Full Length Research Paper

ABSTRACT
The rapid expansion and reproduction of certain plant species represents one of the biggest problems in aquatic environments, ranging from eutrophication to the limited availability of water for human consumption. Among these plants is water hyacinth (Eichhornia crassipes), a herbaceous hydrophyte often branded the world’s worst aquatic weed due to its invasive aggression, negative impact on aquatic environments, and the cost usually associated with its management. Water hyacinth is a biomass, typically rich in lignocellulosic material and making it a potential raw material for the synthesis of products of industrial and domestic interest; e.g. edible fungi. Among the commonly known edible fungi is Lentinus edodes, a commercial mushroom whose versatile nature as a white rot fungus provides basis for the continued exploration of its biochemical processes during solid state fermentation on various lignocellulosic biomass as potential substrates. The fungus naturally feeds on lignocellulose by secreting various extracellular enzymes responsible for breaking down this organic polymer into simple soluble molecules that the hyphae can absorb and develop into mycelia. In this study, L. edodes was assessed for its ability to grow on water hyacinth and possibly utilizing it as a substrate. When cultured onto this noxious biomass followed by assessment by agar plate-based clearing assay and spectrophotometry, the fungus demonstrated its ability to secrete cellulases, xylanases, pectinases, peroxidases and laccases, thus showing its capabilities to physiologically utilize this hydrophyte as a substrate. If properly optimized, this approach can be remarkably used as a sustainable way to control water hyacinth in Zimbabwe.
Key words: Lentinus edodes, water hyacinth, lignocellulosic biomass, lignocellulolytic enzymes, cellulases, xylanases, pectinases, lignin peroxidases, laccases.
INTRODUCTION
MATERIALS AND METHODS
RESULTS

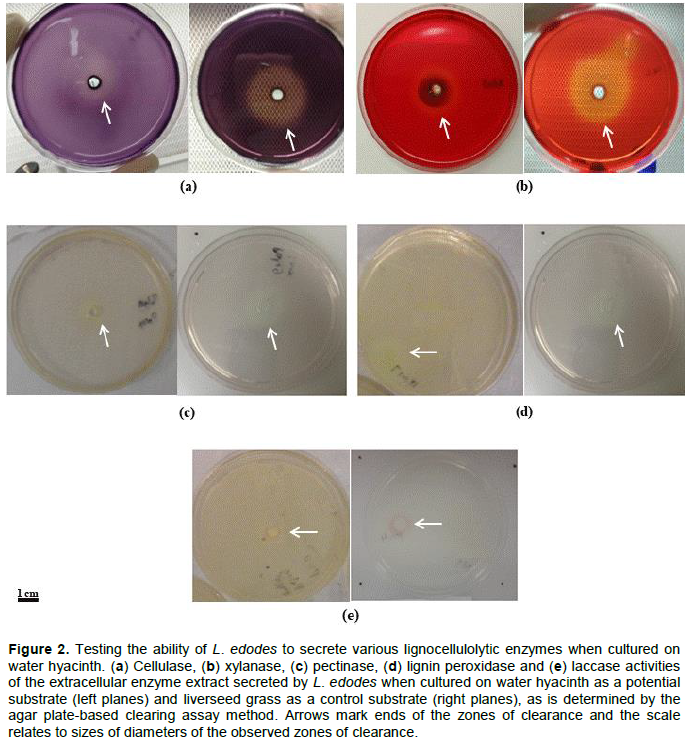
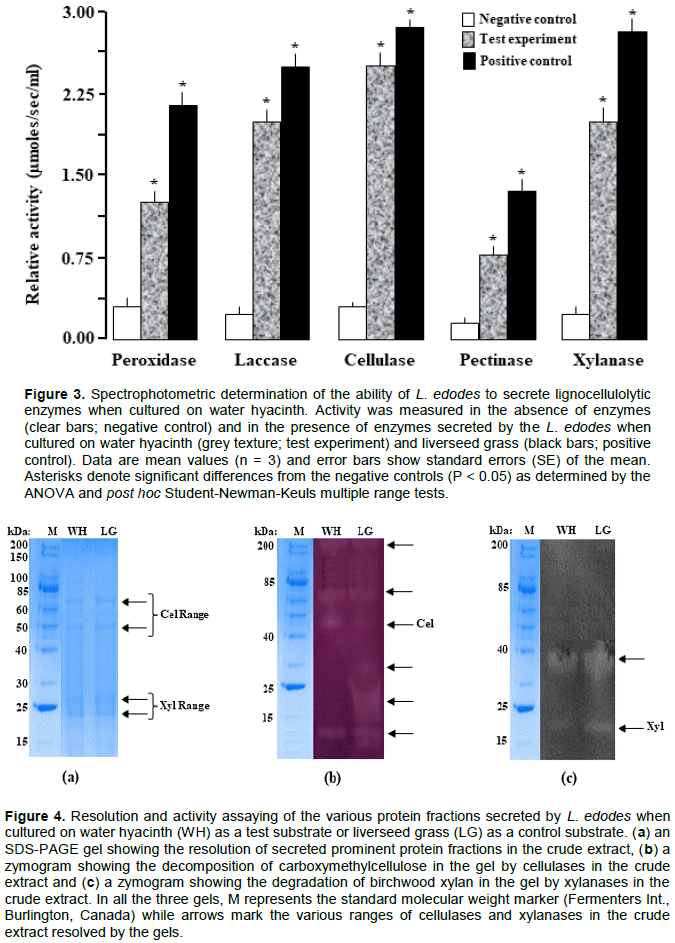
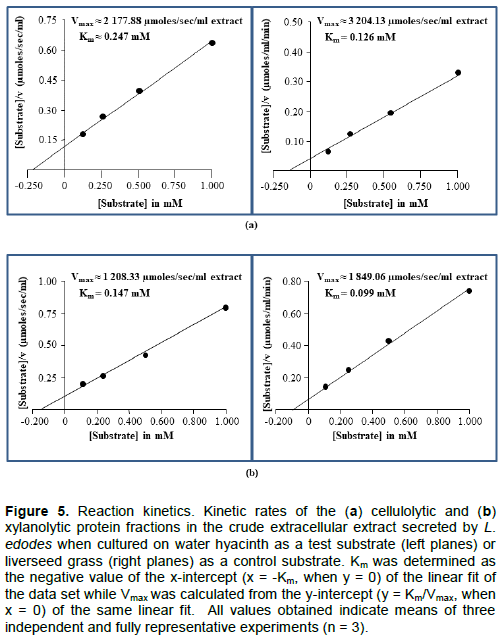
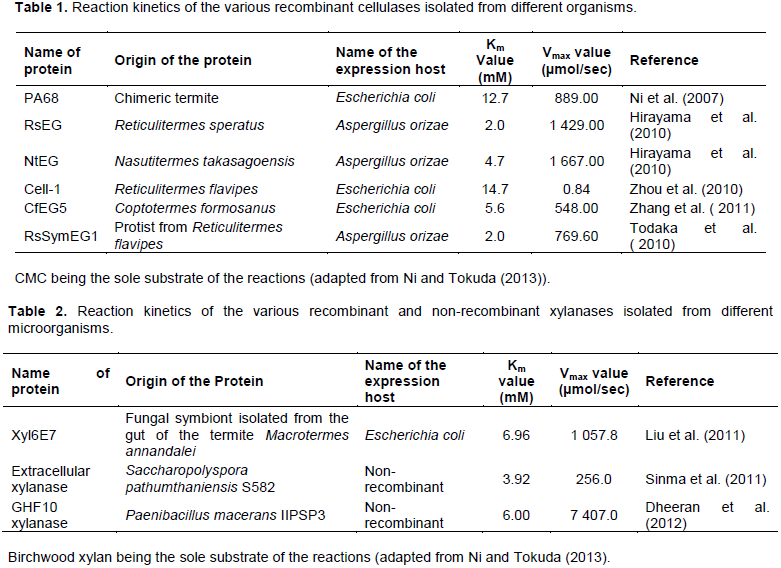
DISCUSSION
CONFLICT OF INTERESTS
The authors declare no competing interests.
ACKNOWLEDGMENTS
REFERENCES
|
Abdalla A, Ambrosano EJ, Vitti SS, Silva FJC (1987). Water-hyacinth (Eichhornia crassipes) in ruminant nutrition. Water Science Technology 19(10):109-112. |
|
|
Avci A, Saha BC, Dien BS, Kennedy GJ, Cotta MA (2013). Response surface optimization of corn stover pretreatment using dilute phosphoric acid for enzymatic hydrolysis and ethanol production. Bioresources Technolology 130:603-612. |
|
|
Baldrian P, Valášková V (2008). Degradation of cellulose by basidiomycetous fungi. FEMS Microbiology Reviews 32:501-521. |
|
|
Bhagobaty RK, Joshi SR, Malik A (2007). Microbial degradation of organophosphorous pesticide: Chlorpyrifos (Mini-Review). International Journal of Microbiology 4:1-5. |
|
|
Bray MR, Clarke AJ (1995). Identification of an essential tyrosyl residue in the binding site of Schizophyllum commune xylanase A. Biochemistry 34:2006-2014. |
|
|
Brown C (2006). Marine and coastal ecosystems and human well-being: Synthesis. United Nations Publications. |
|
|
Buswell J, Cai Y, Chang S (1993). Fungal- and substrate-associated factors affecting the ability of individual mushroom species to utilize different lignocellulosic growth substrates. Conference Proceedings of The First International Conference on Mushroom Biology and Mushroom Products pp. 111-140. |
|
|
ÇaÄŸlarırmak N (2007). The nutrients of exotic mushrooms (Lentinula edodes and Pleurotus species) and an estimated approach to the volatile compounds. Food Chemistry 105:1188-1194. |
|
|
Chen H (2014). Chemical composition and structure of natural lignocellulose. In Biotechnology of lignocellulose. Springer pp. 25-71. |
|
|
Chikwenhere GP (1994). Biological control of water hyacinth (Eichhornia crassipes) in Zimbabwe: Results of a pilot study. FAO Plant Protection Bulletin 42:185-190. |
|
|
Cilliers CJ, Hill MP, Ogwang JA, Ajuonu O (2003). Aquatic weeds in Africa and their control. CABI Publishing. Technology and Engineering 1991:161-178. |
|
|
Cohen R, Persky L, Hadar Y (2002). Biotechnological applications and potential of wood-degrading mushrooms of the genus Pleurotus. Applied Microbiology and Biotechnology 58:582-594. |
|
|
Collins T, Gerday C, Feller G (2005). Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiology Review 29:3-23. |
|
|
Crow GE, Hellquist CB, Fassett NC (2000). Aquatic and wetland plants of northeastern North America pp. 121-150. |
|
|
Da Vinha FNM, Gravina-Oliveira MP, Franco MN, Macrae A, da Silva Bon EP, Nascimento RP, Coelho RRR (2011). Cellulase production by Streptomyces viridobrunneus SCPE-09 using lignocellulosic biomass as inducer substrate. Applied Biochememistry and Biotechnology 164:256-267. |
|
|
Dada SAO, Akinsoyinu AO, Smith JW, Dashiell KE (2002). The effect of leaf-pruning on nutrient intake and in vivo digestibility of soybean stovers by sheep. Journal of Sustainable Agriculture 19:5-14. |
|
|
Dheeran P, Nandhagopal N, Kumar S, Jaiswal YK, Adhikari DK (2012). A novel thermostable xylanase of Paenibacillus macerans IIPSP3 isolated from the termite gut. Journal of Industrial Microbiology and Biotechnology 39:851-860. |
|
|
de Segura BG, Fevre M (1993). Purification and characterization of two 1, 4-beta-xylan endohydrolases from the rumen fungus Neocallimastix frontalis. Applied Environmental Microbiology 59(11):3654-3660. |
|
|
Dorado J, Field JA, Almendros G, Sierra-Alvarez R (2001). Nitrogen-removal with protease as a method to improve the selective delignification of hemp stemwood by the white-rot fungus Bjerkandera sp. strain BOS55. Applied Microbiology and Biotechnology 57:205-211. |
|
|
Eichlerová I, Šnajdr J, Baldrian P (2012). Laccase activity in soils: Considerations for the measurement of enzyme activity. Chemosphere 88:1154-1160. |
|
|
Elisashvili V, Kachlishvili E, Asatiani MD (2015). Shiitake medicinal mushroom, Lentinus edodes (higher basidiomycetes) productivity and lignocellulolytic enzyme profiles during wheat straw and tree leaf bio-conversion. International Journal of Medicinal Mushrooms 17:77-86. |
|
|
El-Sayed AFM (2003). Effects of fermentation methods on the nutritive value of water hyacinth for Nile tilapia Oreochromis niloticus (L.) fingerlings. Aquaculture 218:471-478. |
|
|
Favaro L, Jooste T, Basaglia M, Rose SH, Saayman M, Görgens JF, van Zyl WH (2013). Designing industrial yeasts for the consolidated bioprocessing of starchy biomass to ethanol. Bioengineered 4(2):97-102. |
|
|
Fujimoto H, Ooi T, Wang SL, Takizawa T, Hidaka H, Murao S, Arai M (1995). Purification and properties of three xylanases from Aspergillus aculeatus. Bioscience, Biotechnology and Biochemistry 59(3):538-540. |
|
|
Gopal B (1987). Water Hyacinth. Aquatic Plant Studies 1:471. |
|
|
Gopal B, Goel U (1993). Competition and allelopathy in aquatic plant communities. The Botanical Review 59:155-210. |
|
|
Gunnarsson CC, Petersen CM (2007). Water hyacinths as a resource in agriculture and energy production: A literature review. Waste Management 27:117-129. |
|
|
Gutierrez EL, Ruiz EF, Uribe EG, Martinez JM (2001). Biomass and productivity of water hyacinth and their application in control programs. Biological and integrated control of water hyacinth, Eichhornia crassipes. ACIAR Proceedings 102:109-199. |
|
|
Hirayama K, Watanabe H, Tokuda G, Kitamoto K, Arioka M (2010). Purification and characterization of termite endogenous β-1,4-endoglucanases produced in Aspergillus oryzae. Bioscience, Biotechnology and Biochemistry 74:1680-1686. |
|
|
Howard RL, Abotsi E, Jansen Van Rensberg EL (2013). Development of a fungal cellulolytic enzyme combination for use in bioethanol production using Hyparrhenia spp as a source of fermentable sugars. PhD Thesis. University of Limpopo P 239. |
|
|
Idrees M, Adnan A, Sheikh S, Qureshi FA (2013). Optimization of dilute acid pre-treatment of water hyacinth biomass for enzymatic hydrolysis and ethanol production. EXCLI Journal 12:30-40. |
|
|
Irving HR, Wheeler JI, Iacuone S, Kwezi L, Gehring C, Thompson PE, Ruzvidzo O, Govender K (2011). The phytosulfokine (PSK) receptor is capable of guanylate cyclase activity and enabling cyclic GMP-dependent signaling in plants. Journal of Biological Chemistry 286:22580-22588. |
|
|
Jia Z, Chen N, Shi W, Tang X, Xu H (2019). Bioremediation of cadmium-dichlorophen co-contaminated soil by spent Lentinus edodes substrate and its effects on microbial activity and biochemical properties of soil. Journal of Soils and Sediments 17(2):315-325. |
|
|
Jianbo LU, Zhihui FU, Zhaozheng YIN (2008). Performance of a water hyacinth (Eichhornia crassipes) system in the treatment of wastewater from a duck farm and the effects of using water hyacinth as duck feed. Journal of Environmental Sciences 20:513-519. |
|
|
Jurado M, Martinèz ÀT, Martinez MJ, Saparrat MCN (2011). Application of white-rot fungi in transformation, detoxification, or revalorization of agriculture wastes. Second Edition. Comprehensive Biotechnology. pp. 75-104. |
|
|
Kiiskinen L-L, Kruus K, Bailey M, Ylo¨sma¨ki E, Siika-aho M, Saloheimo M (2004). Expression of Melanocarpus albomyces laccase in Trichoderma reesei and characterization of the purified enzyme. Microbiology 150:3065-3074. |
|
|
Kivaisi A, Magingo F, Mamiro B (2003). Performance of Pleurotus flabellatus on water hyacinth (Eicchornia crassipes) shoots at two different temperature and relative humidity regimes. Tanzanian Journal of Science 5:11-25. |
|
|
Koshy J, Nambisan P (2012). Pre-treatment of agricultural waste with pleurotus sp. for ethanol production. International Journal of Plant, Animal and Environmental Sciences 2(2):244-249. |
|
|
Kuhad RC, Gupta R, Singh A (2011). Microbial cellulases and their industrial applications. Enzyme Research 20:11-24. |
|
|
Kushwaha SPS (2012). Remote sensing of invasive alien plant species. In: Bhatt JR, Singh JS, Singh SP, Tripathi RS, Kohli RK, editors. Invasive alien plants: An ecological appraisal for the Indian sub-continent. CABI International; United Kingdom pp. 131-138. |
|
|
Kwezi L, Ruzvidzo O, Wheeler JI, Govender K, Iacuone S, Thompson PE, Irving HR (2011). The phytosulfokine (PSK) receptor is capable of guanylate cyclase activity and enabling cyclic GMP-dependent signaling in plants. Journal of Biological Chemistry 286(25):22580-22588. |
|
|
Laemmli UK (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227(5259):680-685. |
|
|
Leatham GF (1985). Extracellular enzymes produced by the cultivated mushroom Lentinus edodes during degradation of a lignocellulosic medium extracellular enzymes produced by the cultivated mushroom lentinus edodes during degradation of a lignocellulosic medium.Applied and Environmental Microbiology 4:859-867. |
|
|
Legodi LM, Grange DL, Van Rensburg EL, Ncube I (2019). Isolation of cellulose degrading fungi from decaying banana pseudo stem and Strelitzia alba. Enzyme Research 2019:1-10. |
|
|
Liming X, Xueliang S (2004). High-yield cellulase production by Trichoderma reesei ZU-02 on corn cob residue. Bioresource Technology 91:259-262. |
|
|
Lindsey K, Hirt HM (1999). Use water hyacinth! A practical hanbook of uses for the water hyacinth from across the world. Anamed: Winnenden P 114. |
|
|
Liu S, Liu L, Uzuner U, Zhou X, Gu M, Shi W, Zhang Y, Dai SY, Yuan JS (2011). HDX-analyzer: A novel package for statistical analysis of protein structure dynamics. BMC Bioinformatics 12(1):S43. |
|
|
Magadza CHD (2003). Lake Chivero: A management case study. Lakes and Reserviors Research and Management 8:69-81. |
|
|
Maganhotto de Souza Silva CM, Soares de Melo I, Roberto de Oliveira P (2005). Ligninolytic enzyme production by Ganoderma spp. Enzyme and Microbial Technology 37:324-329. |
|
|
Mako AA, Babayemi OJ and Akinsoyinu AO (2011). An evaluation of nutritive value of water hyacinth (Eichhornia crassipes Mart. Solms-Laubach) harvested from different water sources as animal feed. Livestock Research for Rural Development 23(103). |
|
|
Meier S, Ruzvidzo O, Morse M, Donaldson L, Kwezi L, Gehring C (2010). The Arabidopsis wall associated kinase-like 10 gene encodes a functional guanylyl cyclase and is co-expressed with pathogen defense related genes. PLoS One 5. |
|
|
Men LT, Yamasaki S, Caldwell JS, Yamada R, Takada R, Taniguchi T (2006). Effect of farm household income levels and rice-based diet or water hyacinth (Eichhornia crassipes) supplementation on growth/cost performances and meat indexes of growing and finishing pigs in the Mekong Delta of Vietnam. Animal Science Journal 77:320-329. |
|
|
Mikiashvili N, Wasser SP, Nevo E, Elisashvili V (2006). Effects of carbon and nitrogen sources on Pleurotus ostreatus ligninolytic enzyme activity. World Journal of Microbiology and Biotechnology 22(9):999-1002. |
|
|
Miller GL (1959). Use of dinitrosalicylic acid reagent for determination of reducing sugars. Analytical Chemistry 31:426-428. |
|
|
Moyo P, Mapira J (2012). Bio-remediation with water hyacinth (Eichhornia crassipes): A panacea for river pollution in the city of Masvingo (Zimbabwe). Journal of Sustainable Development in Africa 14:115-131. |
|
|
Murugesan K, Arulmani M, Nam IH, Kim YM, Chang YS, Kalaichelvan PT (2006). Purification and characterization of laccase produced by a white rot fungus Pleurotus sajor-caju under submerged culture condition and its potential in decolorization of azo dyes. Applied Microbiology and Biotechnology 72:939-946. |
|
|
Mwangi T (2013). Water hyacinth - Can its aggressiveness be controlled? Ecosystem Management-UNEP/DEWA, Nairobi, Kenya. |
|
|
MWBP/RSCP (2006). Invasive alien species in the lower Mekong basin: Current state of play. Mekong Wetland Biodiversity Programme and Regional Species Conservation Programme, The World Conservation Union (IUCN), Asia, Sri Lanka. |
|
|
Nayebyazdi N, Salary M, Ghanbary MAT, Ghorbany M, Bahmanyar MA (2012). Investigation of cellulase activity in some soil borne fungi isolated from agricultural soils. Annals of Biological Research 3(12):5705-5713. |
|
|
Ncube T, Howard RL, Abotsi EK, van Rensburg ELJ, Ncube I (2012). Jatropha curcas seed cake as substrate for production of xylanase and cellulase by Aspergillus niger FGSCA733 in solid-state fermentation. Industrial Crops and Products 37:118-123. |
|
|
Ni J, Tokuda G, Takehara M, Watanabe H (2007). Heterologous expression and enzymatic characterization of beta-glucosidase from the drywood-eating termite, Neotermes koshunensis. Applied Entomology and Zoology 42:457-463. |
|
|
Nigam P, Singh D (2002). Enzyme and microbial systems involved in starch processing. Enzyme and Microbial Technology 22:375-407 |
|
|
Parsons WT, Parsons WT, Cuthbertson EG, Cuthbertson EG (2001). Noxious weeds of Australia (CSIRO Publishing) Available at: |
|
|
Pandya B, Albert S (2014). Evaluation of Trichoderma reesei as a compatible partner with some white rot fungi for potential bio-bleaching in paper industry. Annals of Biological Research 5:43-51. |
|
|
Penfound WT, Earle TT (1948). The biology of water hyacinth. Ecological Monographs 18:447-472. |
|
|
Pointing SB (1999). Qualitative methods for the determination of lignocellulolytic enzyme production by tropical fungi. Fungal Diverssity 2:17-33. |
|
|
Polizeli M, Rizzatti A, Monti R, Terenzi H, Jorge J, Amorim D (2005). Xylanases from fungi: Properties and industrial applications. Applied Microbiology and Biotechnology 67(5):577-591. |
|
|
Punniavan S (2012). Effect of medium composition and ultrasonication on xylanase production by Trichoderma harzianum MTCC 4358 on novel substrate. African Journal of Biotechnolology 11:12067-12077. |
|
|
Ramanjaneyulu G, Reddy GPK, Kumar KD, Reddy BR (2015). Isolation and screening of xylanase producing fungi from forest soils. International Journal of Current Microbiology and Applied Sciences 4:586-591. |
|
|
Rashid M, Iqbal M (2012). Effect of phosphorus fertilizer on the yield and quality of maize (Zea mays L) fodder on clay loam soil. Journal of Animal and Plant Sciences 22(1):199-203. |
|
|
Reddy KR, D'Angelo EM (1990). Biomass yield and nutrient removal by water hyacinth (Eichhornia crassipes) as influenced by harvesting frequency. Biomass 21:27-42. |
|
|
Ritter CET, Camassola M, Zampieri D, Silveira MM, Dillon AJP (2013). Cellulase and xylanase production by Penicillium echinulatum in sub-merged media containing cellulose amended with sorbitol. Enzyme Research 2013:1-9. |
|
|
Saha P, Alam MF, Baishnab AC, Khan MR, Islam MA (2014). Fermentable sugar production and separation from water hyacinth using enzymatic hydrolysis. Sustainable Energy 2:20-24. |
|
|
Saito T, Hong P, Kato K, Okazaki M, Inagaki H, Maeda S, Yokogawa Y (2003). Purification and characterization of an extracellular laccase of a fungus (family Chaetomiaceae) isolated from soil. Enzyme and Microbial Technology 33:520-526. |
|
|
Sakthiselvan P, Naveena B, Partha N (2014). Molecular characterization of a xylanase-producing fungus isolated from fouled soil. Brazilian Journal of Microbiology 45(4):1293-1302. |
|
|
Shawky BT, Mahmoud MG, Ghazy EA, Asker MM, Ibrahim GS (2011). Enzymatic hydrolysis of rice straw and corn stalks for monosugars production. Journal of Genetic Engineering and Biotechnology 9(1):59-63. |
|
|
Sibanda A, Ruzvidzo O, Ncube I, Ncube T (2019a). Diversity of cellulase- and xylanase-producing filamentous fungi from termite mounds. Journal of Yeast and Fungal Research 10(2):15-29. |
|
|
Sibanda N, Ruzvidzo O, Zvidzai CJ, Mashingaidze AB, Murungweni C (2019). Exploring for the possibility of utilizing Pleurotus ostreatus to manage Eichhornia crassipes in Zimbabwe. Journal of Yeast and Fungal Research 10(1):1-14. |
|
|
Sinma K, Khucharoenphaisan K, Kitpreechavanich V, Tokuyama S (2011). Purification and characterization of a thermostable xylanase from Saccharopolyspora pathumthaniensis S582 isolated from the gut of a termite. Bioscience, Biotechnology and Biochemistry 75:1957-1963. |
|
|
Silva CHC, Puls J, Sousa MV, Ferreira-Filho EX (1999). Purification and characterization of a low molecular weight xylanase from solid-state cultures of Aspergillus fumigatus Fresenius. Revista de Microbiologia 30:114-119. |
|
|
Sornvoraweat B, Kongkiattikajorn J (2010). Separated hydrolysis and fermentation of water hyacinth leaves for ethanol production. KKU Research Journal 15:794-802. |
|
|
Stajić M, Persky L, Friesem D, Hadar Y, Wasser SP, Nevo E, Vukojević J (2006). Effect of different carbon and nitrogen sources on laccase and peroxidases production by selected Pleurotus species. Enzyme and Microbial Technology 38:65-73. |
|
|
Téllez-téllez M, Díaz R, Sánchez C, Díaz-godínez G (2013). Hydrolytic enzymes produced by Pleurotus species. African Journal of Microbial Research 7:276-281. |
|
|
Thakur M (2018). Mushrooms as a biological tool in myco-remediation of polluted soils. Emerging Issues in Ecology and Environmental Science. Briefs in Environmental Science 15:27-42. |
|
|
Tham HT (2012). Water hyacinth (Eichhornia crassipes) - Biomass production, esilability and feeding value to growing cattle pp. 115-132. |
|
|
Todaka N, Lopez CM, Inoue T, Saita K, Maruyama J, Arioka M (2010). Heterologous expression and characterization of an endoglucanase from a symbiotic protist of the lower termite, Reticulitermes speratus. Applied Biochemistry and Biotechnology 160:1168-1178. |
|
|
Usha KY, Praveen K, Reddy BR (2014). Enhanced production of ligninolytic enzymes by a mushroom Stereum ostrea. Biotechnology Research International 2014:815-849. |
|
|
Villamagna AM, Murphy BR (2010). Ecological and socio-economic impacts of invasive water hyacinth (Eichhornia crassipes): A review. Freshwater Biology 55:282-298. |
|
|
Virabalin R, Kositsup B, Punnapayak H (1993). Leaf protein concentrate from water hyacinth. Journal of Aquatic Plant Management 31:207-209. |
|
|
Vivekanandan KE, Sivaraj S, Kumaresan S (2014). Characterization and purification of laccase enzyme from Aspergillus nidulans CASVK3 from vellar estuary South East Coast of India. International Journal of Current Microbiology and Applied Sciences 3(10):213-227. |
|
|
Wang X, Ding Y, Gao X, Liu H, Zhao K, Gao Y, Qui L (2019). Promotion of the growth and plant biomass degrading enzymes production in solid-state cultures of Lentinula edodes expressing Vitreoscilla hemoglobin gene. Journal of Biotechnology 302:42-47. |
|
|
Wesenberg D, Kyriakides I, Agathos SN (2003). White-rot fungi and their enzymes for the treatment of industrial dye effluents. Biotechnology Advances 22:161-187. |
|
|
Yagi F, Minami Y, Yamada M, Kuroda K, Yamauchi M (2019). Development of animal feeding additives from mushroom waste media of Shochu lees. International Journal of Recycling of Organic Waste in Agriculture 8(2):215-220. |
|
|
Yopi, Tasia W, Melliawati R (2014). Cellulase and xylanase production from three isolates of indigenous endophytic fungi. International Journal for Qualitative Research 8:73-86. |
|
|
Yuan SF, Wu TH, Lee HL, Hsieh HY, Lin WL, Yang B, Huang CH (2015). Biochemical characterization and structural analysis of a bifunctional cellulase/xylanase from Clostridium thermocellum. Journal of Biological Chemistry 290(9):5739-5748. |
|
|
Zhang DH, Lax AR, Bland JM, Allen AB (2011). Characterization of a new endogenous endo-β-1,4-glucanase of Formosan sub-terranean termite (Coptotermes formosanus). Insect Biochemistry and Molecular Biology 41:211-218. |
|
|
Zhang XZ, Zhang YHP (2013). Cellulases: Characteristics, sources, production, and applications. Bioprocessing Technology in Biorefinery for Sustainable Production of Fuels and Chemical Polymers 8:131-146. |
|
|
Zhou X, Kovaleva ES,Wu-Scharf D, Campbell JH, Buchman GW, Boucias DG (2010). Production and characterization of a recombinant β-1,4-endoglucanase (glycohydrolase family 9) from the termite Reticulitermes flavipes. Archives of Insect Biochemistry and Physiology 74:147-162. |
|
|
Zirbes M, Waldvogel SR (2018). Electro-conversion as sustainable method for the fine chemical production from the biopolymer lignin. Current Opinion in Green and Sustainable Chemistry 14:19-25. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


