Full Length Research Paper

ABSTRACT
INTRODUCTION
Finger millet (Eleusine coracana (L.) Gaertn), believed to be adapted to the arid and semi-arid regions of the world, is highly tolerant to pests, diseases and drought (Gowda et al., 2015). It serves as a food and nutritional security crop because of its adaptability to a wide range of environments, high nutritional value, excellent storage qualities and its importance as a low input crop (Dida et al., 2007). It is cultivated on more than 31 million ha of land globally (FAO, 2014).
Finger millet straw is used for livestock feed in manycountries, however, it is mainly grown for food (Upadhyaya et al., 2006) and also for brewing of gluten free beverages (Shayo et al., 2001; Fernandez et al., 2003; Bano et al., 2015). Finger millet grain is nutritionally rich as it contains high levels of protein and minerals (Upadhyaya et al., 2006). David et al. (2014) reported that the proximate composition of finger millet to be moisture (6.99%), ash (2.37%), crude protein (10.28%), crude fibre (3.10%), crude lipid (0.83%), carbohydrate (76.43%) and mineral like potassium (14.19 mg/g), sodium (6.86 mg/g), copper (0.10 mg/g), calcium (1.13 mg/g), magnesium (6.25 mg/g), zinc (0.22 mg/g), manganese (0.32 mg/g), iron (0.11 mg/g) and lead (0.001 mg/g). It contains appreciable amount of the essential amino acid methionine which is lacking in most food grains (National Academies, 1996). Finger millet is suitable to design and develop value-added nutritive food products. Woldemichael and Admasu (2017) reported germination to be effective in starch and protein hydrolysis; while fermentation was more effective in reducing phytate, consequently increasing mineral bioavailability.
In Ethiopia, finger millet is produced by 1.6 million farmers on 454662.33 ha of land (CSA, 2014). Appreciable variability in Ethiopian finger millet genotypes has been documented by various workers (Bezaweletaw et al., 2006; Lule et al., 2012; Wolie, 2013). Although about 15 improved finger millet varieties have been developed in Ethiopia (MoANR, 2016), and some of these varieties were tested for adaptability and introduced to farmers in the north western part of Ethiopia (Fentie, 2012); the use of these varieties in the north eastern part of Ethiopia is not common. Hence, there is a need to study the adaptability of finger millet varieties in order to recommend varieties in that part of Ethiopia. The objectives of this experiment were to identify adaptable improved finger millet varieties and to assess the magnitude of genotype-environment interaction in north eastern part of Ethiopia.
MATERIALS AND METHODS
The experiment was conducted on three locations for two years, 2013 and 2014. The locations were Sirinka, Kobo and Jari, which represent the finger millet growing areas of North Eastern Ethiopia (Table 1).
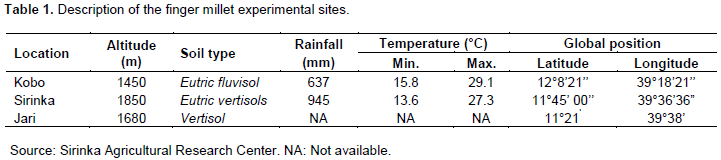
Eight finger millet varieties were included in the study and a local (farmers’) variety was also added for comparison. The eight varieties were released in Ethiopia by three different agricultural research centers (ARCs). Varieties Necho and Degu were released by Adet ARC in 2011 and 2005, respectively; Bareda and Gute in 2009, Wama and Boneya in 2007 and 2002, respectively, by Bako ARC; Padet and Tadesse in 1999 by Melkassa ARC. The experiment was laid out in a randomized complete block (RCB) design with three replicates. A plot size of 5 rows × 40 cm × 5 m was used, and a space of 1 m between plots and 0.5 m between blocks was also left. Seeding rate was 20 kg/ha. Nitrogen and
phosphorus fertilizers, as Urea and DAP, were applied at the rate of 50 kg/ha urea and 100 kg/ha DAP. Weeding was applied as the case may be, uniformly to all the plots. Data were collected both on plant and plot basis for phenological, yield and agronomic traits.
In 2013, data were recorded from the three locations. In 2014, however, data were recorded from Jari and Sirinka only, as the experiment at Kobo was failed due to deficit moisture stress and grass hopper attack. Additive Main-effect and Multiplicative Interaction (AMMI) analysis was done based on Zobel et al. (1988); genotype and genotype by environment interaction (GGE) analysis was done as per Yan et al. (2000). Analysis of variance, AMMI and GGE analyses were carried out by suing GenStat (16th edition) software.
RESULTS AND DISCUSSION
The results of the combined analyses of variance across locations for 2013 and 2014 are depicted on Tables 2 and 3. The year 2013 was relatively better than 2014 for finger millet yield, as evidenced by higher yield in 2014. In 2013, the location effect was significant (P≤0.01) for all the traits considered. However, genotypes (varieties) and G × L interaction term differed significantly for days to heading and maturity, and grain yield. In that same year, varieties Degu and Necho were late to flower and to mature; however, the rest were comparatively early to flower and to mature. Bareda, the local variety and Tadesse were the high-yielding varieties (Table 2).
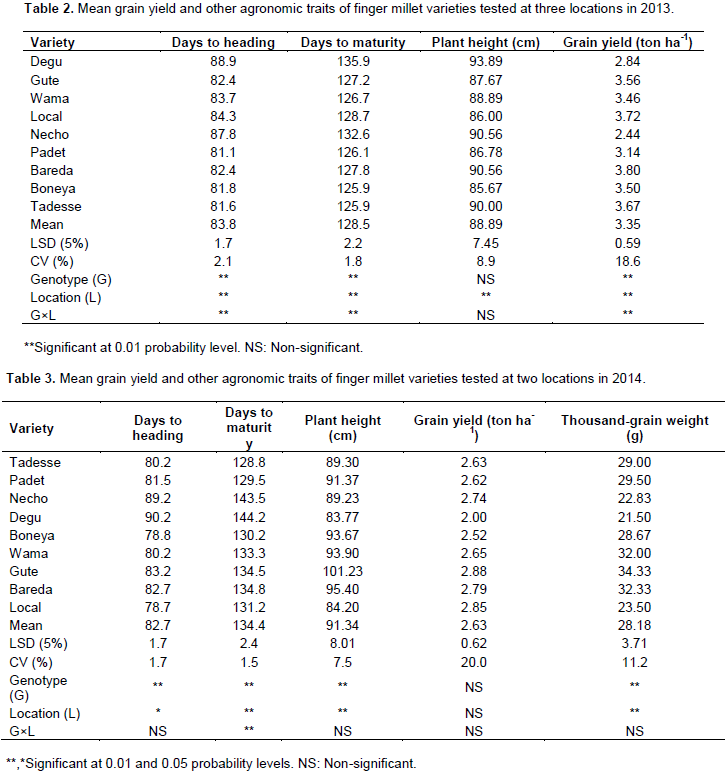
In the year 2014, varieties differed significantly (P≤0.01) for days to heading and maturity, plant height and 1000-grain weight. Similarly, the location effect was significantly different for all the traits considered except grain-yield. The G × L interaction term was significant only for days to maturity. Like the year 2013, Degu and Necho were late to flower and to mature in 2014 also. Gute was the tallest variety. As evidenced by higher 1000-seed weight, varieties Gute, Bareda and Wama were bold-seeded varieties as compared to the others (Table 3).
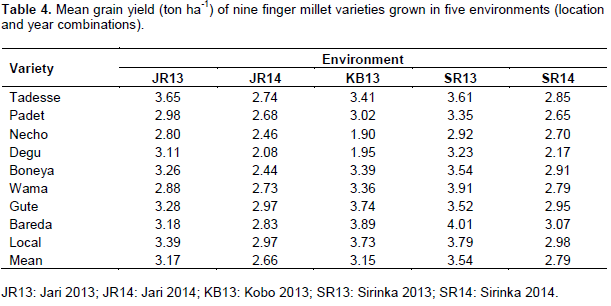
Location and year combinations were considered as environments shown in Table 4. Bareda was the highest yielder in the high-yielding environment (SR13). Similarly, Gute and the local check were the highest yielders in the low-yielding environment (JR14). Bareda ranked first at Sirinka and Kobo (SR13, SR14 and KB13) in terms of yield. Tadesse ranked first at JR13; however, both the local check and Gute ranked first at JR14. Except JR14, the local check ranked second in all the environments (Table 4).
The AMMI model combines AOV with additive parameters and PCA with multiplicative parameters into a single analysis (Gauch and Zobel, 1997). The AMMI and GGE analyses were based on five environments. The AMMI analysis demonstrated that genotype, environment, and G × E were significant (P≤0.01). Genotype, environment, and G × E explained 23.16, 36.71 and 40.12% of the total treatment variation (Table 5). Fentie et al. (2013) demonstrated 59.64, 9.61 and 30.75% of the total variation in finger millet grain yield to be accounted for by environment, genotype and G × E, respectively. In a mixed and colored sets of finger millet types, Adugna et al. (2011) reported 2.5, 79.1 and 18.3% of the total sum of squares in the mixed set and 2.1, 86.9 and 11.0% in the colored set to be attributed to genotype, environment, and G × E interaction effects, respectively. In a study of 15 finger millet genotypes under three environmental conditions, Misra et al. (2009) explained 7.88, 79.69 and 13.43% of the treatment variation in yield by genotype, environment and G × E, respectively. From the sources of variation of treatments, the genotype and interaction sources affect genotype rankings within environment and hence are relevant for targeting genotypes; these relevant effects account for 10 to 40% of the treatment variation (Gauch and Zobel, 1997). In the study, the treatment sum of squares (SS) was 57.12. However, the relevant portion of this SS was only 36.15 (genotype SS + interaction SS) or 63.29% of the treatment SS (Table 5).
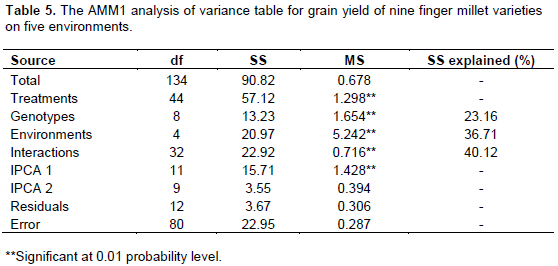
According to Gauch and Zobel (1997), in addition to the environmental effect, errors from uncontrolled variation within experiments are also irrelevant and should be removed from the interaction. Accordingly, from the interaction SS, 9.184 (the product of 32 and 0.287) was the real structure SS. As a result, the relevant variation was only 26.97 (13.23+13.74) or 47.21 % of the treatment SS, the rest would be irrelevant environment effect and interaction noise.
The AMMI analysis showed that the best fit model was AMMI1 in this experiment, as only the first interaction principal component axis (IPCA1) was significant (p<0.01). The IPCA1 explained 68.54% of the G × E interaction SS (Table 4). In agreement to the present investigation, Misra et al. (2009) demonstrated that the significant IPCA1 accounted for 93% of the total G × E interaction sum of squares. However, Wolie (2013) reported 61.22% of the G × E to be explained by the first two significant IPCAs. In mixed and colored finger millet sets of experiments, Adugna et al. (2011) explained 54.6 and 46.19% of the G×E sum of squares in the mixed and in the colored set, respectively, by the first two IPCAs. Similarly, Lule et al. (2014) explained 88.32% of the total G × E interaction in 32 finger millet genotypes by IPCA1 (66.05%) followed by IPCA2 (12.81%) and IPCA-3 (9.46%).
In the biplot system, main effects (both genotypes and environments) and IPCA1 (interaction) are used as abscissa and ordinates (Zobel et al., 1988; Gauch, 1992). The AMMI biplot showing the main and IPCA1 effects of both genotypes and environments on finger millet grain yield is as shown in Figure 1. As shown in Figure 1, distances along the abscissa show main effect differences, whereas the ordinate shows differences in interaction. In the present study, the AMMI1 biplot showed that genotypes Bareda, Local check and Gute had higher grain yield in that order; however, genotypes Padet, Necho and Degu had below average grain yield. The IPCA scores of genotypes in the AMMI analysis indicate the stability or adaptation over environments (Gauch and Zobel, 1996).
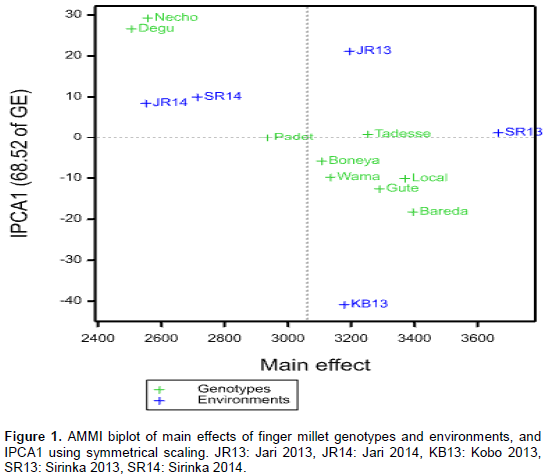
Similarly, environments SR13, JR13 and KB13 had above average grain yield; whereas, SR14 and JR14 had below average grain yield. Varieties Tadesse and Padet had small interaction effect; however, Bareda and Gute had relatively higher interaction effect. Likewise, SR13 contributed minimum interaction effect, whereas, KB13 and JR13 contributed higher interaction effect (Table 6 and Figure 1).
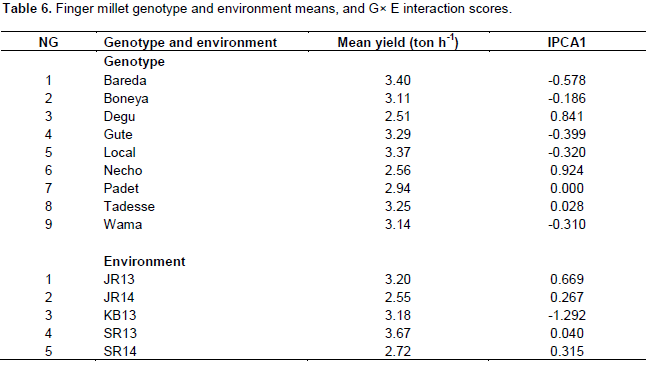
The GGE biplot is useful for identification of mege-environments, ideal genotype and test environments, among other things. Figure 2 shows the average-environment coordination (AEC) view of ranking genotypes relative to an ideal genotype. The center of the concentric circles pinpoints the ideal genotype, a genotype absolutely stable and the highest yielder. Thus, genotypes situated closer to the ideal genotype are more desirable (Yan and Tinker, 2006). Accordingly, the local check, Bareda and Gute are more desirable varieties than the others.
Necho has the only advantage of being white as farmers in some areas prefer white seeded varieties to brown seeded types for injera making (Assefa et al., 2009). Varieties Necho and Degu were neither stable nor high-yielding. In line with this, Vadivoo et al. (1998) reported that the yield potential of white grained types is generally lower than the brown ones. Besides, as these two varieties are late maturing, they could not fit in the moisture deficit areas like Kobo, Sirinka and Jari.
Based on the overall performance and adaptability of the nine finger millet verities over the five environments, as evidenced by AMMI and GGE analyses, the bold-seeded variety, Bareda could be recommended for production at Sirinka and Kobo, whereas the local check could still be used at Jari.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
This research work was financed by Amhara Regional Agricultural Research Institute.
REFERENCES
|
Adugna A, Tesso T, Degu E, Tadesse T, Merga F, Legesse W, Tirfessa A, Kidane H, Wole A, Daba C (2011). Genotype-by-environment interaction and yield stability analysis in Finger millet (Elucine coracana L. Gaertn) in Ethiopia. American Journal of Plant Sciences 2:408-415. |
|
|
Assefa A, Amare D, Tilahun D, Andargie D, Belay D, Asarigew F, Ayalew M, Wale M, Asfaw M, Altaye S, Amare T, Nigatie W (2009). Finger millet production in the Amhara region of Ethiopia. Research Report No. 1. ARARI, Bahir Dar, Ethiopia. |
|
|
Bano I, Gupta K, Singh A, Shahi NC, Khanchand, Gangwar V (2015). Finger millet: A potential source for production of gluten free beer. International Journal of Advanced Engineering Research and Applications 5(7):74-77. |
|
|
Bezaweletaw K, Sripichitt P, Wongyai W, Hongtrakul V (2006). Genetic variation, heritability and path-analysis in Ethiopian finger millet [Eleusine coracana (L.) Gaertn] landraces. Kasetsart Journal - Natural Science 40:322-334. |
|
|
David BM, Michael A, Doyinsola O, Patrick I, Abayomi O (2014). Proximate composition, mineral and phytochemical constituents of finger millet (Eleusine coracana). International Journal of Advances in Chemistry 2(2):171-174. |
|
|
Dida MM, Srinivasachary, Ramakrishnan S, Bennetzen JL, Gale MD, Devos KM (2007). The genetic map of finger millet, Eleusine coracana. Theoretical and Applied Genetics 114:321-332. |
|
|
Fentie M (2012). Participatory evaluation and selection of improved finger millet varieties in north western Ethiopia. International Research Journal of Plant Science 3(7):141-146. |
|
|
Fentie M, Assefa A, Belete K (2013). AMMI analysis of yield performance and stability of finger millet genotypes across different environments. World Journal of Agricultural Sciences 9(3):231-237. |
|
|
Fernandez DR, Vanderjagt DJ, Millson M, Huang Y-S, Chuang L-T, Pastuszyn A, Glew RH (2003). Fatty acid, amino acid and trace mineral composition of Eleusine coracana (Pwana) seeds from northern Nigeria. Plant Foods for Human Nutrition 58:1-10. |
|
|
Food and Agriculture Organization (2014). FAOSTAT. [Available from: http: //www.faostat. fao. org/site/567/default. aspx]. [Sourced: 23 November 2016]. |
|
|
Gauch HG (1992). Statistical analysis of regional yield trials: AMMI analysis of factorial designs. Elsevier, Amsterdam, 278 p. |
|
|
Gauch HG, Zobel RW (1996). Additive Main Effect and Multiplicative Interaction analysis of yield trials. In: Kang MS and Gauch HG (Eds.), Genotype by environment interaction. CRC Press, Boca Raton, FL. pp. 85-122 |
|
|
Gauch HG, Zobel RW (1997). Identifying mega-environments and targeting genotypes. Crop Science 37:311-326. |
|
|
Gowda PH, Prasad PVV, Angadi SV, Rangappa UM, Wagle P (2015). Finger millet: An alternative crop for the southern high plains. American Journal of Plant Sciences 6:2686-2691. |
|
|
Lule D, Fetene M, de Villiers S, Tesfaye K (2014). Additive Main Effects and Multiplicative Interactions (AMMI) and genotype by environment interaction (GGE) biplot analyses aid selection of high yielding and adapted finger millet varieties. Journal of Applied Biosciences 76:6291-6303. |
|
|
Lule D, Tesfaye K, Fente M, de Villiers S (2012). Multivariate analysis for quantitative traits in finger millet (Eleusine coracana subsp. coracana) population collected from eastern and southeastern Africa: Detection for patterns of genetic diversity. International Journal of Agricultural Research 7(6):303-314. |
|
|
Misra RC, Das S, Patnaik MC (2009). AMMI model analysis of stability and adaptability of late duration finger millet (Eleusine coracana) Genotypes. World Applied Sciences Journal 6(12):1650-1654. |
|
|
MoANR (2016). Crop variety register. Issue No. 18. Plant variety release, protection and seed quality control directorate. Addis Ababa, Ethiopia. |
|
|
National Academies (1996). Finger millet. P 39-58 In: Lost Crops of Africa: Volume 1. Board on Science and Technology for International Development, Office of International Affairs, National Research Council 408 p. |
|
|
Shayo NB, Tiisekwa BPM, Laswai HS, Kimaro JR (2001). Malting characteristics of Tanzanian finger millet varieties. Lishe Journal 10(1):1-4. |
|
|
Upadhyaya HD, Gowda C, Pundir R, Reddy VG, Singh S (2006). Development of core subset of finger millet germplasm using geographical origin and data on 14 quantitative traits. Genetic Resources and Crop Evolution 53:679-685. |
|
|
Vadivoo AS, Joseph R, Ganesan NM (1998). Genetic variability and diversity for protein and calcium contents in finger millet (Eleusine coracana (L.) Gaertn) in relation to grain color. Plant Foods for Human Nutrition 52:353-364. |
|
|
Woldemichael H, Admasu S (2017). Effects of processing on phytonutrient and nutritional composition of finger millet (Eleusine coracana): The neglected crop of Africa. Ethiopian Journal of Crop Science 5(1):17-36. |
|
|
Wolie A (2013). Performance of elite finger millet (Eleusine coracana) genotypes in West Amhara, In: Amhara Region Agricultural Research Institute (ARARI). Proceedings of the 5th Annual Regional Conference on Completed Research Activities of 2010 and 2011. ARARI. Bahir Dar, Ethiopia pp. 182-192. |
|
|
Yan W, Tinker NA (2006). Biplot analysis of multi-environment trial data: Principles and applications. Canadian Journal of Plant Science 86:623-645. |
|
|
Yan W, Hunt LA, Sheng Q, Szlavnics Z (2000). Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Science 40:597-605. |
|
|
Zobel RW, Wright MJ, Gauch HG (1988). Statistical analysis of a yield trial. Agronomy Journal 80:388-393. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


