetifolia leaves were macerated in 5 L of hot deionized water for 15 min (Balamurugan et al., 2019). The solution was first filtered using a muslin cloth and the filtrate was filtered again using Whatman number one filter paper. The filtrate was concentrated in a rotary evaporator under a vacuum at 55°C for 6 h and lyophilized overnight to dryness using a freeze drier under a vacuum.
Preparation of ethanolic extracts
The extraction was done using cold maceration where 250 g of L. nepetifolia leaf powder was mixed with 2 L of 70% ethanol for 32 h (Balamurugan et al., 2019). The solution was agitated frequently every after 8 h, to increase the extraction yield (Azmir et al., 2013). Excess ethanol was evaporated off using the counter-current method of drying. Only 2 g of the resultant sample extract was dissolved in 10 ml of distilled water to make solutions for phytochemical screening.
Each extract was weighed (dry mass – DM) on a digital balance. The yield extract (EY), was calculated in percentage using the following formula (dos Santos Nascimento et al., 2018):
EY (%) = DM/PM × 100
where PM represents the plant material used for extraction.
Phytochemical screening and quantification of the major anthelminthic phytochemicals
The aqueous and ethanolic extracts were qualitatively phytochemically analyzed using standard methods described by Balamurugan et al. (2019) for the presence of alkaloids (Dragendorff’s test), flavonoids (10% Lead acetate test), phenols (Gelatin test), quinones (Alcoholic KOH test), saponins (Foam test), steroids (chloroform and conc. Sulphiric acid), tannin (ferric chloride test), glycosides (Glacial acetic acid, ferric chloride and conc. Sulphiric acid), and polyphenols (Lead acetate test). Quantification of phytochemicals was done using standard methods.
The total tannin content was determined following the Folin-Denis standard method using a UV spectrophotometer at a wavelength of 726 nm (Katoch, 2011). 5 ml of Folin-Denis reagent and 10 ml of a sodium carbonate solution were added to 100 ml volumetric flasks containing 1 ml test solutions of 1 mg/ml concentration. The resultant solutions were filled up to the mark with distilled water and shaken then left to settle for 30 min. The absorbances were read at a wavelength of 700 nm using a Jenway UV-VIS spectrophotometer. The calibration curve was prepared using absorbances of varying dilutions of 0.4, 0.6, 0.8, 1.0 and 1.2 ml standard tannic acid stock solution. The results were expressed in mg of tannic acid equivalent per g.
The total flavonoid content was determined spectro-photometrically at a wavelength of 430 nm employing the aluminium chloride method with little modification as described by Quettier-Deleu et al. (2000). To 1 ml of the extract solution, 1 ml of 2% methanolic aluminium chloride, 6 H2O solutions was added. The solutions were left to settle for 10 min. The absorbances of the resultant solutions were determined using a Jenway UV-VIS spectrophotometer at 430 nm. The calibration curve was prepared using absorbances of varying dilutions of 0.4, 0.6, 0.8, 1.0 and 1.2 ml standard rutin solution. The results were expressed in mg rutin per g dry matter.
The total Saponin content was determined spectro-photometrically using a vanillin-sulphuric acid assay at wavelength 543 nm (Hiai and Oura, 1976). 0.5 ml of 8% (W/V) vanillin solution was added to 1 ml extract solution of 1 mg/ml concentration. 5 ml of 72% (v/v) sulphuric acid was added and the resultant solution was homogenized. The mixture was incubated at 60°C in a water bath followed by cooling in an ice-cold bath for 5 min. The absorbances of the solutions were determined using a Jenway UV-VIS spectrophotometer at 550 nm. The calibration curve was prepared using absorbances of varying dilutions of 0.4, 0.6, 0.8, 1.0 and 1.2 ml Diosgenin. The total saponin content was measured in mg Diosgenin equivalent per gram of extract. All the phytochemicals quantified were measured in triplicates. Only tannin, flavonoids and saponin in the plant extracts were quantified because these phytochemicals have been reported by several studies as being common and in high concentrations, in plant extracts with anthelminthic activity hence they are the major anthelminthic agents (Ndjonka et al., 2011, 2014; Samje et al., 2014; Santos et al., 2019).
Collection of F. gigantica
F. gigantica flukes were collected from the infected livers of cattle at the Mbarara City abattoir in Ruti, western Uganda (S00°37.253’E030°39.589’ (GPS Essentials Version 4.4.64)). The specimen were identified by examining the morphological appearance and measuring the body dimensions (Joan et al., 2015a). The flukes were put in flasks containing warm (30-35°C) normal saline – 5% glucose solution and transported to the laboratory where they were maintained in an incubator at 37°C in normal saline solution (Goswami et al., 2011).
Evaluation of anthelmintic activity
A total of 210 F. gigantica of relatively the same size were selected randomly and considered for the whole study. Treatment solutions were prepared by dissolving 0.18, 0.54, and 0.90 g of the extracts in 90 ml of normal saline solution in separate beakers to make treatment solutions of 2, 6 and 10 mg/ml concentrations, respectively. Concentrations were adopted from Ibekwe (2019). In the treatment groups, ten flukes were placed in 30 ml of each of the following treatments in beakers. This was performed for both the ethanolic and the aqueous extract in a parallel arrangement. All these treatments were made in triplicates. Flukes in the control group were dipped in normal saline without any treatment solution. All the beakers were then placed in an incubator at 37°C. Observations were made in time intervals of 0, 2, 4 and 6 h. Death of the fluke was determined by observing their movement. If the fluke did not show any movement when touched by anatomy tweezers, it was dipped in slightly warm water (50°C), and if no movement was observed still, the fluke was considered dead (Dash et al., 2017).
Data analysis
One-way ANOVA test was used to determine the significant difference between the anthelmintic activities caused by different concentrations of the test solutions and the control for normally distributed data while for the non-normally distributed data Kruskal Wallis test was employed. A post hoc Fisher’s LSD test was performed for the pairwise comparison to determine the significant difference among treatments. Independent samples t-test (two-tailed test) was used to compare the mean percentage mortality caused by the ethanolic and aqueous extracts of L. nepetifolia. An independent sample t-test was also employed to compare the mean concentrations of the major anthelminthic phytochemicals in the ethanolic and aqueous plant extracts. The effect of the concentration of each treatment on the mean % mortality of F. gigantica was determined using regression analysis. A two-way ANOVA test was carried out to determine the combined effect of time of exposure and concentration of the test solution on the mean percentage mortality of the flukes. All the analysis was performed at 0.05 level of significance.
The LC50 (lethal concentration that causes 50% death) and LC90 (lethal concentration that causes 90% death) for each treatment solution were determined using the probit regression model at 95 level of confidence in SPSS Version 20 (Ibekwe, 2019). The LT50 (lethal time required for a given treatment to cause 50% death) and LT90 (lethal time required for a given treatment to cause 90% death) were determined using the probit regression model. The probit model equation Probit (P) = βlog10(θ) + γ, was employed where β represents the gradient, θ represents time for LT / concentration for LC, and γ represents y-intercept.
RESULTS
Anthelminthic phytochemicals in the aqueous and ethanolic extract of L. nepetifolia leaves
The percentage yield for L. nepetifolia aqueous extract was 20.25% while for its ethanolic extract it was 21.29%. Qualitative tests done on leaf extracts of L. nepetifolia revealed that the ethanolic extract had plenty of flavonoids, saponins and tannins though alkaloids were moderate. Glycosides were present only in traces. The aqueous extract had plenty of flavonoids, alkaloids, and tannins, moderate saponins and was devoid of glycosides (Table 1).
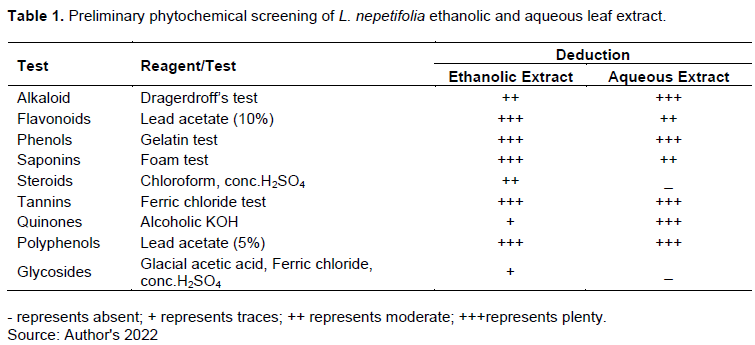
L. nepetifolia’s aqueous extract had a significantly higher concentration of tannin than in its ethanolic extract (t =204.137; p <0.001; df =4); however, the ethanolic extract has significantly higher concentrations of flavonoids and saponin (P<0.001; df=4) (Figure 1).
Efficacy of ethanolic and aqueous leaf extracts of L. nepetifolia in killing F. gigantica
There was no death in the control group (0.00 mg/ml). The mean percentage mortality caused by all treatments was higher than in the control (p<0.001) except for 2 mg/ml aqueous extract of L. nepetifolia.
The mean percentage mortality of the flukes increased with an increase in the concentration of the treatments for both the ethanolic (r2= 0.968, p < 0.001) and the aqueous (r2=0.907, P<0.001) leaf extract of L. nepetifolia. Mann Whitney-U test revealed no significant difference between the anthelminthic activity of the aqueous and ethanolic extracts (p = 0.253, N=12), though the aqueous extract (Mean Rank = 14.13) showed slightly higher activity compared to the ethanolic extract (Mean Rank=10.88) after 4 h of exposure. The anthelmintic activity of the aqueous extract was significantly higher than that of the ethanolic extract only at 6 mg/ml (p = 0.043).
For the ethanolic leaf extract of L. nepetifolia, there existed a significant difference in its anthelmintic activity between all the concentrations (p<0.001, N=12); hence, showing a dose-dependent manner with 10 mg/ml being the most effective. For the aqueous leaf extract of L. nepetifolia, there existed no significant difference between 6 and 10 mg/ml and hence 6 mg/ml was the most effective concentration.
The interaction of concentration and time shows a strong effect in killing F. gigantica (F(6)=16.697, p<0.001, Eta squared (?p2)=0.807). The highest mortality was caused by 10 mg/ml concentration of the treatments after 6 h of exposure (Mean=86.667(3.191)).
LC50, LC90 and LT50, LT90
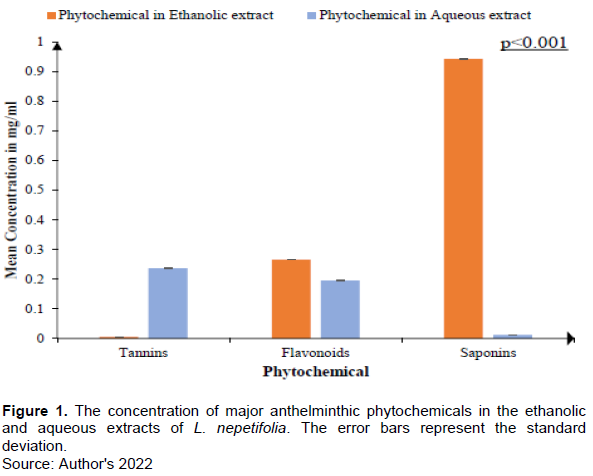
The lower the LC50 and LC90, the more lethality of the plant extracts to the parasite. Generally, the concentration of different extracts required to kill 50% (LC50) and 90% (LC90) decreases with exposure time. This indicates that the anthelmintic activity of L. nepetifolia aqueous and ethanolic leaf extracts area time of exposure dependent. The aqueous extract of L. nepetifolia leaves shows the lowest LC50 of 0.698 mg/ml and LC90 of 4.435 mg/ml in compulsion with that shown by the ethanolic extract shown in Table 2.
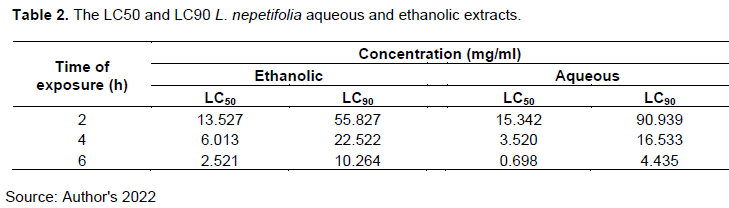
The lower the LT50 and LT90, the more lethality of the plant extracts to the flukes. The time required for different plant extracts to cause 50% (LT50) and 90% (LT90) fluke death decreases with an increase in the dosage of the treatments. This indicates that the anthelmintic activity of L. nepetifolia aqueous and ethanolic leaf extracts is dose-dependent. The aqueous extract required the shortest time to cause 50 and 90% fluke death for all the treatment dosages considered in this study as shown in Table 3; this confirms that the aqueous leaf extract of L. nepetifolia has higher anthelminthic efficacy against F. gigantica than the ethanolic extract.
DISCUSSION
Phytochemical screening of L. nepetifolia ethanolic and aqueous extracts
Plant polyphenols and phenolics (secondary metabolites) are widely distributed in the plant kingdom, surprisingly, sometimes in high concentrations (Harbonne, 1993; Khan et al., 2019). The present study reports a rich composition of phytochemicals including polyphenols and phenol in the ethanolic and aqueous extracts of L. nepetifolia dry leaves. Polyphenolic and phenolic compounds show anthelmintic activity (Bate-Smith, 1962; Soldera-Silva et al., 2018). For example flavonoids, mainly isoflavones inhibit glycolysis enzymes and disturb calcium homeostasis and nitrous oxide activity hence leading to the death of the parasite (Stepek et al., 2006). Some synthetic phenolic anthelmintics for example niclosamide, oxyclozanide and bithionol are shown to interfere with energy generation in helminth parasites by uncoupling oxidative phosphorylation (Martin, 1997). It is possible that natural phenolic compounds in the extracts of L. nepetifolia leaves act in the same way hence the evidence of their anthelminthic activity. Tannins have been reported to interfere with cell membrane glycoprotein and interfering with energy generation by uncoupling oxidative phosphorylation (Roy et al., 2010). Similarly, it has been reported that alkaloids may act on the central nervous system and cause paralysis in worms (Roy et al., 2010). Several studies have reported steroids and cardiac glycosides to possess anthelmintic potency (Chang and Flores, 2015; Mahardika et al., 2017).
The presence of different phytochemicals in the ethanolic and aqueous extracts of L. nepetifolia dry leaves varies greatly. The differences in the presence of different phytochemicals in the extracts is due to difference in the extraction abilities of the solvents used (Bwalya, 2015). Various plant secondary metabolites are soluble in specific extractants depending on the solvent polarity (Cabardo and Portugaliza, 2017). This also explains a reason why different extracts of the same plants have slightly different phytochemicals and varying quantities.
Anthelmintic activity of ethanolic and aqueous extracts of L. nepetifolia dry leaves
The mean percentage mortality caused by all treatments was significantly higher than that caused by the control (p<0.001). This may be attributed to the presence of anthelminthic phytochemicals in both the aqueous and ethanolic extracts of L. nepetifolia leaves which is absent in the control group. However, 2 mg/ml of aqueous leaf extract of L. nepetifolia showed no significant difference in its anthelminthic activity with that of the control, this may be attributed to the presence of anthelminthic phytochemicals in low quantities below the lethal dosage that is required to cause significant death of flukes.
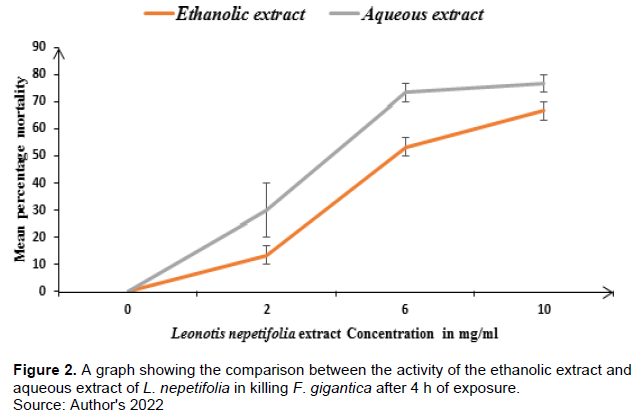
The anthelmintic activity of the aqueous extract of L. nepetifolia dry leaves was significantly higher than that of its ethanolic extract at 6 mg/ml though there was no significant difference between the anthelmintic activity of the ethanolic and aqueous extract of L. nepetifolia dry leaves at 2 and 10 mg/ml. The aqueous extract showed greater activity than the ethanolic extract (Figure 2) because it is richer in phenolic compounds which have anthelmintic properties; for example, it has plenty of alkaloids which are moderate in the ethanolic extract (Table 1). Alkaloids have demonstrated anthelminthic potency against blood flukes (Schistosoma mansoni) (Wangchuk et al., 2016; Perrett and Whitfield, 1995). Alkaloids act on the central nervous system of the parasite causing paralysis and eventually death of the worms (Manke et al., 2015), this is due to the fact that alkaloid salts compete with acetylcholine for the same binding site preventing acetylcholine from binding (Tarnopolsky and Beal, 2001). This greater anthelminthic activity of the aqueous extract may also be attributed to the presence of significantly higher tannin content in the aqueous extract than in the ethanolic extract (p < 0.001, N=3). Tannins have been reported to have an anthelminthic effect against helminths such as Ascari ssuum (Williams et al., 2014). Tannins are found of forming tannin-protein complexes with helminthic parasites that interfere with physiological processes necessary for the survival of these parasites for example tannin-nematode protein complexes have been reported to alter feeding, motility, fecundity, and a range of enzyme-mediated biochemical functions in nematodes (Hoste and Quijada, 2016; Hansen et al., 2016), Tannins such as proanthocyanidins cause direct physical damage on the external and internal parasite structures; hence, leading to death (Hoste and Karla, 2011). Proanthocyanidins have also demonstrated the ability to inhibit glutathione-S-transferases enzymes which are essential for parasite detoxification and also contribute to parasitic mortality (Hansen et al., 2016). This maybe the same way it affects Fasciola gigantica in the present study. Quinones and their derivatives are reported to have anthelmintic activity against Taxocara canis larvae in both in vivo and in vitro evaluation (Mata-Santos et al., 2016). From the findings of this study, the aqueous dry leaf extract of L. nepetifolia was found to contain plenty of quinones which were in traces in its ethanolic extract; this probably may also be the reason why the aqueous extract has a greater anthelmintic activity than the ethanolic extract.
On the other hand, saponins are also reported to have anthelminthic activity due to their ability to form complexes with the parasitic cell membrane components hence forming membrane pores that alter the cell permeability of the parasites hence contributing to their death (Santos et al., 2018). In this study, saponins were found significantly higher in the ethanolic extract than in the aqueous extract, though in contrast, the aqueous extract showed greater activity than the ethanolic extract. This may be attributed to a stronger anthelminthic efficacy of tannins to saponins as reported by a review on the anthelminthic activity of plants against gastrointestinal nematodes of goats by Santos et al. (2019).
The plant secondary metabolites can act individually however their combined interactions have been reported to be more potent (Kaufman et al., 1998). The co7mbinations of these metabolites may offer additive, synergistic or antagonistic anthelmintic activity at a single or multiple target sites (Wynn and Fougère, 2006). Combinations of condensed tannins and flavonoids are found of showing an anthelminthic synergistic effect (Klongsiriwet et al., 2015). In this study, tannins and flavonoids were found to be in comparatively equal amounts in the aqueous extract (Figure 1) hence synergism which may also be contributing to a higher anthelminthic efficacy of the aqueous extract.
Generally, the concentration of different extracts required to kill 50% (LC50) and 90% (LC90) decreases with the time of exposure (Table 2). This indicates that the efficacy of the plant extracts in killing F. gigantica is dependent on the time of exposure. This may be due to the uptake of the bioactive phytochemicals from the extract which increases in the worms’ body with the time of exposure hence an increase in the anthelminthic activity.
The time required for different plant extracts to cause 50% (LT50) and 90% (LT90) fluke death decreases with an increase in the dosage of the treatments (Table 3). This indicates that the anthelminthic efficacy of the plant extracts works in a dose-dependent manner, a similar observation was made by Vasuki et al. (2017) when treating Pheretima posthuma using the aqueous and ethanolic whole plant extract, this may be attributed to the fact that an increase in the concentration of the extract increases the amounts of the bioactive substances to which the flukes are exposed hence increasing worm mortality.
CONCLUSION
Both ethanolic and aqueous extracts of L. nepetifolia contain anthelmintic phytochemicals. The aqueous extract of L. nepetifolia dry leaves is more effective in killing F. gigantica compared to its ethanolic extract with 6 mg/ml concentration being its most effective concentration while 10 mg/ml concentration is the most effective for its ethanolic extract. The anthelmintic activity of L. nepetifolia dry leaves is concentration and time of exposure dependent. Further studies are required to examine its in vivo activity and the mode of action of its phytochemicals against F. gigantica at a molecular level. Also, further studies are required to isolate and elucidate the structure of the anthelminthic bioactive molecules in L. nepetifolia
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Ahmad I, Aqil F, Owais M (2006). Medicinal Plant Biotechnology Medicinal Chemistry of Bioactive Natural Products. Provide Valid online source/link |
|
|
Alvarez-Sanchez MA, Mainar-Jaime RC, Perez-Garcia J, Rojo-Vázquez FA (2006). Short Communications Resistance of Fasciola hepatica to triclabendazole and albendazole in sheep in Spain. Veterinary Record 159(13):424. |
|
|
Anuracpreeda P, Chawengkirttikul R, Ngamniyom A, Panyarachun B, Puttarak P, Koedrith P, Intaratat N (2017). The in vitro anthelmintic activity of the ethanol leaf extracts of Terminalia catappa L. on Fasciola gigantica. Parasitology 144(14):1931. |
|
|
Azmir J, Zaidul ISM, Rahman MM, Sharif KM, Mohamed A, Sahena F, Jahurul MHA, Ghafoor K, Norulaini NAN, Omar AKM (2013). Techniques for extraction of bioactive compounds from plant materials: A review. Journal of Food Engineering 117(4):426-436. |
|
|
Balamurugan V, Fatima S, Velurajan S (2019). a Guide To Phytochemical Analysis. International Journal of Advance Research and Innovative Ideas In Education 5(1):236-245. |
|
|
Bate-Smith EC (1962). The phenolic constituents of plants and their taxonomic significance. I. Dicotyledons. Botanical Journal of the Linnean Society 58(371):95-173. |
|
|
Boadu AA, Asase A (2017). Documentation of herbal medicines used for the treatment and management of human diseases by some communities in southern Ghana. Evidence-Based Complementary and Alternative Medicine, 2017. |
|
|
Borgsteede FHM, MollL, Vellema P (2005). Lack of reversion in triclabendazole resistant Fasciola hepatica. Veterinary Record 156(11):350-351. |
|
|
Bwalya AG (2015). Evaluation of the in vitro biological activities and phytochemical profiling of eight ficus species collected in Zambia. UCL (University College London). |
|
|
Cabardo Jr DE, Portugaliza HP (2017). Anthelmintic activity of Moringa oleifera seed aqueous and ethanolic extracts against Haemonchus contortus eggs and third stage larvae. International Journal of Veterinary Science and Medicine 5(1):30-34. |
|
|
Castro-Hermida JA, González-WarletaM, Martínez-Sernández V, Ubeira FM, Mezo M (2021). Current challenges for fasciolicide treatment in ruminant livestock. Trends in Parasitology 37(5):430-444. |
|
|
Chang ACG, Flores MJC (2015). Morphology and viability of adult Fasciola gigantica (giant liver flukes) from Philippine carabaos (Bubalus bubalis) upon in vitro exposure to lead. Asian Pacific Journal of Tropical Biomedicine 5(6):493-496. |
|
|
Coles GC, Stafford KA (2001). Activity of oxyclozanide, nitroxynil, clorsulon and albendazole against adult triclabendazole-resistant Fasciola hepatica. The Veterinary Record 148(23):723-724. |
|
|
Dash PR, Mou KM, Erina IN, Ripa FA, Al Masud KN, Ali MS (2017). Study of anthelmintic and insecticidal activities of different extracts of Kaempferia galanga. International Journal of Pharmaceutical Sciences and Research 8(2):729. |
|
|
dos Santos Nascimento LB, de Aguiar PF, Leal?Costa MV, Coutinho MAS, Borsodi MPG, Rossi?Bergmann B, Tavares ES, Costa SS (2018). Optimization of aqueous extraction from Kalanchoe pinnata leaves to obtain the highest content of an anti?inflammatory flavonoid using a response surface model. Phytochemical Analysis 29(3):308-315. |
|
|
Eguale T, Tilahun G, Debella A, Feleke A, Makonnen E (2007). In vitro and in vivo anthelmintic activity of crude extracts of Coriandrum sativum against Haemonchus contortus. Journal of Ethnopharmacology 110(3):428-433. |
|
|
Fairweather I, Brennan GP, Hanna REB, Robinson MW, Skuce PJ (2020). Drug resistance in liver flukes. International Journal for Parasitology: Drugs and Drug Resistance 12:39-59. |
|
|
Giang NTH, Van Ngot P, Thanh DTN (2021). Morphological, anatomical and antibacterial characteristics of Leonotis nepetifolia plants growing in Binh Thuan Province, Vietnam. GSC Biological and Pharmaceutical Sciences 14(2):53-63. |
|
|
Goel V, Kaur P, Singla LD and Choudhury D (2020a). Biomedical evaluation of Lansium parasiticum extract protected silver nanoparticles against Haemonchus contortus, a parasitic worm. Frontiers in Molecular Biosciences 7:595646. |
|
|
Goel V, Singla LD and Choudhury D (2020b).Cuminaldehyde induces oxidative stress-mediated physical damage and death of Haemonchus contortus. Biomedicine and Pharmacotherapy 130:110411. |
|
|
Goswami S, Pandey A, Tripathi P, Singh A, Rai A (2011). An in vitro evaluation of the anthelmintic activity of Hedychium spichatum rhizomes and Zingiber zerumbet rhizomes on the Pheritima Posthuma model: A comparative study. Pharmacognosy Research 3(2):140. |
|
|
Gumisiriza H, Sesaazi CD, Olet EA, Kembabazi O, Birungi G (2021). Medicinal plants used to treat" African" diseases by the local communities of Bwambara sub-county in Rukungiri District, Western Uganda. Journal of Ethnopharmacology 268:113-578. |
|
|
Hansen TVA, Fryganas C, Acevedo N, Caraballo L, Thamsborg SM, Mueller-Harvey I, Williams AR (2016). Proanthocyanidins inhibit Ascaris suum glutathione-S-transferase activity and increase susceptibility of larvae to levamisole in vitro. Parasitology International 65(4):336-339. |
|
|
Harbonne JB (1993). New naturally occurring plant polyphenols. |
|
|
Hiai S, Oura H (1976). Color Reaction of Some Sapogenins With Vanillin Sulphuric Acid. Planta Medica 29(2):116-122. |
|
|
Hoste H, Karla M (2011). Ultrastructural changes in the third- stage , infective larvae of ruminant nematodes treated with sainfoin. |
|
|
Hoste H, Quijada J (2016). Interactions Between Nutrition and Infections With Haemonchus contortus and Related Gastrointestinal Nematodes in Small Ruminants. In Advances in Parasitology (Vol. 93). Elsevier Ltd. |
|
|
Ibekwe HA (2019). In vitro anthelmintic activities of aqueous crude extract of Azadirachta indica on Paramphistomum cervi and Fasciola hepatica. International Journal of Veterinary Sciences and Animal Husbandry 4(1):14-18. |
|
|
Imran S, Suradkar SS, Koche DK (2012). Phytochemical analysis of leonotis nepetifolia (l.) r. br., a wild medicinal plant of lamiaceae. Bioscience Discovery 3(2):196-197. |
|
|
Joan N, Stephen MJ, Bashir M, Kiguli J, Orikiriza P, Bazira J, Itabangi H, Stanley IJ (2015a). Prevalence and economic impact of bovine fasciolosis at Kampala City abattoir, Central Uganda. |
|
|
Joan N, Stephen MJ, Bashir M, Kiguli J, Orikiriza P, Bazira J, Itabangi H, Stanley IJ (2015b). Prevalence and Economic Impact of Bovine Fasciolosis at Kampala City Abattoir , Central Uganda. 7(November 2014):109-117. |
|
|
Katoch R (2011). Analytical techniques in biochemistry and molecular biology. Springer Science and Business Media |
|
|
Kaufman PB., Cseke LJ, Warber S, Duke JA, Brielmann, HL.(1998). Natural products from plants. CRC Press Inc. |
|
|
Kaur P, Singla LD, Singh E, Pawar P (2015) Anthelmintic resistance menace in small ruminants: Solutions to this emerging issue. Punjab Veterinary Journal (June-December) 81-85. |
|
|
Kelley JM, Elliott TP, Beddoe T, Anderson G, Skuce P, Spithill TW (2016). Current threat of triclabendazole resistance in Fasciola hepatica. Trends in Parasitology 32(6):458-469. |
|
|
Khan AM, Bhadauria S, Yadav R (2019). Phytochemical Screening and Antioxidant activity of extract of different parts of Adhatoda vasica. Research Journal of Pharmacy and Technology, 12(12):5699-5705. |
|
|
Klongsiriwet C, Quijada J, Williams AR, Mueller-Harvey I, Williamson EM, Hoste H (2015). Synergistic inhibition of Haemonchus contortus exsheathment by flavonoid monomers and condensed tannins. International Journal for Parasitology: Drugs and Drug Resistance 5(3):127-134. |
|
|
Mahardika MM, Sudjarwo SA, Koesdarto S (2017). Anthelmintic Activity of Ocimum sanctum Linn. Leaves Ethanol Extract Against Fasciola gigantica in vitro. KnE Life Sciences, 266-277. |
|
|
Malatji MP, Pfukenyi DM, Mukaratirwa S (2020). Fasciola species and their vertebrate and snail intermediate hosts in East and Southern Africa: a review. Journal of Helminthology, p. 94. |
|
|
Manke MB, Dhawale SC, Jamkhande PG (2015). Helminthiasis and medicinal plants: a review. Asian Pacific Journal of Tropical Disease 5(3):175-180. |
|
|
Martin RJ (1997). Modes of action of anthelmintic drugs. The Veterinary Journal 154(1):11-34. |
|
|
Mata-Santos T, Mata-Santos HA, Carneiro PF, De Moura KCG, Fenalti JM, Klafke GB, Cruz LAX, Martins LHR, Pinto NF, Pinto M (2016). Toxocara canis: anthelmintic activity of quinone derivatives in murine toxocarosis. Parasitology 143(4):507-517. |
|
|
Mbanzamihigo L, Dethie F, Kabera NJ, Ugirinshuti V, Nyirabageni A (2013). Evaluation of the effectiveness of two medicinal plants Vernonia amygdalina and Leonotis nepetaefolia on the gastrointestinal parasites of goats in Rwanda: Case study of Huye and Gisagara districts. Journal of Veterinary Medicine and Animal Health 5(8):229-236. |
|
|
Nasma A, Aishath N, Azilah A, Sulaiman AZ (2018). Optimization of vitexin and isovitexin compounds extracted from dried Mas Cotek leaves using one-factor-at-a-time (OFAT) approach in aqueous extraction. International Food Research Journal 25(6):2560-2571. |
|
|
Ndjonka D, Abladam ED, Djafsia B, Ajonina-Ekoti I, Achukwi MD, Liebau E (2014). Anthelmintic activity of phenolic acids from the axlewood tree Anogeissus leiocarpus on the filarial nematode Onchocerca ochengi and drug-resistant strains of the free-living nematode Caenorhabditis elegans. Journal of Helminthology 88(4):481-488. |
|
|
Ndjonka D, Agyare C, Lüersen K, Djafsia B, Achukwi D, Nukenine EN, Hensel A, Liebau E (2011). In vitro activity of Cameroonian and Ghanaian medicinal plants on parasitic (Onchocerca ochengi) and free-living (Caenorhabditis elegans) nematodes. Journal of Helminthology 85(3):304-312. |
|
|
Olaechea F (2004). Red de Helmintología de FAO para América Latina y el Caribe. Conferencia Electrónica. |
|
|
World Health Organization (WHO) (2010). Working to overcome the global impact of neglected tropical diseases: first WHO report on neglected tropical diseases (Issue WHO/HTM/NTD/2010.1). |
|
|
World Health Organization (WHO) (2013). WHO traditional medicine strategy: 2014-2023. |
|
|
Pawar PD, Singla LD, Kaur P Bal MS and Javed M (2019). Evaluation and correlation of multiple anthelmintic resistance to gastrointestinal nematodes using different faecal egg count reduction methods in small ruminants of Punjab, India. Acta Parasitologica 64:456-463. |
|
|
Perrett S, Whitfield PJ (1995). Atanine (3-dimethylallyl-4-methoxy-2- quinolone), an alkaloid with anthelmintic activity from the Chinese medicinal plant, Evodia rutaecarpa. Planta Medica 61(03):276-278. |
|
|
Quettier-Deleu C, Gressier B, Vasseur J, Dine T, Brunet C, Luyckx M, Cazin M, Cazin JC, Bailleul F, Trotin F (2000). Phenolic compounds and antioxidant activities of buckwheat (Fagopyrum esculentum Moench) hulls and flour. Journal of Ethnopharmacology 72(1-2):35-42. |
|
|
Rehbein S, Visser M (1999). Efficacy of an injectable ivermectin/clorsulon combination against Fasciola hepatica in sheep. Veterinary Record 145(16):468. |
|
|
Roy H, Chakraborty A, Bhanja S, Nayak BS, Mishra SR, Ellaiah P (2010). Preliminary phytochemical investigation and anthelmintic activity of Acanthospermum hispidum DC. Journal of Pharmaceutical Science and Technology 2(5):217-221. |
|
|
Samje M, Metuge J, Mbah J, Nguesson B, Cho-Ngwa F (2014). In vitro anti-Onchocerca ochengi activities of extracts and chromatographic fractions of Craterispermum laurinum and Morinda lucida. BMC Complementary and Alternative Medicine 14(1):1-12. |
|
|
Santos ACV, Santos FO, Lima HG, Da Silva GD, Uzêda RS, Dias ÊR, Branco A, Cardoso KV, David JM, Botura MB (2018). In vitro ovicidal and larvicidal activities of some saponins and flavonoids against parasitic nematodes of goats. Parasitology 145(14):1884-1889. |
|
|
Santos FO, Cerqueira APM, Branco A, Batatinha MJM, Botura MB (2019). Anthelmintic activity of plants against gastrointestinal nematodes of goats: A review. Parasitology, 146(10):1233-1246. |
|
|
Shokier KM, Aboelhadid SM, Waleed MA (2013). Efficacy of five anthelmintics against a natural Fasciola species infection in cattle. Beni-Suef University. Journal of Basic and Applied Sciences 2(1):41-45. |
|
|
Simoben CV, Ntie-Kang F, Akone SH, Sippl W (2018). Compounds from African Medicinal Plants with Activities against Selected Parasitic Diseases: Schistosomiasis, Trypanosomiasis and Leishmaniasis. Natural Products and Bioprospecting 8(3):151-169. |
|
|
Soldera-Silva A, Seyfried M, Campestrini LH, Zawadzki-Baggio SF, Minho AP, Molento MB, Maurer JBB (2018). Assessment of anthelmintic activity and bio-guided chemical analysis of Persea americana seed extracts. Veterinary Parasitology 251:34-43. |
|
|
Stepek G, Lowe AE, Buttle DJ, Duce IR, Behnke JM (2006). In vitro and in vivo anthelmintic efficacy of plant cysteine proteinases against the rodent gastrointestinal nematode, Trichuris muris. Parasitology 132(5):681-689. |
|
|
Sumbria D, Singla LD (2020). Fascioliasis in ruminants. Livestock line 13(12):10-11. |
|
|
Tariq KA, Chishti MZ, Ahmad F, Shawl AS (2009). Anthelmintic activity of extracts of Artemisia absinthium against ovine nematodes. Veterinary Parasitology 160(1-2):83-88. |
|
|
Tarnopolsky MA, Beal MF (2001). Potential for creatine and other therapies targeting cellular energy dysfunction in neurological disorders. Annals of Neurology 49(5):561-574. |
|
|
Vasuki K, Murugananthan G, Ragavi P (2017). Phytochemical and Anthelmintic Studies on the Whole Plant of Leonotis Nepetifolia (L). R. BR. |
|
|
Walker SM, Makundi AE, Namuba FV, Kassuku AA, Keyyu J, Hoey EM, Prödohl P, Stothard JR, Trudgett A (2008). The distribution of Fasciola hepatica and Fasciola gigantica within southern Tanzania-constraints associated with the intermediate host. Parasitology 135(4):495-503. |
|
|
Waller PJ (1997). Sustainable helminth control of ruminants in developing countries. Veterinary Parasitology 71(2-3):195-207. |
|
|
Wangchuk P, Giacomin PR, Pearson MS, Smout MJ, Loukas A (2016). Identification of lead chemotherapeutic agents from medicinal plants against blood flukes and whipworms. Scientific Reports 30;6(1):1-0. |
|
|
Williams AR, Fryganas C, Ramsay A, Mueller-harvey I, Thamsborg SM (2014). Direct anthelmintic effects of condensed tannins from diverse plant sources against Ascaris suum. PloS one 9(5):e97053. |
|
|
Wynn SG, Fougère B (2006). Veterinary herbal medicine. Elsevier Health Sciences P 695. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


