Review

ABSTRACT
Small ruminants farming is a traditional activity mostly practiced by local populations in developing countries since several centuries. Nowadays, due to many biotic and climatic factors, it faces various problems which damage smallholders’ income especially those related to gastrointestinal parasites. In opposite to the chemical drugs use in controlling those parasites, medicinal plants have been investigated with fewer side effects on both the meat quality and the environment. This current study aimed at reviewing Haemonchus contortus prevalence in small ruminants across the world and present medicinal plants that have been investigated in the last decades. H. contortus is identified as the most significant nematode parasite in small ruminants due to its high prevalence reported by many studies. Its presence in small ruminants results in a loss of feed absorption and disturbance of nutrient metabolism, which lead to poor performance and significant economic loss in the herds, especially in rural areas of developing countries. For the past decades, its control was mainly based on the use of chemical anthelmintics; whose use has been limited due to several factors like the irrational and misuse. Recently, the use of medicinal plants has been identified as alternatives methods of its control with conclusive results. Parts of plants or the whole plants of several plant species were reported to be relevant to control H. contortus infection in small ruminants such as: Bridelia ferruginea, Mitragyna inermis, Combretum glutinosum, Hagenia abyssinica, Chenopodium ambrosioides, Leucaena leucocephala, Phytolacca icosandra, Eucalyptus staigeriana, Carica papaya, Newbouldia laevis and Zanthoxylum zanthoxyloïdes.
Key words: Economic losses, gastrointestinal nematodes, chemical anthelmintics, medicinal plants, poor performance.
Abbreviation: BW, Body weight; FSA, Faculty of Agronomic Sciences; GIN, gastrointestinal nematode/GINs: gastrointestinal nematodes; LESA, Laboratory of Ethnopharmacology and Animal Health; MP, metabolisable protein; UAC, University of Abomey-Calavi; WAAPP, West Africa Agricultural Productivity Project.INTRODUCTION
Small ruminants are essential in subsistence agriculture owing to their exceptional adaptability in difficult environmental conditions. They provide raw materials for the agro-industries and their manure is used as a source of biogas (Adua and Hassan, 2016) and fertilizer for promoting crop production. Additionally, they perform key sociocultural functions that are hardly quantifiable in monetary terms; for example, their use for rituals and sacrifices, in or during festivities, and as insurance against poor harvests (Hassan et al., 2013) and are also used for teaching and research. Despite all these benefits, the sector receives little attention and faces various challenges, mainly feed and health problems, particularly those related to gastrointestinal nematodes that are very detrimental to livestock (Hounzangbe-Adote et al., 2005). Haemonchus contortus infections are commonly identified as the most significant (Emery et al., 2016; Jamalm et al., 2016) with significant rates of growth and milk yield reduction in small ruminants in tropical environments leading them to production losses in herds, especially grass-fed small ruminants (Andrea et al., 2011). Several studies carried out in small ruminants revealed the existence of polyparasitism with strongles and prevalence rates of digestive strongyles, especially H. contortus (Salifou, 1996; Attindehou et al., 2012; Adua and Hassan, 2016). As a direct consequence, both the carcass yield of these animals and the income of the small farmers are decreasing. In some areas, particularly in the tropics and subtropics where environmental conditions are ideal for the development and transmission of the nematode parasite, frequent use of synthetic anthelmintics has been successful in solving the problem of nematodes (Knox et al., 2006; Torres-Acosta and Hoste, 2008). In parallel with increasing and not always reasoned use of these chemicals, parasites have become increasingly resistant to anthelmintics. In addition, the high cost, limited availability of these chemicals and the drug residues in final products and the environment following their use are other factors that discourage many farmers from using them in some emerging countries (Knox et al., 2006).
Faced all these challenges, it becomes essential to develop new methods to control parasitism. Indeed, improvement of animal nutrition through feed supplements and the use of medicinal plants have been identified as alternatives adapted to the financial means and socio-cultural environment of the populations (Wabo et al., 2012). Recent studies have revealed the anthelmintic effect of several medicinal plants in the control of gastrointestinal nematodes parasites, especially H. contortus in small ruminants that can be considered when designing parasites control programmes. Therefore, this current study aims to present an overview of the prevalence and the effects of H. contortus on small ruminants’ production (growth and milk production), then to summarize studies conducted on some medicinal plants with anthelmintic properties tested against H. contortus.
MORPHOLOGY AND BIOLOGICAL CHARACTERISTICS OF H. CONTORTUS
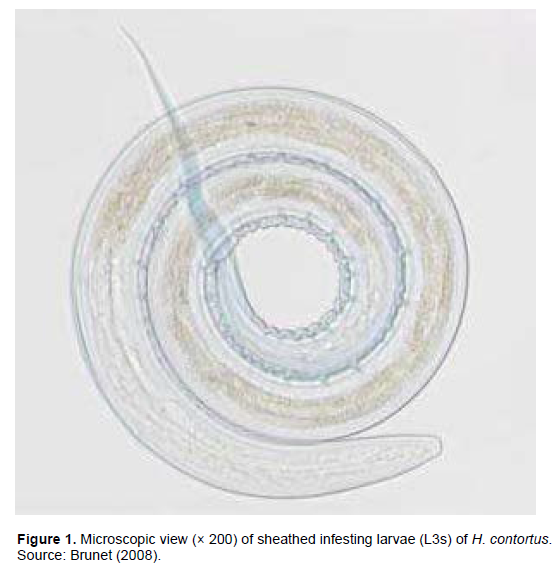
Also called the Barber’s pole worm (Brightling, 2006), Haemonchus is a gender of gastrointestinal parasite belonging to the class of Nematodae, the family of Trichostrongylidae and the sub-family of Haemonchinae. The gender counts three species such as: H. contortus, Haemonchus placei and Haemonchus longistipes. H. contortus (Figure 1) has been reported to affect goats, sheep and cattle (Sutherland and Scott, 2010), H. placei affects mostly cattle (Taylor et al., 2007; Sutherland and Scott, 2010) and H. longistipes affects dromedary (Urquhart et al., 1996). H. contortus is the main species of strongle found in small ruminants in tropical areas of Africa, Central America, Southeast Asia and subtropics in Australia and South America (Alowanou, 2016). Measuring about 15 to 30 mm long, an adult H. contortus male is shorter than the female. H. contortus male is characterized by its copulatory bursa formed of two large lateral lobes and a small asymmetrically positioned dorsal lobe (Morales and Pino, 1987). Female parasites have a reddish digestive tube containing ingested blood, spirally surrounded by two white genital cords (Getachew et al., 2007). H. contortus is a hematophagous strongle located in the abomasum of small ruminants. This characteristic leads to a greater pathogenicity compared to other gastrointestinal nematodes (Penicaud, 2007). In fact, as a blood-sucking parasite, it absorbs the blood of the fine capillary vessels of the digestive mucosa of animals, which can cause more or less severe anemias. In addition, the female is extremely active in terms of spawning with excretion of about 5000 to 7000 eggs/day (Coyne and Smith, 1992).
H. contortus is an extremely prolific parasite possessing different strategies for evading unfavorable environmental conditions and immune reactions of the host. Due to its unique ability for producing eggs in large number during its lifetime, H. contortus has an important advantage over other parasites. In that, it can easily contaminate grazing areas and may survive in its hosts through frequent and rapid re-infections. In addition, because the degree of infectivity varies significantly according to the H. contortus isolates, studies concluded on the importance to take in to account the parasite genetic diversity in various agro-ecological zones (Aumont et al., 2003) in all prevention and control measures. These above factors justify its high pathogenicity which is a requirement to the treatment and control of this parasite in small ruminants (Can, 2015) (Figure 1).
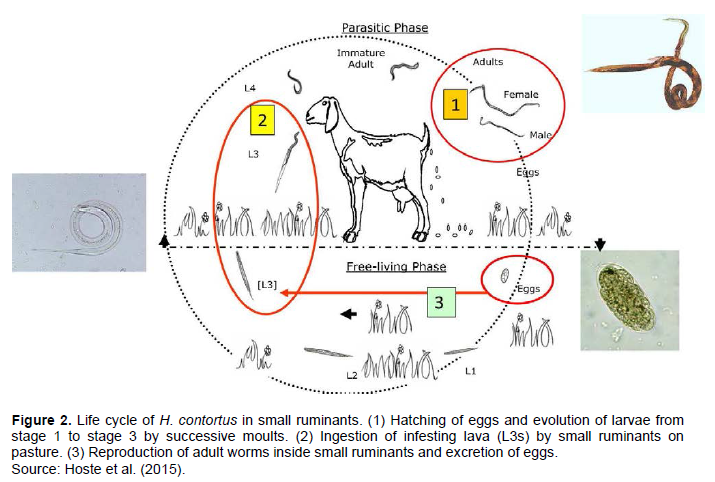
LIFE CYCLE OF H. CONTORTUS
The life cycle of a parasite, like every living thing, describes the whole development process of its life which follows a certain pattern designated under the term life cycle. As regards H. contortus, the life cycle is deemed as a direct life cycle which comprise two phases: a parasitic phase that takes place in the host, and a free-living phase that takes place in the external environment (Walken-Brown et al., 2008; Solaiman, 2010) (Figure 2). According to Ballweber (2004), H. contortus life development may generally take 2 to 4 weeks to complete after infection. Walken-Brown et al. (2008) described H. contortus life cycle in seven stages: the egg stage, four larvae stages (L1, L2, L3, and L4), and two adult stages, although the sexually immature adult stages are sometimes named L5. In the development process, the adult female mates with a male and lays the fertile eggs in the digestive tract of the host. Through defecation, those eggs are freely released into the environment by the host. With favorable environmental conditions, the eggs hatch to free-living L1 (Bush et al., 2001; Brightling, 2006), which at their turn, moult to the L2 stage. Both L1 and L2 stages larva feed on bacteria within the host faeces (Walken-Brown et al., 2008). Then, the L2 stage partially changes into the L3 stage, which is unable to feed on bacteria due to its envelope (Bush et al., 2001). So, the amount of energy left after the L2 stage determines the survival of the L3 larvae (Brightling, 2006). The ingestion of the L3 by the host is then necessary to complete their life cycle. Thus, the L3 larvae leaves the faeces, migrates up grass leaves in the pasture, and remains suspended, during the morning dew (Brightling, 2006). After its ingestion by the host, the L3 larvae then changes into L4 that then enters the abomasum mucous of the host to advance into L5 which later becomes sexually mature in the gastrointestinal tract of the host (Walken-Brown et al., 2008). When adults of H. contortus attains maturity, they mate, and begin laying eggs inducing a new cycle.
FACTORS INFLUENCING H. CONTORTUS DEVELOPMENT
Many previous studies reported external factors that influence the patterns of H. contortus development. Indeed, temperature, rainfall, humidity and vegetation cover are environmental factors which influence Gastrointestinal Nematodes (GINs) development (Selemon, 2018). El-Ashram et al. (2017), early, had revealed a direct correlation between the harshness of gastrointestinal nematodes problems and rainfall during the wet periods of the year where livestock are raised in the developing countries. Furthermore, Attindehou et al. (2012) also reported, in Benin Republic, a significant association of the haemonchosis rates with the season; the minimum and maximum infection rate respectively 36.06% in January (a dry month) and 79.41% July (a very wet month). Definitely, this seasonal trend of prevalence of these parasitic infections will assist in preparing appropriate control strategies, that will be beneficial for goats rearing and industry (Singh et al., 2015). Beside these environmental factors, many other factors have also been reported to influence parasitic infections in small ruminants: the nutrition (Bricarello et al., 2005; Knox et al., 2006), the management practices such as overcrowding, poor management and hygiene (Olanike et al., 2015), the differential management practices (Mandonnet et al., 2003), the drug treatments (Barnes et al., 2001), the genetic factors that provide natural resistance to the host like the breed of host (Chaudary et al., 2007; Saddiqi et al., 2011; Solomon-Wisdom and Matur, 2014; Singh et al., 2015), the age of the host (Solomon-Wisdom et al., 2014; Singh et al., 2015) and the sex of the host (Attindehou et al., 2012; Olanike et al., 2015; Poddar et al., 2017). Contrary to all the above factors, Attindehou et al. (2012) reported no significant difference in relation to animal’s age, origin, sex or species, even if animals less than a year old and especially goats were mostly infected. Finally, both the body weight and reproductive status of the host, according to Tasawar et al. (2010), influence the parasitic infection development due to the development of acquired immunity with gradual increase in weight along with age of the animals.
PREVALENCE OF H. CONTORTUS IN SMALL RUMINANTS
H. contortus is a serious nematode in small ruminants and has been found as a dominant parasite of goat and sheep among the nematodes (Jamalm et al., 2016). Several parasitological surveys carried out in many regions of Africa have shown convincing results regarding the prevalence of gastrointestinal nematodes in small ruminants’ herds. Indeed, in Benin Republic, 55.56% of the examined animals were infested by H. contortus; and the monthly trend of infections showed that in all areas, haemonchosis is endemic with no significant differences in terms of origins or species (Attindehou et al., 2012). According to the same study, the minimum and maximum recorded H. contortus infection rate was respectively of 16.9% in January (a dry month) and 88.7% in July (a very wet month). In Nasarawa State (Nigeria), Adua and Hassan (2016) reported an overall nematodes infection rate of 32.40 and 17.01% in Red Sokoto goats and West African Dwarf goats respectively. According to the same study, the prevalence rate of nematodes infection was 22.45 and 17.82% in Red Sokoto goats while West African Dwarf goats had 14.58 and 8.33% in young and adults respectively. In addition, in the same country, Olanike et al. (2015) reported in Ibadan, 75.85% small ruminants positive for gastrointestinal parasites with the higher prevalence of 54.25% in Red Sokoto breed and the lower prevalence of 21.5% in West African Dwarf breed. According to the results of the same study, 22.75 and 10.5% Red Sokoto and West African Dwarf breeds respectively had mixed helminths (Strongyle spp, Strongyloides spp and Coccidia spp) and protozoa infections (Olanike et al., 2015). In the Plateau region of Togo, Bonfoh et al. (1995) reported a H. contortus prevalence up to 82%. Later, in peri-urban area of Sokodé, in Togo, approximatively the same prevalence rate of gastrointestinal nematodes in small ruminants was recorded (88% represented by Haemonchus sp. and Trichostrongylus sp.) with a negative effect of the season. In a similar way, in urban and peri-urban areas in Maroua, Far North of Cameroon, Ngambia Funkeu et al. (2000) have reported the presence of five species of parasitic nematodes: Haemonchus, Trichostrongylus, Cooperia, Oesophagostomum and Strongyloides papillosus with a predominance of Trichostrongylus and Haemonchus respectively in dry and rainy season. This same study revealed a prevalence of 27 to 31% for these two species depending on the age of the sheep without any significant influence of sex. An epidemiological investigation of small ruminants parasites in the southern forest zone of Ivory Coast carried out by Oka et al. (1999) has revealed a parasite fauna which consisted of nine species of nematodes with a predominance of Trichostrongylus colubriformis (89.7%) and H. contortus (84.1%) in terms of prevalence. Furthermore, in Eastern Ethiopia, Sissay et al. (2007), reported a prevalence of 60% in small ruminants. This is below the results of Mengist et al. (2014) who recorded an overall prevalence of H. contortus of 71.03% with prevalence in sheep and goat up to 67.57 and 71.39% respectively in and around Finoteselam, Ethiopia. According to the same study, the prevalence of haemonchosis was higher in males (73.22%) and adult animals (71.43%). The high rate of prevalence of infection among the goats could be attributed to poor management practices and lack of veterinary services in the area (Osakwe and Anyigor, 2007). A prevalence assessment of H. contortus infections in Goats in Nyagatare District (Rwanda) showed that 75.7% of the small ruminants had H. contortus eggs in faeces with a prevalence rate of 71.8% in goats (Mushonga et al., 2018). Moreover, a 12 months period of survey in the local abattoir of Nyala town, South Darfur State, Sudan revealed 85% of slaughtered goats harbored both adults and immature worms of H. contortus (Abakar, 2002) while an overall prevalence of H. contortus eggs of 12.1% with a 95% CI ranging from 7.97 to 16.23% has been reported in Khartoum State (Sudan) by Boukhari et al. (2016).
Other recent studies conducted on goats in the rest of the world, particularly in Madhya Pradesh (India) concluded that H. contortus was the most predominant parasite followed by Trichostrongylus sp., Oesophagostomum sp., Strongyloides sp. and Bunostomum sp. Of the 960 faecal samples of goats examined, 94.48% were found positive for one or more gastrointestinal parasitism viz., coccidian (82.4%), strongyle (69.27%), amphistomes (22.71%), Strongyloides (9.17%), Trichuris (3.85%), Moniezia (3.02%), Schistosoma (2.29%) and Fasciola sp. 1.77% (Singh et al., 2015). Furthermore, various others studies had earlier reported the high prevalence rates of gastro-intestinal parasites in goats, especially H. contortus, from Indonesia (89.4%) (Widiarso et al., 2018) and different parts of India like 88.23% prevalence of helminthes in Nagpur (Maske et al., 1990), 90.05% from Jabalpur (Lalbiaknungi, 2002), 96% in Tarai region of Uttarakhand (Pant et al., 2009). In Markhor of Chitral Gol National Park, a prevalence rate of 40% of H. contortus has been recorded by Jamalm et al. (2016) against 56-61% prevalence that has been recorded for the parasite in goat in previous studies especially in the Potohar area of Pakistan (Chaudary et al., 2007) and 77.7% Jehangirabad District Khanewal, Punjab, Pakistan recorded by Tasawar et al. (2010). Furthermore, Adhikari et al. (2017) reported a polyparasitism with the higher prevalence for H. contortus of 13.89% in goats of Western Chitwan of Nepal. According to the same study, H. contortus was more prevalent in non-dewormed (40.32%) than in dewormed (5.26%). Finally and in agreement with the previous reports, H. contortus has been reported, in the region of Valle de Lerma (northwestern Argentina), by Suarez et al. (2013) to be the most prevalent nematode species.
EFFECTS OF H. CONTORTUS INFECTIONS ON SMALL RUMINANTS’ PRODUCTION
Information on the effects of H. contortus infections on small ruminant production mostly concern milk production (both the yield and quality). And even in this context, compared to dairy cows, effects of H. contortus infections on dairy goats and sheep are not well documented. However, several decades ago, while comparing milk yield in ewes orally infected with 2500 H. contortus larvae weekly during pregnancy and lactation, Thomas and Ali (1983) reported a striking weight loss and reduction of sheep milk yield by 23%. This result was then greater than 2.5% to 10% milk yield reduction that had been recorded by Hoste and Chartier (1993) from machine-milked goats infected three times with H. contortus L3 larvae at 50-day intervals. But recent studies have revealed greater reduction rates than these previous ones. Indeed, in Italy, a study involving untreated naturally infected and anthelminthic-treated animals has revealed significantly effect of GINs infections on milk production, with the highest milk yield recorded in the treated goats (Rinaldi et al., 2007). More recently, in Argentina, Suarez et al. (2017) reported a significant difference in the mean total milk production between treated (399.5 L ± 34.0 L) and untreated goats (281.6 L ± 37.5 L), amounting to 41.8% increase in total milk yield. The same study also revealed a post-partum peak in egg count and a negative effect of gastrointestinal nematodes (GINs) on milk yield, even with moderate infections. In addition, studies have gone further by assessing the effects of those GINs infections on the lactation length in small ruminants. Considering milk production of the whole period in naturally infected goats in France, Chartier et al. (2000) r eported a significant effect of GINs infections on the lactation period length with a longer duration of lactation in the high protein diet treated group compared to the group treated with normal protein diet (301.5 vs. 294.9 days) and a similar tendency for the total milk yield. According to Suarez et al. (2009), anthelmintic treatment positively affects the length of the milking period with regard to the length of the milking period of untreated dairy sheep. The same way, Suarez et al. (2017) revealed, in goats instead, a significant negative effect of the GIN infections on the milking period length of the goats after kidding (262.3±9.8 days and 223.3±10.8 days respectively for treated and untreated goats). These different results could explain the positive correlation between the GINs infections treatment and the persistence increase in milk yield in dairy goats, ranging from 7.4 to 18.5% with respect to control values observed by Rinaldi et al. (2007). The same study (Rinaldi et al., 2007) highlighted the deteriorating effect on milk quality caused by nematode infections, when they observed that 29.9% lower fat, 23.3% lower protein and 19.6% lower lactose contents in milk from the untreated goats than that from the control group. However, these finding were not in accordance with Hoste and Chartier (1993) who previously had reported no changes in fat and protein contents between infected and uninfected dairy goats. This might be due to the high level of resistance development in the GINs that occurred more recently in small ruminants herds and reported by several studies in small ruminants (Kaplan and Vidyashankar, 2012; Torres-Acosta et al., 2012b; Geurden et al., 2014; Besier et al., 2016). Finally, in Pakistan, Muhammad et al. (2011) estimated the effect of haemonchosis on milk yield and goats weight respectively up to 29 and 27% reduction.
Losses due to H. contortus infections are related to productivity performances, particularly to decrease in body weight that can range from 20 to 60% (Kawano and Yamamura, 2001). These losses could be explained by the loss of appetite (reduction of voluntary feed intake), diarrhoea, anemia and reduced growth (Khan et al., 2008) and disturbance in the nutrient metabolism that cause young H. contortus. In overall, Muhammad et al. (2011) estimated losses due to haemonchosis in sheep and goats at 10-20% reduction of the production.
In disease pathogenesis, anorexia or depression of voluntary feed intake is properly recognized as a critical factor that is capable of revealing largely the response to imbalance of nutrition during gastrointestinal nematodes infection (Sahoo et al., 2011). Even in subclinical infections, anorexia is present (Sykes and Greer, 2003), and may account for around 40 to 90% production losses detected during intestinal parasitism (Greer, 2008). According to Sahoo et al. (2011), in a parasitized animal, anorexia occurrence is as a result of the different factors, viz: a) triggered by the parasite itself for its own advantage; b) reduction of voluntary feed intake is aimed at starving the parasites; c) in the host, it helps in promoting an effective immune response; and d) anorexia affords the host an opportunity to chose diets that minimize infection risk. According to both the nutrient contents of feed offered to parasitized animals and the number of established parasites present, Petkevi?ius (2007) revealed voluntary feed intake reductions varying from 6 to 50% which, according to Greer (2008), could be understood to be the cost of the developing immune response. Feed intake of parasitized animals usually returns toward normality as animals acquire resistance to infection (Sahoo et al., 2011). More recently, on artificial infection with 15 000 third-stage larvae of H. contortus given as three divided doses, Tonin et al. (2014) concluded on progressive degradation of physiological condition; weakness, lethargic and pale state; and depressed feed intake of crossbred Corriedale lambs.
On the other hand, one of the key features of GINs infection, such as H. contortus infection is an increased loss of endogenous protein into the gastrointestinal tract, partly due to plasma protein leakage and partly because of increased production of muco-protein and sloughing of epithelial cells into the alimentary tract (Petkevi?ius, 2007; Sahoo et al., 2011). A substantial amount of these proteins are redigested before absorbtion at sites distal to infection; however, subsequent recycling of digested nutrients would result to additional energy expense by the small ruminants (Knox et al., 2006). The quantity of nutrients reabsorbed endogenously depends on the distal tract (whether there is adequate compensatory absorptive capacity or the lesions position (whether they are in the anterior) (Coop and Kyriazakis, 2001). A proportion that is not resorbed is either further digested in the large intestine or waits to be excreted in the faeces, absorbed as ammonia and excreted as urea in the urine and can therefore constitute a major drain to the infected animals’ overall nitrogen economy (Knox et al., 2006). In parasitized animals, nutrients diversion from production towards specific proteins synthesis for replacement, repair, and reaction to the gut wall damage, to whole blood or plasma loss as well as to mucus production can inflict a significant drain on resources that otherwise would have contributed to fiber, bone, milk and muscle synthesis (Liu et al., 2003; Sahoo et al., 2011). For instance, according to Liu et al. (2003), an additional 17g/day Metabolisable Protein (MP), which is equivalent to 0.57, 0.71, and 0.14 of the MP requirement, is respectively needed for growth, late pregnancy, and early lactation as compensation for losses owing to GINs infection. According to Colditz (2003), GINs adult and larval stages incidence in the gastrointestinal tract leads to inflammation and activation of the acute phase response to infection and occurs locally and systemically. These responses may cause significant drain on the nutritional resources at the disposal of the animals along with protein redirection away from other body processes (Knox et al., 2006).
Finally, the analysis of the situation on the economic plan designates H. contortus as the most economically vital gastrointestinal nematode in its main endemic zones (Perry et al., 2002; Mcleod, 2004) ma inly owing to the common occurrence and potential for substantial rates of mortality in small ruminants. Animal losses vary significantly between seasons, years and regions, contingent on environmental conditions as well as control measures’ effectiveness, including anthelmintic resistance impact (Besier et al., 2016). Although it is difficult to assess the impact of chronic H. contortus infection, and also critically significant in wide grazing situations where routine monitoring is seldom conducted, Muhammad et al. (2011) ascribed considerable loss to the reduced value of animal production. For example, in Australia, gastrointestinal nematodes cost the sheep industry $369 million annually or around 8.7% of its total value (Sackett et al., 2006). All these results revealed the negative interaction between small ruminants and nematode (Hoste et al., 2010), and could justify the fact that, even at moderate burdens, GIN control should not be neglected in small ruminants production.
CONTROL OF GASTROINTESTINAL NEMATODES PARASITES IN SMALL RUMINANTS
Use of chemical anthelmintics
Anthelmintics have continued to be the bedrock of many GIN control programmes in grazing animals owing to their ease of use, low cost, and lack of real alternative options (Kenyon and Jackson, 2012). However, in many countries, the resistance of gastrointestinal parasites to chemical anthelmintic is an increasing burden and poses real concern to numerous countries (Kaplan and Vidyashankar, 2012; Torres-Acosta et al., 2012b; Geurden et al., 2014). Anthelmintics resistance is an increasing challenge not only in small ruminants (Kaplan and Vidyashankar, 2012) but also in cattle (Cotter et al., 2015) and horses (Nielsen et al., 2014). GINs resistance to the three classes of anthelmintics (macrolytic lactones, nicotinic agonists, and benzimidazoles) has become recurrent globally, since the foremost case of resistance was identified in the early 1960s (Fleming et al., 2006; Kaplan and Vidyashankar, 2012; Cotter et al., 2015). Moreover, in single nematode strains, multiple resistance remains a concern (Taylor et al., 2009; Geurden et al., 2014). Nevertheless, GINs resistance levels against anthelmintics may vary between areas (Torres-Acosta et al., 2012b).
As regards anthelmintic resistance of GIN in goats, since the very first reported cases in different areas of the world like New Zealand (Kettle et al., 1983), Australia (Barton et al., 1985), France (Kerboeuf and Hubert, 1985), this challenge has become globally prevalent as in sheep (Fleming et al., 2006; Jackson et al., 2012; Chandra et al., 2015). In Australia and South America, there is particularly high prevalence; however, in Europe there are increasing reports of elevated prevalence (Váradi et al., 2011). Though both goats and sheep are infected with the same nematode species (Hoste et al., 2008), parasites in goats seem to be more resistant to chemical drugs, especially in large flocks characterized by high stocking rates, industrial schemes of production, and frequent treatment based on anthelmintics. Thus, resistance to chemical anthelminthic is assumed to be more frequent in goats’ parasites than in sheep (Váradi et al., 2011). According to Jackson et al. (2012), this low sensitivity to anthelmintics in goats parasites primarily results from difficulties in ascertaining the precise dose of drugs in goats as compared to sheep. A number of anthelmintics are registered for use in sheep, but in goats, they are used off-licence. Thus, goats treatment at the recommended dose rates of sheep led to routine underdosing which reduces the efficacy of the drug used and partly explains the high prevalence of anthelmintic resistance of parasites in goats in comparison with sheep (Hoste et al., 2011).
To retain anthelmintics effectiveness for a prolonged period, a detailed comprehension of the factors that are likely to initiate anthelmintic resistance of GINs is necessary. As a result, there is need for appropriate approaches implementation and development that will slow or impede possible resistance (Leathwick et al., 2015). Resistance development in GINs against anthelmintic may be influenced by many factors, e.g those listed in Table 1.

Alternatives control methods
Elimination of the source of contamination of animals
The purpose of the depletion of the source of contamination is to block the biological cycle of gastrointestinal nematodes (GINs) by controlling the infestation of grazing and thus minimizing the risk of contact between sensitive hosts and L3s larvae (Paolini et al., 2004; Heckendorn, 2007). Various methods of grazing management exist to achieve this goal. These methods are based on three main principles: prevention, evasion and dilution (Pomroy, 2006). Prevention is to put healthy animals on clean pastures (free of L3s) while evasion involves transferring treated animals with anthelmintics from contaminated pastures to clean pastures. Finally, the last principle is to dilute the infestation of grazing.
Improvement of the host resistance
The improvement of the host resistance may be done by two ways: selection of genetically resistant animals and improvement of the host diet.
Selection of genetically resistant animals
The selection of animals resistant to gastrointestinal nematodes (GINs) is a long-standing approach to reduce the use of synthetic anthelmintics (Pomroy, 2006), as such selection would theoretically reduce host infestations and gradually decrease pasture contamination. Genetic variability in GINs resistance has been reported either between breeds or between individuals of the same breed (Bishop and Morris, 2007). Selection of resistant animals may also present some limitations such as the risk of increased host susceptibility to other pathogens (Gruner et al., 1998) or an adverse effect on productivity (Stear and Murray, 1994; Gray, 1997). In addition, these resistant animal selections remain long-term programmes that must take into account local breeding conditions, availability of breeds, and breeding objectives (Pomroy, 2006).
Improvement of the host diet
Gastrointestinal nematodes (GINs) cause severe disruption of digestive physiology and induce an increase in the host's dietary requirements to overcome the strong disturbances of protein and energy metabolism (Hoste et al., 2005). On the basis of this observation, it has been suggested that an improvement in the feed ration to cover the additional needs associated with the presence of nematodes would contribute to improving the host's response to parasitism, particularly when corrections are made. In general, it has been shown that protein metabolism is far more affected by gastrointestinal parasitism than energy metabolism (Coop and Kyriazakis, 1999). As a result, the studies have focused on the benefits of protein supplementation. The notion of
immuno-nutrition has been suggested because improving the diet leads to greater resilience by reducing the consequences of subclinical infestations and improved resistance (Hoste et al., 2008).
Use of medicinal plants
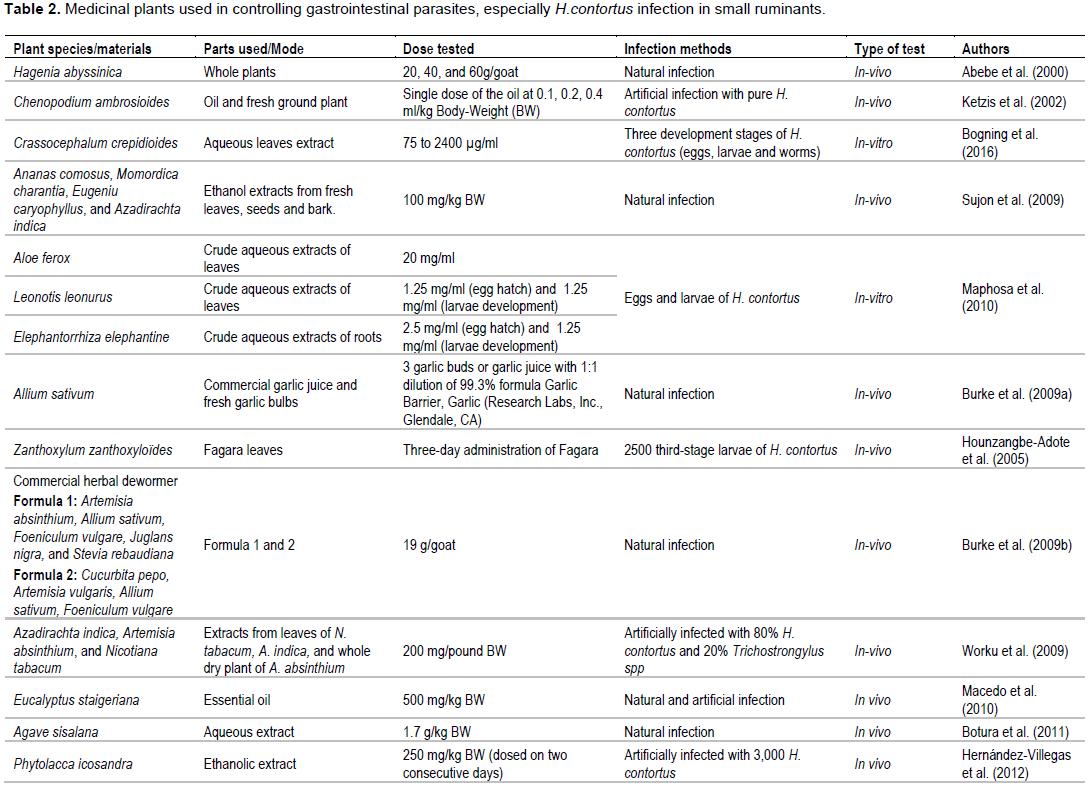
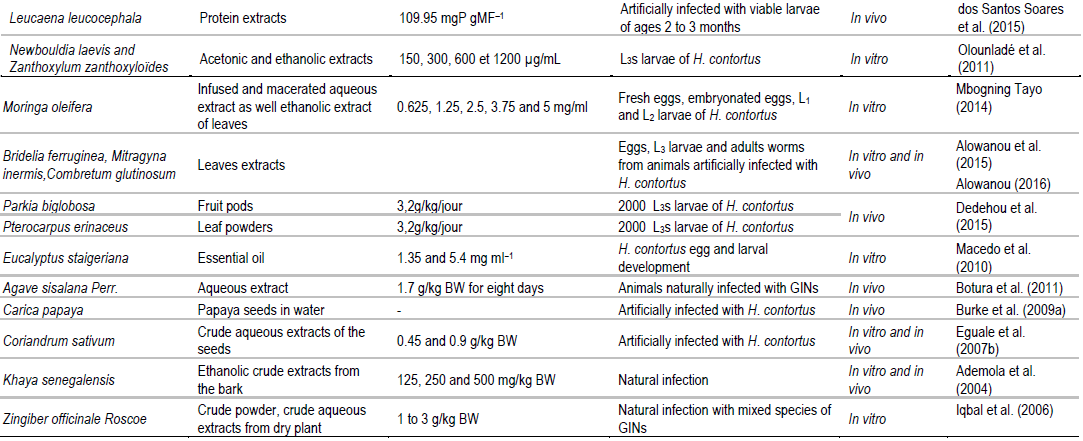
For centuries, medicinal plants and their extracts have been employed in treatment of diseases in man as well as animals (Akhtara et al., 2000; Hounzangbe-Adote et al., 2005; Athanasiadou et al., 2007). Worldwide, anthelmintic resistance occurrence in GIN populations has inspired investigation pertaining to plants and their extracts’ usage as a substitute approach for controlling GINs in ruminants. These medical plants are reasonably inexpensive, generally accepted by small landholders and available locally (Athanasiadou et al., 2007; Hoste et al., 2011). Thus, several review works have already been conducted on use of medicinal plants as a substitutive means for controlling GINs in ruminants (Akhtara et al., 2000; Athanasiadou et al., 2007; Hoste et al., 2011). Table 2 summarizes information on some of these plants that have been used in the recent studies for controlling H. contortus infection in small ruminants.
CONCLUSION
This current study attempted to provide an overall view about the prevalence and the methods of control of gastrointestinal nematodes parasites, particularly H. contortus. Several previous studies have revealed a high prevalence of H. contortus in small ruminants, and in goats in particular all over the world. Its development is favored by many external factors mainly the climatic factors (temperature, humidity, etc.), management practices. The presence of H. contortus in small ruminants is associated to many problems (weight losses and milk yield reduction) that lead to significant economic losses. Conventional control methods used by farmers during decades are no more adequate to address parasite infections in small ruminants considering their negative impacts on cattle and farmers’ benefits. Medicinal plants with anthelmintic properties have been investigated and can be used as alternatives to chemicals especially for small scale farmers. Knowing, understanding and mastering these alternatives methods might help the small ruminants’ value chain actors to design appropriate control programmes adapted to the financial conditions and geographical area of small scale farmers.
CONFLICT OF INTERESTS
The author has not declared any conflict of interests.
ACKNOWLEDGEMENTS
This work was fully funded by the West Africa Agricultural Productivity Project (WAAPP-Benin).
REFERENCES
|
Abakar A (2002). Prevalence of haemonchus contortus in goats in south darfur, sudan: An abattoir survey. 27th World Vetenary Congress, Tunis, 25th to 29th September 2002. |
|
|
Abebe G, Dawson LJ, Detweiler G, Gipson TA, Sahlu T (2000). Hagenia abyssinica (kosso) for internal parasite control in goats Conference held at Debub University, Awassa, Ethiopia, November 10 to 12, 2000. |
|
|
Ademola I, Fagbemi B, Idowu S (2004). Evaluation of the anthelmintic activity of Khaya senegalensis extract against gastrointestinal nematodes of sheep: In vitro and in vivo studies. Veterinary Parasitology 122:151-164. |
|
|
Adhikari K, Bahadur Rana H, Kaphle K, Khanal T, Raut R (2017). Prevalence of haemonchus contortus in goats of western chitwan of nepal. International Journal of Applied Sciences and Biotechnology 5(3):321-325.10.3126/ijasbt.v5i3.18268 |
|
|
Adua MM, Hassan DI (2016). Prevalence of nematode infestation in goats reared in nasarawa state, nigeria Nigerian Journal of Agriculture, Food and Environment 12(3):79-84. |
|
|
Akhtara M, Iqbal Z, Khan MN, Lateef M (2000). Anthelmintic activity of medicinal plants with particular reference to their use in animals in the Indo-Pakistan subcontinent. Small Ruminant Research 38(2):99-107. |
|
|
Alowanou GG (2016). Utilization of Bridelia ferruginea Benth, Mitragyna inermis (willd.) Kuntze et Combretum glutinosum Perr. Ex dc., in the control of Haemonchus contortus in three genetic types of sheep in benin republic. PhD Thesis, Faculty of Agronomic Sciences (FSA), University of Abomey-Calavi (UAC). |
|
|
Alowanou GG, Olounladé AP, Azando EVB, Dedehou VFGN, Daga FD, Hounzangbé-Adoté SM (2015). A review of Bridelia ferruginea, Combretum glutinosum and Mitragina inermis plants used in zootherapeutic remedies in west africa: Historical origins, current uses and implications. Journal of Applied Biosciences 87:8003-8014. |
|
|
Andrea BDW, Davidson R, Conington J, Roughsedge T, Hutchings MR, Villanueva B (2011). Implications of host genetic variation on the risk and prevalence of infectious diseases transmitted through the environment. Genetics 188:683-693. |
|
|
Athanasiadou A, Githiori J, Kyriazakis I (2007). Medicinal plants for helminth parasite control: Facts and fiction. Animal 1(9):1392-1400. |
|
|
Attindehou S, Salifou S, Biaou CF, Gbati OB, Adamou-N'diaye M, Pangui LJ (2012). Epidemiology of haemonchosis in sheep and goats in benin. Journal of Parasitology and Vector Biology 4(2):20-24. 10.5897/JPVB12.012 |
|
|
Aumont G, Gruner L, Hostache G (2003). Comparison of the resistance to sympatric allopatric isolates of haemonchus contortus of black belly sheep in guadeloups. Veterinary Parasitology 116(2):139-150. |
|
|
Ballweber LR (2004). Waterfowl parasites. Seminars in Avian and Exotic Pet Medicine 13:197-205. |
|
|
Barnes EH, Dobson RJ, Stein PA, Lejambre LF (2001). Selection of different genotype larva and adult worms for anthelmintic resistance by persistent and short acting avermectin/ milberrycins. International Journal for Parasitology 31(7):720-727. |
|
|
Bartley DJ (2008). Anthelmintic resistance in sheep and goats. Proceedings of the sheep veterinary society Spring meeting, Darlington and Autumn meeting, Lancaster 32:79-83. |
|
|
Barton NJ, Trainor BL, Urie JS, Atkins JW, Pymans MFS, Wolstencroft IR (1985). Anthelmintic resistance in nematode parasites of goats. Australian Veterinary Journal 62(7):224-227. |
|
|
Besier B, Kahn LP, Sargison ND, Van Wyk J (2016). Diagnosis, treatment and management of haemonchus contortus in small ruminants. In Haemonchus contortus and haemonchosis-past, present and future trends 93:181-238. |
|
|
Bishop SC, Morris CA (2007). Genetics of disease resistance in sheep and goats. Small Ruminant Research 70(1):70:48-59. |
|
|
Bogning ZC, Olounlade PA, Alowanou GG, Nguemfo EL, Dongmo AB, Azebaze AGB, S. H-A (2016). In vitro anthelmintic activity of aqueous extract of Crassocephalum crepidioides (benth.) s. Moore on Haemonchus contortus. Journal of Experimental and Integrative Medicine 6:1-7. |
|
|
Bonfoh B, Zinsstag J, Ankers P, Pangui L, K KP (1995). Epidémiologie des nématodes gastro-intestinaux chez les petits ruminants dans la région des plateaux au togo. Revue d'Elevage et de Médecine Vétérinaire des Pays Tropicaux 48(4):321-326. |
|
|
Botura MB, Silva GD, Lima HG, Oliveira JVA, Souza TS, Santos JDG, Branco A, Moreira ELT, Almeida MAO, Batatinha MJM (2011). In vivo anthelmintic activity of an aqueous extract from sisal waste (Agave sisalana perr.) against gastrointestinal nematodes in goats. Veterinary Parasitology 177(1-2):104-110. |
|
|
Boukhari MI, Elfadil AAM, Omer FA, Shuaib YA (2016). Prevalence and risk factors of Haemonchus contortus in sheep in khartoum state, the sudan. Journal of Agriculture and Veterinary Science (IOSR-JAVS). 9(2):77-83. |
|
|
Bricarello PA, Amarante AFT, Rocha RA, Cabral Filho SL, Huntley JF, |
|
|
Houdijk JGM, Abdalla AL, Gennari SM (2005). In?uence of dietary protein supply on resistance to experimental infections with Haemonchus contortus in ile de france and santa ines lambs. Veterinary Parasitology 134(1-2):99-109. |
|
|
Brightling T (2006). Livestock diseases in australia. CH Jerram. Science Publishers. |
|
|
Brunet S (2008). Analyse des mécanismes d'action antiparasitaire de plantes riches en substances polyphénoliques sur les nématodes du tube digestifs des ruminants. Thèse de Doctorat, Université de Toulouse. |
|
|
Burke JM, Wells A, Casey P, Kaplan RM (2009b). Herbal dewormer fails to control gastrointestinal nematodes in goats. Veterinary Parasitology 160(1-2):168-170. |
|
|
Burke JM, Wells A, Casey P, Miller JE (2009a). Garlic and papaya lack control over gastroin testinal nematodes in goats and lambs. Veterinary Parasitology 159(2):171-174. |
|
|
Bush AO, Fernandez JC, Esch GW, Seed JR (2001). Parasitism: The diversity and ecology of animal parasites. Cambridge University Press, Cambridge. |
|
|
Can TV (2015). Effects of dietary protein supplementation and plane of nutrition on the resistance and the resilience of boer goats against artificial Haemonchus contortus infection under confined conditions. School of Agriculture and Food Sciences, The University of Queensland, p. 164. |
|
|
Chandra S, Prasad A, Yadav N, Latchumikanthan A, Rakesh RL, Praveen K, Khobra V, Subramani KV, Misri L, Sankar M (2015). Status of benzimidazole resistance in Haemonchus contortus of goats from different geographic regions of Uttar Pradesh, India. Veterinary Parasitology 208:26-37. |
|
|
Chartier C, Etter E, Hoste H, Pors I, Mallereau M, Broqua C (2000). Effects of the initial level of milk production and of the dietary protein intake on the course of natural nematode infection in dairy goats. Veterinary Parasitology 92(1):1-13. |
|
|
Chaudary FR, Khan MFU, Qayyum M (2007). Prevalence of Haemonchus contortus in naturally infected small ruminants grazing in the Potohar area of Pakistan. Pakistan Veterinary Journal 27(2):73-79. |
|
|
Colditz IG (2003). Metabolic effects of immune activation during gastrointestinal parasitism in sheep. Australian Journal of Experimental Agriculture 43:1437-43. |
|
|
Coop RL, Kyriazakis I (1999). Nutrition-parasite interaction. Veterinary Parasitology 84(3-4):187-204. |
|
|
Coop RL, Kyriazakis I (2001). Influence of host nutrition on the development and consequences of nematode parasitism in ruminants. Trends in Parasitology 17:325-30. |
|
|
Cotter JL, Burgel AV, Besier RB (2015). Anthelmintic resistance in nematodes of beef cattle in southwest western australia. Veterinary Parasitology 207(3-4):276-284. |
|
|
Coyne MJ, Smith G (1992). The development and mortality of the free-living stages of Haemonchus contortus in laboratory culture. International Journal of Parasitology 22(5):641-650. |
|
|
Dedehou NVFG, Awohouedji DYG, Hounzangbe-Adote S, Gangbo F (2015). Effects of administration of Pterocarpus erinaceus leaf powders and Parkia biglobosa fruit pods on Haemonchus contourus-parallel djallonke ovins. Journal de la Société de Biologie Clinique du Bénin 2015:20-26. |
|
|
dos Santos Soares AM, de Araújo SA, Lopes SG, Costa Junior LM (2015). Anthelmintic activity of leucaena leucocephala protein extracts on Haemonchus contortus. Brazilian Journal of Veterinary Parasitology, Jaboticabal 24(4):396-401. |
|
|
Eguale T, Tilahun G, Debella A, Feleke A, Makonnen E (2007b). In vitro and in vivo anthelmintic activity of crude extracts of Coriandrum sativum against Haemonchus contortus. Journal of Ethnopharmacology 110(3):428-433. |
|
|
El-Ashram S, Al Nasr I, Mehmood R, Hu M, He L, Suo X (2017). Haemonchus contortus and ovine host: A retrospective review. International Journal of Advanced Research 5(3):972-999. |
|
|
Emery DL, Hunt PW, Le Jambre LF (2016). Haemonchus contortus: The then and now, and where to from here? International Journal for Parasitology 46(12):755-769. |
|
|
Falzon LC, Menzies PI, Vanleeuwen J, Jones-Bitton A, Shakya KP, Avula J, Jansen JT, Peregrine AS (2013). A survey of farm management practices and their associations with anthelmintic resistance in sheep flocks in ontario, canada. Veterinary Parasitology 114(1):41-45. |
|
|
Falzon LC, O'Neill TJ, Menzies PI, Peregrine AS, Jones-Bitton A, vanLeeuwen J, Mederos A (2014). A systematic review and meta-analysis of factors associated with anthelmintic resistance in sheep. Preventive Veterinary Medicine 117(2):388-402. |
|
|
Fleming SA, Craig T, Kaplan RM, Miller JE, Navarre C, Rings M (2006). Anthelmintic resistance of gastrointestinal parasites in small ruminants. Journal of Veterinary Internal Medicine 20(2):435-444. |
|
|
Getachew T, Dorchies P, Jacquiet P (2007). Trends and challenges in the effective and sustainable control of Haemonchus contortus infection in sheep. Review Parasite 14(1):3-14. |
|
|
Geurden T, Hoste H, Jacquiet P, Traversa D, Sotiraki S, di Regalbono AF, Tzanidakis N, Kostopoulou D, Gaillac C, Privat S, Giangaspero A, Zanardello C, Noé L, Vanimisetti B, Bartram D (2014). Anthelmintic resistance and multidrug resistance in sheep gastro-intestinal nematodes in France, Greece and Italy. Veterinary Parasitology 201(1-2):59-66. |
|
|
Gray GD (1997). The use of genetically resistant sheep to control nematode parasitism. Veterinary Parasitology. 72(3-4):345-366. |
|
|
Greer AW (2008). Trade-offs and bene?ts: Implications of promoting a strong immunity to gastrointestinal parasites in sheep. Parasite Immunology 30(2):123-132. |
|
|
Gruner L, Bouix J, Vu Tien Khang J (1998). Genetic resistance to internal parasitosis: Example of studies engaged in France and Poland. Point Vétérinaire 29:1129-1137. |
|
|
Hassan DI, Mbap ST, Naibi SA (2013). Prevalence of worm infection in yankasa sheep and west african dwarf goats in Lafia town and environs, Nigeria. Journal of Agriculture and Veterinary Science (IOSR-JAVS). 4(4):84-90. |
|
|
Heckendorn F (2007). The control of gastrointestinal sheep nematodes with tanniferous forage plants. Swiss Federal Institute of Technology, Zurich, Suisse. |
|
|
Hernández-Villegas MM, Borges-Argáez R, Rodríguez-Vivas RI, Torres-Acosta JFJ, Méndez-González M, Cáceres-Farfán M (2012). In vivo anthelmintic activity of Phytolacca icosandra against Haemonchus contortus in goats. Veterinary Parasitology 189(2-4):284-290. |
|
|
Hoste H, Chartier C (1993). Comparison of the effects on milk production of concurrent infection with Haemonchus contortus and Trichostrongylus colubriformis in high and low producing dairy goats. American Journal Veterinary Research 54(11):1886-1893. |
|
|
Hoste H, Sotiraki S, Landau S, Jackson F, Beveridge I (2010). Goat-nematode interactions: Think differently. Trends in Parasitology 26(8):376-381. |
|
|
Hoste H, Sotiraki S, Torres-Acosta JFJ (2011). Control of endoparasitic nematode infections in goats. Veterinary Clinics of North America: Food Animal Practice 27(1):163-173. |
|
|
Hoste H, Torres-Acosta J, Sandoval-Castro C, Mueller-Harvey I, Sotiraki S, Louvandini H, Thamsborg S, Terrill T (2015). Tannin containing legumes as a model for nutraceuticals against digestive parasites in livestock. Veterinary Parasitology 212(1-2):5-17. |
|
|
Hoste H, Torres-Acosta JFJ, Alonso-Díaz MA, Brunet S, Sandoval-Castro C, Houzangbe-Adote S (2008). Identification and validation of bioactive plants for the control of gastrointestinal nematodes in small ruminants. Tropical Biomedicine 25(1):56-72. |
|
|
Hoste H, Torres-Acosta JFJ, Paolini V, Aguilar-Caballero A, Etter E, Lefrileux Y, Chartier C, Broqua C (2005). Interactions between nutrition and gastrointestinal infections with parasitic nematodes in goats. Small Ruminant Research 60(1-2):141-151. |
|
|
Hounzangbe-Adote MS, Paolini V, Fouraste I, Moutairou K, Hoste H (2005). In vitro effects of four tropical plants on three life-cycle stages of the parasitic nematode, Haemonchus contortus. Research in Veterinary Science 78(2):155-160. |
|
|
Iqbal Z, Lateef M, Akhtar M, S., Ghayur M, N., Gilani A, H. (2006). In vivo anthelmintic activity of ginger against gastrointestinal nematodes of sheep. Journal of Ethnopharmacology 106(2):285-287. |
|
|
Jackson F, Varady M, Bartley DJ (2012). Managing anthelmintic resistance in goats-can we learn lessons from sheep? Small |
|
|
Ruminant Research 103(1):3-9. |
|
|
Jamalm Q, Jafar S, Shah A (2016). Prevalence of Haemonchus contortus in markhor of chitral gol national park. Department of Zoology, University of Peshawar 40:19-23. |
|
|
Kaplan RM, Vidyashankar AN (2012). An inconvenient truth: Global worming and anthelmintic resistance. Veterinary Parasitology 186(1-2):70-78. |
|
|
Kawano M, Yamamura E (2001). Efeitos do tratamento com atihelmintico em cordeiros naturalmente infectados com helmintos gastrintestinais sobre os parametros hematologicos, ganho de peso e qualidade da carcac¸ a. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 29(2):113-121. |
|
|
Kenyon F, Jackson F (2012). Targeted flock/herd and individual ruminant treatment approaches. Veterinary Parasitology 186:10-17. |
|
|
Kerboeuf D, Hubert J (1985). Benzimidasole resistance in field strains of nematodes from goats in france. Veterinary Record 116:133. |
|
|
Kettle PR, Vlassoff A, Reid TC, Horton CT (1983). A survey of nematode control measures used by milking goat farmers and of anthelmintic resistance on their farms. New Zealand Veterinary Journal 31(8):139-143. |
|
|
Ketzis JK, Taylor A, Bowman DD, Brown DL, Warnick LD, Erb HN (2002). Chenopodium ambrosioides and its essential oil as treatment for Haemonchus contortus and mixed adult-nematode infections in goats. Small Ruminant Research 44(3):193-200. |
|
|
Khan U, Tanveer A, Maqbool A, Masood S (2008). Epidemiological studies of haemonchosis in sheep. Veterinary Archives. 78:243-251. |
|
|
Knox MR, Torres-Acosta JFJ, Aguilar-Caballero AJ (2006). Exploiting the effect of dietary supplementation of small ruminants on resilience and resistance against gastrointestinal nematodes. Veterinary Parasitology 139(4):385-393. |
|
|
Lalbiaknungi K (2002). Epidiological studies on gastro-intestinal parasites of the goats (capra hircus) in rural areas around jabalpur. M.V.Sc. Thesis submitted to Jawaharlal Nehru Krishi Vishwa Vidyalaya, Jabalpur. |
|
|
Leathwick DM, Ganesh S, Waghorn TS (2015). Evidence for reversion towards anthelmintic susceptibility in teladorsagia circumcincta in response to resistance management programmes. International Journal for Parasitology: Drugs and Drug Resistance 5(1):9-15. |
|
|
Liu SM, Masters DG, Adams NR (2003). Potential impact of nematode parasitism on nutrient partitioning for wool production, growth and reproduction in sheep. Australian Journal of Experimental Agriculture 43(12):1409-1417. |
|
|
Macedo ITF, Bevilaqua CML, Oliveira LMB, Camurca-Vasconcelos ALF, Vieira L, Fabrício R. Oliveira FR, Queiroz-Junior EM, Tomé A, Nascimento NRF (2010). Anthelmintic effect of eucalyptus staigeriana essential oil against goat gastrointestinal nematodes. Veterinary Parasitology 173(1-2):93-98. |
|
|
Mandonnet N, Ducrocq V, Arquet R, Aumont G (2003). Mortality of creole kids during infection with gastrointestinal strongyles: A survival analysis. Journal of Animal Science 81(10):2401-2408. |
|
|
Maphosa V, Masika PJ, Bizimenyera ES, Eloff JN (2010). In-vitro anthelminthic activity of crude aqueous extracts of Aloe ferox, Leonotis leonurus and Elephantorrhiza elephantina against Haemonchus contortus. Tropical Animal Health and Production 42(2):301-307. |
|
|
Maske DK, Bhilegaonkar NG, Sardey MR (1990). Prevalence of parasitic infection in domestic animals at nagpur. Journal of Veterinary Parasitology 4(2):23-25. |
|
|
Mbogning Tayo G, Poné JW, Komtangi MC, Yondo J, Ngangout AM, Mbida M (2014). Anthelminthic activity of Moringa oleifera leaf extracts evaluated in-vitro on four developmental stages of Haemonchus contortus from goats. American Journal of Plant Sciences 5:1702-1710. |
|
|
Mcleod R. (2004). Economic impact of worm infections in small ruminants in south east asia, india and australia. In: Worm control of small ruminants in tropical asia. In Aciar monograph 113, edited by Gray G.D and Baker R.L.eds Sani R. A., 23-33 |
|
|
Mengist Z, Abebe N, Getachew G, Kumar N (2014). Assessment of small ruminant haemonchosis and its associated risk factors in and around Finoteselam, Ethiopia. Journal of Agriculture and Veterinary Science (IOSR-JAVS) 7(12):36-41. |
|
|
Morales G, Pino LA (1987). Ecoepidemilogy of Haemonchus contortus bahiensis, ecotype present in sheep of arid zones of Venezuela. Memorias do Instituto Oswaldo Cruz 82(3):359-369. |
|
|
Muhammad F, Azhar M, Nisar A (2011). Economic losses due to haemonchosis in sheep and goats. Sciences Internationales. (Lahore) 23(4):321-324. |
|
|
Mushonga B, Habumugisha D, Kandiwa E, Madzingira O, Samkange A, Segwagwe BE, Jaja IF (2018). Prevalence of haemonchus contortus infections in sheep and goats in nyagatare district, rwanda. Journal of Veterinary Medicine, 9 p. |
|
|
Ngambia Funkeu R, Pandey VS, Dorny P, Killanga S (2000). Epidemiological study of gastrointestinal nematodes in sheep in urban and peri-urban areas in maroua, far north of cameroon. Revue d'Elevage et de Médecine Vétérinaire des Pays Tropicaux 53(1):17-22. |
|
|
Nielsen MK, Reinemeyer CR, Donecker LM, Leathwick MD, Marchiondo AA, Kaplan RM (2014). Anthelmintic resistance in equine parasites - current evidence and knowledge gaps. Veterinary Parasitology 204(1-2):55-63 |
|
|
Oka CK, Zinsstag J, Pandey VS, Fofana F, N'Depo A (1999). Epidemiology of ovine parasites in the southern forest zone of Côte d'Ivoire. Journal of Livestock and Veterinary Medicine in Tropical Countries 52(1):39-46. |
|
|
Olanike AO, Olayide AJ, Oludunsin FO, Aderoju OR, Japhe DW (2015). Prevalence of gastrointestinal parasites of goats in Ibadan, southwest, Nigeria. World Journal of Agricultural Research 3(2):49-51. 10.12691/wjar-3-2-2 |
|
|
Olounladé PA, Hounzangbé-Adoté MS, Azando EVB, Tam Ha TB, Brunet S, Moulis C, Fabre N, Fouraste I, Hoste H, Valentin A (2011). Etude in vitro de l'effet des tanins de Newbouldia laevis et de Zanthoxylum zanthoxyloïdes sur la migration des larves infestantes de Haemonchus contortus. International Journal of Biology and Chemistry Sciences. 5(4):1414-1422. |
|
|
Osakwe II, Anyigor SI (2007). Prevalence of gastrointestinal helminths in west african dwarf (wad) goats in an agrarian agro-ecosystem. Animal Research International 4:728-732. |
|
|
Pant K, Rajput MKS, Kumar J, Sahu S, Rajkumari V, Gangwar P (2009). Prevalence of helminthes in small ruminants in Tarai region of Uttarakhand. Veterinary World 2(7):265-266. |
|
|
Paolini V, Fouraste I, Hoste H (2004). In vitro effects of three woody plant and sainfoin extracts on 3rd stage larvae and adult worms of three gastrointestinal nematodes. Parasitology 129(1):69-77. |
|
|
Penicaud JM (2007). Comparaison de la réponse anticorps systémique des ovins barbados black belly et inra 401 lors d'infestations expérimentales par haemonchus contortus (nematoda : Trichostrongylidae). Thèse de doctorat, Ecole Nationale vétérinaire de l'Université Paul-Sabatier de Toulouse (France). |
|
|
Perry B, Randolph R, McDermott J, Sones K, Thornton P (year).Investing in animal health research to alleviate poverty. Edited by, International Livestock Research Institute (ILRI). Nairobi, Kenya: 148p |
|
|
Petkevi?ius S (2007). The interaction between intestinal helminth infection and host nutrition: Review. Veterinarija IR Zootechnika 37(59):73-60. |
|
|
Poddar PR, Begum N, Alim MA, Dey AR, Hossain MS, Labony SS (2017). Prevalence of gastrointestinal helminths of sheep in Sherpur, Bangladesh. Journal of Advanced Veterinary and Animal Research 4(3):274-280. |
|
|
Pomroy WE (2006). Anthelmintic resistance in new zealand: A perspective on recent findings and options for the future. New Zealand Veterinary Journal 54(6):265-270. |
|
|
Rinaldi L, Veneziano V, Cringoli G (2007). Dairy goat production and the importance of gastrointestinal strongyle parasitism. Transactions of the Royal Society of Tropical Medicine and Hygiene 101(8):745-746. |
|
|
Sackett D, Holmes P, Abbott K, Jephcott S, Barber M (2006). Assessing the economic cost of endemic disease on the profitability of australian beef cattle and sheep producers. In MLA Final Report, edited by AHW.087. |
|
|
Saddiqi HA, Jabbar A, Sarwar M, Iqbal Z, Muhammad G, Nisa M, Shahzad A (2011). Small ruminant resistance against gastrointestinal nematodes: A case of Haemonchus contortus. Parasitology Research 109(6):1483-1500. |
|
|
Sahoo A, Khan FA, Karim SA (2011). A review on nutrition and gastro-intestinal nematode parasitsm: Interaction and implication in ruminant livestock. Indian Journal of Small Ruminants 17(1):1-20. |
|
|
Salifou S (1996). Nématodes et nématodoses du tube digestif des petits ruminants du sud bénin: Taxonomie, epidémiologie et les facteurs de variation. PhD Thesis, Université Cheikh Anta Diop de Dakar. |
|
|
Selemon M (2018). Review on control of Haemonchus contortus in sheep and goat. Journal of Veterinary Medicine and Research. 5(5):1139. |
|
|
Singh AK, Das G, Roy B, Nath S, R.Naresh, Kumar S (2015). Prevalence of gastro-intestinal parasitic infections in goat of madhya pradesh, india. Journal of Parasitic Diseases 39(4):716-719. |
|
|
Sissay M, Uggla A, Waller P (2007). Prevalence and seasonal incidence of nematode parasites and fluke infections of sheep and goats in eastern Ethiopia. Tropical Animal Health Production 39(7):521-531. |
|
|
Solaiman SG (2010). Goat science and production. 1st ed, Wiley-Blackwell. |
|
|
Solomon-Wisdom GO, Matur BM, Ibe KC (2014). Prevalence of intestinal helminth infection among sheep and goats raised for slaughtering in Gwagwalada abattoir, Abuja-Nigeria. Journal of Global Pharmaceutical Sciences 02(1):12-19. |
|
|
Stear MJ, Murray M (1994). Genetic resistance to parasitic disease: Particularly of resistance in ruminants to gastrointestinal nematodes. Veterinary Parasitology 54(1-3):161-176. |
|
|
Suarez V, Cristel S, Busetti M (2009). Epidemiology and effects of gastrointestinal nematode infection on milk production of dairy ewes. Parasite 16(2):141-147. |
|
|
Suarez V, Fondraz M, Viñabal A, Martínez G, Salatin A (2013). Epidemiología de los nematodes gastrointestinales en caprinos lecheros en los valles templados Del Noa, Argentina. Revista de Investigaciones Agropecuarias, INTA 39:191-197. |
|
|
Suarez V, Martínez G, Viñabal A, Alfaro J (2017). Epidemiology and effect of gastrointestinal nematodes on dairy goats in Argentina. Onderstepoort Journal of Veterinary Research 84(1):1-5. |
|
|
Sujon MA, Mostofa M, Jahan MS, Das AR, Rob S (2009). Studies on medicinal plants against gastrointestinal nematodes of goats. Bangladesh Journal of Veterinary Medicine 6(2):179-183. |
|
|
Sutherland I, Scott I. (2010). Gastrointestinal nematodes of sheep and cattle biological control. In, edited by Wiley-Blackwell. |
|
|
Sykes AR, Greer AW (2003). Effects of parasitism on nutrition economy of sheep: An overview. Australian Journal of Experimental Agriculture 43(12):1393-1398. |
|
|
Tasawar Z, Ahmad S, Lashari MH, Hayat CS (2010). Prevalence of haemonchus contortus in sheep at research centre for conservation of sahiwal cattle (rccsc) jehangirabad district khanewal, punjab, pakistan. Pakistan Journal of Zoology 42(6):735-739. |
|
|
Taylor MA, Coop RL, Wall RL (2007).Veterinary parasitology. Edited by: Blackwell Publishing Ltd: |
|
|
Taylor MA, Learmount J, Lunn E, Morgan C, Craig BH (2009). Multiple resistance to anthelmintics in sheep nematodes and comparison of methods used for their detection. Small Ruminant Research 86(1-3):67-70. |
|
|
Thomas R, Ali D (1983). The effect of Haemonchus contortus infection on the pregnant and lactating ewes. International Journal for Parasitology 13(4):393-398. |
|
|
Tonin A, Silva AD, Schafer A, Aires A, Oliveira C, Zanini D (2014). Influence of experimental infection by Haemonchus contortus on acetyl cholinesterase activity in lymphocytes of lambs. Experimental Parasitology 139:19-23. |
|
|
Torres-Acosta JFJ, Hoste H (2008). Alternative or improved methods to limit gastro-intestinal parasitism in grazing sheep and goats. Small Ruminant Research 77(2-3):159-173. |
|
|
Torres-Acosta JFJ, Mendoza-de-Gives P, Aguilar-Caballero AJ, Cuéllar-Ordaz JA (2012b). Anthelmintic resistance in sheep farms: Update of the situation in the american continent. Veterinary Parasitology 189(1):89-96. |
|
|
Urquhart GM, Armour J, Duncan JL, Dunn AM, Jennings FW (year). Veterinary parasitology. Edited by 2nd ed. Oxford: |
|
|
Váradi M, Paradopoulos E, Dolinská M, Königová A (2011). Anthelmintic resistance in parasites of small ruminants: Sheep versus goats. Heminthologia 48(3):137-144. |
|
|
Wabo PJ, Yondo J, Fossi TO, Marie CK, Bilong BCF, Mpoame M |
|
|
(2012). The in vitro effects of Chenopodium ambrosioides (chenopodiaceae) extracts on the parasitic nematode Heligmosomoides bakeri (nematoda, heligmosomatidae). Journal of Pharmacognosy and Phytotherapy 3(4):56-62. |
|
|
Walken-Brown WS, Sunduimijid B, Olayemi M, Van Der Werf J, Ruvinsky A (2008). Breeding firbre goats for resistance to worm infection. Rural Industry Research and Development Corporation. |
|
|
Widiarso BP, Kurniasih K, Prastowo J, Nurcahyo W (2018). Morphology and morphometry of Haemonchus contortus exposed to Gigantochloa apus crude aqueous extract. Veterinary World. 11(7):921-925. 10.14202/vetworld.2018.921-925 |
|
|
Worku M, Franco R, Miller JH (2009). Evaluation of the activity of plant extracts in boer goats. American Journal of Animal and Veterinary Sciences 4(4):72-79. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


