Biotic inducing agents such as Trichoderma spp. are an alternative for the induction of resistance by activating defense mechanisms of plants. This work aimed to study the interaction between bean and Trichoderma spp. evaluating the effects of the induction of resistance in bean through the defense enzymes activity and ability to endofitism. The experimental was design in randomized blocks, with four replications, being 21 isolates of Trichoderma spp. and control. The isolates were inoculated at planting, with adding 0.8 ml of the suspension 1×108 conidia/ml in groove, in sterilized soil. After 30 days, leaf and roots samples were taken to determine the enzyme activity. To verify the endophytic colonization capacity of isolated, bean root fragments were sanitized and placed in Petri dishes containing PDA medium. Data were submitted to analyzes of variance and compared by Scott-Knott test (p<0.05). The enzymatic activity of peroxidase, polyphenoloxidase and phenylalanine ammonia-lyase was not significant, but the β-1-3-glucanase activity in leaf tissue was suppressed by isolates TI1, TM1, TLB15 (Trichoderma virens), TI2, TLB3, TLB4, TLB12, TOD1, TOD3 (Trichoderma harzianum), TLB6 (Trichoderma asperellun), TLB14, TLB17 (Trichoderma koningiopsis) and TOD2B (Trichoderma longibrachiatum). The isolates TM4, TLB9, TLB15 (T. virens), TI2, TI4, TLB2, TOD1 (T. harzianum) and TLB17 (T. koningiopsis), were able to colonize endophytically the bean roots. Trichoderma has endophytic capacity and interfere in β-1-3-glucanase activity.
The common bean (Phaseolus vulgaris L.) is exposed to factors that causes reduction on productivity and final quality of grain. Brazil is one of the major producer and consumer of this legume, with national production of 3,185,400,000 tons in the 2014/1015 harvest (Conab, 2015). However, there are several phytosanitary problems, that have great destructive potential, causing enormous losses in production of the country.
The Trichoderma is a fungus naturally found in different types of soils, living saprophyte and parasitizing other fungi, behaving as an antagonist to various phytopathogens, being the most searched fungus nowadays, once they are able to act as biological control agents, growth promoters and plant disease resistance inducers (Louzada et al., 2009).
Biological control is considered to be one of the method that may occur simultaneously throughout the antagonist life cycle, whose capacity can induce the synthesis of substances with antagonistic action by producing secondary metabolites which inhibit or stimulate the development of a microorganism or plant. In addition to biological control, there may be resistance induction, in which the plant actives defense mechanisms, signaled by elicitors of biotic and abiotic origin (Papavizas, 1985; Ridout et al., 1988).
The interaction between plant and fungus may have endophytic relations, establishing symbiotic relations still not well understood, but of great importance for plants (Azevedo, 1998). In interactions, the microorganisms produce or induce the production of secondary metabolites, which can provide many benefits, such as inducting resistance by the activation of defense mechanisms present at low levels in plant (Hammerschmidt and Dann, 1997; Schwan-Estrada et al., 2000).
Thus, the research aimed to study the interaction of different isolates of Trichoderma spp. in bean plants, by the effect on enzymes activity related to plant defense and endophytic capacity.
The experiment was conducted in laboratory and greenhouse. Isolates of Trichoderma spp., from native forest soils and of agricultural property in system of organic production, were isolated by the bait method (Ethur et al., 2005) and identified in species level by the sequencing region method internal transcriberd spacer (ITS) and part of gene elongation fator (EF) gene (Table 1).
To prepare the Trichoderma spp. inoculum, potato dextrose agar (PDA) discs having mycelium of each isolates, were transferred to erlenmeyers containing rice (20 g of rice to 25 ml of water and autoclaved for 20 min at 120°C), then, were incubated at 25 ± 2°C at photoperiod of 12 h, according to Silva et al. (2011) with adaptations.
After seven days, conidia suspension was prepared with 50 mL of sterile and distilled water, added to the Erlenmeyer to make rice washing. The suspension was filtered in gauze and the concentration was adjusted to 1×106 conidia mL-1, with the aid of a Neubauer chamber.
The experiment was conducted in plastic pots for up to three liters, which contained soil, sand and organic compound in a proportion of 3: 2: 1, autoclaved for 25 min at 120°C, repeating the operation in 24 h.
Seed were treated by the suspension of Trichoderma spp. isolates at planting, by application of 0.8 ml of the adjusted suspension (Broetto, 2013). In each pot were seeded two bean seeds of the cultivar IAPAR 81 (Iapar, 2015).
The experimental design was in randomized blocks, being 22 treatments with four replications (21 Trichoderma spp isolates and a control (without Trichoderma spp. inoculation)), deployed in 88 experimental plots.
For carrying out the biochemical analysis, 30 days after the emergence of bean the samples were collected samples from leaf and root tissue, wich were weighed and packed in aluminum envelopes, frozen and kept in a freezer at -20°C until the moment of analysis.
Samples from leaves and roots (0.5 g) were macerated mechanically in 4 mL of sodium acetate buffer 100 mM (pH 5.0), being added 0.04 g of polyvinylpyrrolidone. The mixture was centrifuged at 20.000 g for 25 minutes at 4°C. The supernate was taken as the enzyme extract and frozen at -20°C for determination of peroxidase (POD), polyphenoloxidase (PPO), phenylalanine ammonia-lyase (PAL) and β-1,3-glucanase (β-GAZE) activities (Lusso and Pascholati, 1999).
The POD activity was determined by direct spectrophotometric method, by the measurement of the conversion of guaiacol in tetraguaiacol to 30ºC, with spectrophotometric reading at 470 nm, for a period of 2 minutes, with optical density recorded every 15 s (Hammerschmidt et al., 1982). The reaction mixture contained 100 µL of the enzyme preparation and 900 µL of solution (12.5 mL of guaiacol and 310 µL of hydrogen peroxide in 87.5 ml of sodium phosphate buffer 0.01 M (pH 6.8)). The activity was expressed by the variation between the values in the linear increase range (E.U. (enzyme unit) min-1 g-1 fresh weight).
The PPO activity was determined using the methodology of Duangmal and Apenten (1999), adapted by Kuhn et al. (2010). The assay measured the catechol oxidation converted into quinone, reaction measured by the enzyme PPO. The substrate was composed by catechol in a concentration of 0.002 M, dissolved in sodium phosphate buffer 0.01 M (pH 6.8). The reaction was developed by mixing 900 µL of substrate and 100 µL of enzymatic preparation, with readings in spectrophotometer at 420 nm, for a a period of one minute, with optical density recorded every 15 s. The activity was determined by the variation between the values in the linear increase range E.U. (enzyme unit) min-1 g-1 fresh weight.
The PAL activity was determined by quantitation of trans-cinnamic acid released from the substrate phenylalanine; the methodology is described by Umesha (2006), in which 50 µL of the enzyme preparation were added to 450 µL of Tris-HCl 0.025 M buffer (pH 8,8) and 500 µL of a solution L-phenylalanine 0.05 M, incubated at 40°C for 2 h. The reaction was stopped by adding 60 µL of HCl 5 M, with read in a spectrophotometer at 290 nm. The PAL activity constitutes of the difference between the absorbance of the mixture and of the control (100 µL of protein extract and 900 µL of Tris-HCl 0.025 M buffer (pH 8.8)), which was guinea fowl in the standard curve to trans-cinnamic acid (y = 0.0095x + 0.0255, where y is the absorbance at 290 nm and x is the concentration of trans-cinnamic acid (µg)), expressed in µg trans-cinnamic acid h-1 g-1 fresh weight.
The β-GAZE activity was determined by colorimetric measurement of reducing sugars released from the laminarin (Vogelsang and Barz, 1993). The reaction involves 100 µL of the enzyme preparation, 50 μL of sodium phosphate buffer 0.01 M (pH 6.0) and 150 μL of laminarininarina (2 mg mL-1), which was developed at 40 ° C for one hour. In control, laminarin was added after the incubation. The sugars formed were quantified by Lever Method (1972), which consists of extracting an aliquot part of 30 μL of the previous solution and adding to this 1.5 mL of hydrazine solution of the p-hydroxybenzoic acid 0.5% (PAHBAH). The mixture was maintained at 100°C for 10 min, cooled on ice for 10 min, and the absorbance determined at 410 nm. The β-1,3-glucanase activity was the difference between the absorbance of the mixture containing the sample and control, which was painted in standard for glucose curve (y = 0.002x + 0.0046, where y is the absorbance and x the concentration of reducing sugars (μg)) and expressed as glucose equivalent μg-1 mL-1 h-1 g-1 fresh weight.
After collecting the roots samples for biochemical analysis, a portion of 10 cm of root from each plant was washed to evaluate the ability of Trichoderma spp. isolates, about its endophytic colonization capacity. This was performed by the indirect method of isolation, using root fragments of 1.5 cm, superficially disinfected with 70% alcohol for one minute, immersed in 3% sodium hypochlorite for one minute and washed three times in distilled autoclaved water. Then, three fragments were transferred into Petri plate, containing PDA médium, in three repetitions. The petri dishes were incubated in BOD at 25 ± 2 ° ° C with a photoperiod of 12 hours. The evaluation was made by viewing the Trichoderma spp. growth, from the root.
Once tabulated, the data were submitted to analysis of variance, and the averages compared by Scott-Knott, at 5% probability, with the assistance of statistical software version 5.3 Sisvar (Ferreira, 2010).
Despite the importance of Trichoderma sp. and research for resistance induction, just a little is understood about the relationship of microorganism-plant interaction of this fungus in the bean, by the fact of the paucity of studies involving the mechanisms by which Trichoderma spp. interacts, activating the biochemical defense systems of bean plants. Several hypotheses are presented, but few of them have been cogent, which are evaluated by chemical and cytological searches from plant tissues presenting these fungal agents (Hanson and Howell, 2004).
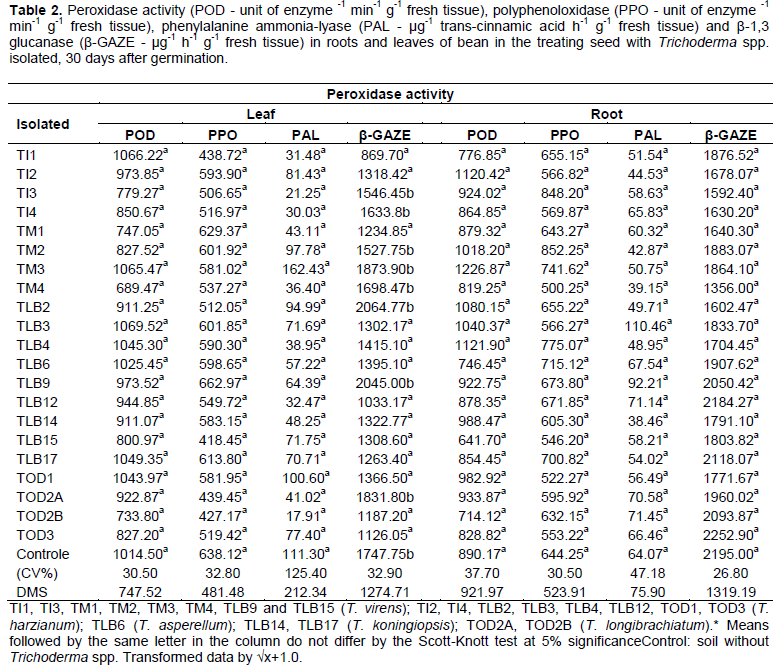
According to Table 2, the results of the enzymatic activity of POD, PPO and PAL in the leaves and roots showed no statistical difference between isolated of Trichoderma spp., but, the β-GAZE was reduced depending on the isolated, presenting statistical difference in bean leaves.
The POD activity in bean, even with no statistical difference between the isolated, can observe a tendency of suppression by the isolated TM1, TM4, TLB15 (T. virens), TOD2A (T. longibrachiatum) and TOD3 (T. harzianum) on the expression of this enzyme activity in leaf tissue of bean, as compared to control.
Similarly, the isolated TI1, TLB15 (T. virens), TOD2B (T. longibrachiatum) and TLB6 (T. asperellum), suppressed the enzymatic activity in root tissue. However, the isolated TM3 (T. virens) and TI2, TLB4 (T. harzianum), showed a tendency to increase the POD enzyme in bean plants, compared to the control.
Similar results for POD were obtained by Silva et al. (2011), that observed the effect of 60 isolated of Trichoderma spp. on growth promoting and systemic resistance induction to anthracnose in cucumber plants. They found no significant difference regarding the activation of the PPO enzyme in the plant between the 7th and 14th day after treated with Trichoderma spp., as compared to control.
However, Yedidia et al. (1999), working with hydroponic cucumber plants inoculated with Trichoderma, observed an increase in the POD enzyme after the time interval until seven days. However there was no increase in activity at 7 and 14 days after treatment, while the untreated plants showed a gradual increase over time, it was observed increase in plant growth, being confirmed, for the first time, the direct impact of this fungus in the interaction plant-fungus.
According to Yedidia et al. (1999), the increase in POD enzyme in leaves observed in their study, may be dued to either a systemic defense response, as the presence of Trichoderma in the rhizosphere, probably decrease of the enzymatic activity could be temporarily related to the root colonization by different isolated of Trichoderma spp.
A similar result was obtained by Dildey et al. (2013), wich have observed no POD and PPO activity induction in bean plants against the pathogen Macrophomina phaseolina, in inoculation of seeds with different isolates of T. harzianum. However, even it does not demonstrate significant results for enzymeshere, there was a tendency of such, on one of the different isolated of Trichoderma compared with the treatent control. From the results, the POD activity is not necessarily involved in this process, but further defense mechanisms such as chitinases, β-GAZE among others, may be acting in the induction of resistance of bean plants.
For the PPO activity, the results of this research showed no statistical difference, however, the isolated TI1, TI3, TLB15, TM4 (T. virens), TOD2A, TOD2B (T. longibrachiatum) and TI4, TLB2 (T. harzianum), had a suppression tendency of PPO enzymatic activity, compared with the control, for the leaf tissue.
For the PPO activity in the root tissues of bean plants, they have shown an increasing trend for the isolated TI3, TM2, TM3 (T. virens) and TLB4 (T. harzianum), however it can be seen that the isolated TLB15, TM4 (T. virens) and TI2, TI4, TLB3, TOD1, TOD3 (T. harzianum), suppressed this enzyme activity.
Pereira et al. (2008), working with chitosan extracted from Trichoderma spp. and Rhizopus sp., obtained a protective effect on cocoa plants, by noting that the application of biotic inducers are able to promote increased in POD and PPO enzymes activity, in addition to reducing Verticillium-wilt.
According to Pinol and Palazón (1996), in some cases, the polyphenols metabolism may act as antioxidant, leading to the inactivation of fenolases enzyme production. In another aspect, the biotic biosynthesis of secondary metabolites is attached the plant genetic constitution determining the activation of genes that synthesize the enzymes.
The PAL activity, showed no significant difference in bean plants. But, for leaf tissue, it can be noticed a sharp increase in the enzymatic activity of isolated TM3 (T. virens) compared to the control. On the other hand, all isolated present a suppression tendency for this enzyme, being this tred more evident in isolated TI1, TI3, TM4 (T. virens), TI4, TLB4, TLB12 (T. harzianum) and TOD2B (T. longibrachiatum), with no statistical difference
In expression of this enzyme in the root system, it is observed a increase tendency for isolated TLB3 (T. harzianum) and TLB9 (T. virens), however, the isolated TI1, TM2, TM3, TM4 (T. virens), TI2, TLB2, TLB4 (T. harzianum), TLB6 (T. asperellum) and TLB14 (T. koningiopsis), showed suppression in PAL enzymatic activity and no statistical difference.
Karthikeyan et al. (2006), evaluated the induction of phenol compounds, the defense enzymes involved in phenylpropanoid pathway and the protein-PR accumulation against Ganoderma in reply to the application of Pseudomonas fluorescens, T. viride and T. harzianum combined with chitin. The authors observed a high increase in PAL activity in all treated palms following application of P. fluorescens, T. viride and chitin. The highest level was reached 3 days after treatment and, then, declined slowly.
According to De Meyer et al. (1999) the rhizosphere colonization by P. aeruginosa, produced PAL activity in bean roots and, consequently, increased the salicylic acid level in the leaves.
As stated by Campos et al. (2003), a complex response wich involves production of phytoalexins and isoflavonoids may make a different role in the related response for the enzyme PAL induction. In addition of there being a complex response, involving phytoalexins and isoflavonoids production, it may play a different role in the response related to induction of that enzyme.
On the other hand, Nicholson and Hammerschmidt (1992), reported that the increase in PAL activity might be directly involved with a biochemical marker for induced resistance, once this enzyme is considered essential for the phenols synthesis, being important for resistance, enabling occurrence of a specific interaction between plant and inducer microorganism.
For β-GAZE activity, there was significant difference for the results of leaf tissue treated with the isolated TI1, TM1, TLB15 (T. virens), TI2, TLB3, TLB4, TLB12, TOD1 and TOD3 (T. harzianum). The isolated TLB6 (T. asperellun), TLB14, TLB17 (T. koningiopsis), TOD2B (T. longibrachiatum) suppressed the this enzyme activity. In tissue root system there was no difference, but was observed a more evident suppression trend in the isolated TM4 (T. virens).
According to Saksirirat et al. (2009), by working with fifteen isolated of Trichoderma spp. to control Xanthomonas campestres pv. vesicatoria there was a decrease of 69,32% of the symptoms, showed that four isolated of species T. harzianum e T. asperellum, induced activity of β-GAZE and chitinase in the leaves, showing that these species are promising for resistance induction in tomato plants. The same autors, They report that these enzymes are known to play an important role in the degradation of fungus cell wall. Besides that, are related to pathogenesis-related proteins (PR proteins) in several plants, including beans (Van Loon and Van Stien, 1999).
As stated by Salzman et al. (1998), these induced proteins are not always present during the infection process, they can be found in healthy tissue of certain plant part, as leaves or roots, or at specific developmental stages.
According to Akrami et al. (2011), several factors may interfere in the efficiency of biotic inducing agents among them the abiotics, mainly temperature, humidity and soil pH, wich are considered crucial on inducer efficiency, ensuring a successful performance of Trichoderma spp. isolated.
Among the 21 isolates of Trichoderma spp. evaluated for the colonization of bean plants roots, it was verified that 8 isolated feature the endophytism capacity in roots (Table 3), The results show that these isolates can be considered endophytic microorganisms
These data confirm those of Benítez et al. (2004), wich found that isolated of Trichoderma spp. are able of colonizing roots, producing secondary compounds that confer stimulate to plant growth by, as well as, induce resistance, by activation of plant defense mechanism. The same has been verified by Pedro (2012), that working with Trichoderma spp. isolated, in growth stimulating and anthracnose resistance induction in bean plants, showed that seven isolated of Trichoderma spp. evaluated were able to colonize the seedlings roots, being possible to check their growth on petri dishes, from the roots.
The gender Trichoderma may be associated with the roots of several plants of several species. Its penetration and colonization, associated with its antibiosis capacity, enhances an hydrolytic enzymatic arsenal, degrader of the plant cell wall. This input on epidermal layers of the roots confers a higher growth, improving nutrient absorption, and, thereby, increasing plant productivity (Harman et al., 2004).
According to the results of the present work, the reduction of enzymatic activity verified in bean plants inoculated with Trichoderma spp., is not very well clarified, it is believed that the Trichoderma spp., stimulates the plant to the equilibrium of its enzymatic arsenal, for the sake of its own benefit, so that, the plant will not endanger the symbiotic relationship between its interaction with the Trichoderma spp., because in case of the plant be stimulated to produce defense enzymes, Trichoderma, probably, would be hampered by the degradation of its structures and its cell death would be occurred.
Lucon (2009), verify that, in the absence of phytopathogens, the action of the mechanisms may be related to the production of hormones or growing factors, increase in the availability and absorption of nutrients or efficiency in the use of some nutrients by the plant.
These same Trichoderma spp. isolated, tested in this study, were used for Broetto (2013), which verified the effectiveness of the isolated TLB2, TLB3, TLB12 and TLB17 in bean against M. phaseolina. Also, stood out isolated TI1, TM3, TLB3, TLB12 and TLB17 as good growth promoters of the bean plants, whereas, the isolated TLB12 presented highlighted grain yield.
Several studies have been conducted to extend the knowledge of the action mechanisms by the biocontrol in diverse cultures. It is believed that, to the extent that the potential of these mechanisms will getting to know, in interactions with plants and microorganisms, lead to promising expectations for use in agriculture, as well as contribute towards the environment. preservation.