Full Length Research Paper

ABSTRACT
The feeding value of mulberry leaves for rabbits was quantitatively evaluated based on a single-factor design with five levels in diets (0, 5, 10, 15 and 20%). Results showed that rabbits given mulberry at 20 and 15% had a relatively lower body weight gain, higher feed conversion ratio, and lower meat ether extract contents than that in the 0% group (P < 0.05). Increased activities of antioxidant enzymes and reduced formation of thiobarbituric acid-reactive substances were detected in the plasma of mulberry-treated rabbits. Mulberry reduced the production of trichloroacetic acid perceptible N and NH3-N and increased total volatile fatty acids in rabbit cecum content through optimizing the intestinal micro-flora. Comparative analysis revealed that the content of phytochemicals in mulberry may be the main factor responsible for the feeding levels in rabbit diet, contributes to the effect of enhancing the antioxidant capacity of rabbit bodies and also optimizes intestinal micro-flora.
Key words: Animal nutrition, feed, rabbit, growth, health, blood, intestinal micro-flora.
INTRODUCTION
The use of alternative feed ingredients is an efficient means of reducing feed costs in animal production, and has attracted considerable attention among animal nutrition researchers in developing countries (Klinger, 2017). Mulberry (Morus spp.) species are widely distributed throughout Asia, Europe, Africa, and the Americas (Sánchez, 2002). The leaves of mulberries are highly palatable and easily digested (70-90%) by herbivores and can also be fed to monogastrics. The crude protein content of mulberry leaves and young stems range from 15 to 28%, the values of which are similar to those of most legume forages and superior to most grasses. Accordingly, mulberry is believed to have considerable potential and utility value as a non-conventional animal feed (Sánchez, 2000; Zhou et al., 2014). Mulberry leaf is non-toxic and can even protect human brain from pesticide toxicity (Yang et al., 2014; Smith, 2017). The Food and Agriculture Organization (FAO) strongly recommends that farmers use mulberry leaves as an animal feed source to replace edible grains consumed in animal production (Uribe and Sanchez, 2001). The nutritive value of mulberry leaves as a protein source has previously been estimated in different animals (Premalatha et al., 2012; Islam et al., 2015; Yupakarn et al., 2015).
The utilization of mulberry leaves as a nutrient source for rabbits has been previously studied. At first, it was advised that mulberry leaves could be used as a single feed for adult rabbits (Deshmukh et al., 1993). However, subsequent substitution experiments indicated that the use of mulberry leaves in feed should be appropriately limited; otherwise, the food intake and live-weight gain of rabbits would decrease, resulting in the production of lean meat. Bamikole et al. (2005) indicated that half of the concentrate in diets could be replaced by mulberry leaves; whereas Prasad et al. (2003) recommended that the proportion used in rabbit diets should be ≤45%, and that 15% could support a good productive performance. Nonetheless, the suitable proportion of mulberry leaves in rabbit diets remains controversial. It has been reported that the nutritional value and digestibility of mulberry leaves are as good as those of lucerne leaves; however, lucerne leaves constitute 48% of the matter in rabbit feed, whereas mulberry leaves are limited to a lower percentage (Premalatha et al., 2012). Consideration of the differences in usage amount should not only be based on difference in nutrient contents, as phytochemicals in mulberry leaves that have anti-obesity and antidiabetic effects may contribute to their inferior application. Previous studies have indicated that mulberry leaf extracts (rich in phenolic acids, flavonoids, and alkaloids) can regulate glycolipid metabolism and remove excess neutral fat and cholesterol from blood, tissues, and organs in humans (Zhang et al., 2014; Chang et al., 2016; Wu et al., 2017). Thus, long-term use and large doses may not fatten animals, thereby resulting in low productive performance. Therefore, as a non-conventional feed ingredient, the nutritive value of mulberry leaves should be assessed not only with respect to animal productivity, but also with regards to its effects on the health of animals, as well as on fat metabolism and mobilization that are critical to the meat type of animals. Numerous phytochemicals have been isolated from mulberry leaves; however, till date the effects of mulberry leaf constituents in feedstuffs on animal production are yet to be analyzed.
In this context, the feeding value of mulberry leaves in rabbits was evaluated. Specifically, the appropriate amount of mulberry leaves that should be used in feedstuff through providing a range of levels in formulated diets was assessed, and the effects of phytochemicals (phenolic acids, flavonoids, and alkaloids) contained in mulberry leaf-supplemented diets on rabbit production, health status, and cecum fermentation were examined.
MATERIALS AND METHODS
The research reported here was approved by the Ethics Committee of Zhenjiang Jiangbin Hospital, and all study procedures were conducted in accordance with national ethics regulations (GB/T 35823-2018 and GB/T 35892-2018). The rabbits slaughtered in this experiment were treated humanely.
Animals, housing, and feeding
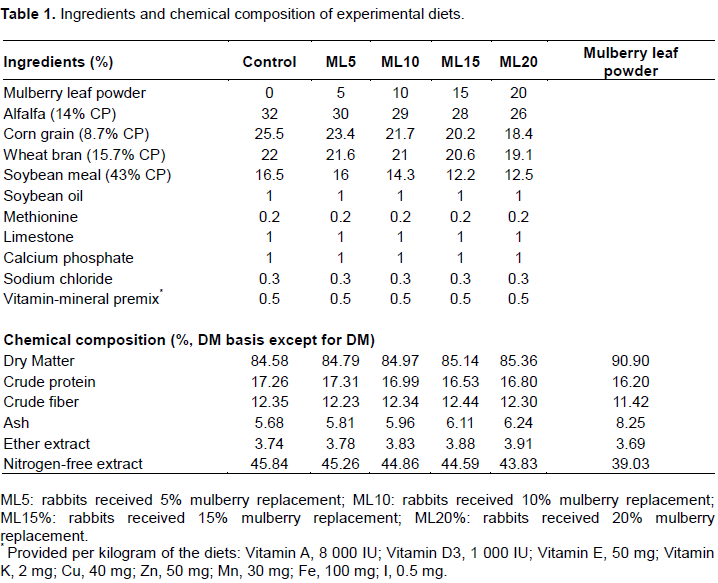
Eighty 2-months-old male New Zealand white rabbits with an initial body weight of 1.24 ± 0.20 kg were randomly distributed into five groups of eight animals each according to a single-factor design with five levels of mulberry leaf powder in the diet (0, 5, 10, 15, and 20%), which represented the following five treatments: control, ML5, ML10, ML15, and ML20. Eight replicates in each group and each replicate contain one animal. The mulberry leaf powder used was a commercial product processed from green leaves and new shoots, which were dried, ground, and passed through a 0.25-mm sieve.
The experiment was conducted for 42 days with a 14-day adaptive phase. During the trial period, animals were kept in galvanized wire batteries (45 × 45 × 40 cm) equipped with feeding hoppers and drinking nipples and maintained under the same managerial, hygienic, and environmental conditions in rooms with a natural temperature of 18–29°C, humidity of 43–70%, and photoperiod of 15L: 9D. All the rabbits were fed the respective diets allowing ad libitum intake and at least 10 g/day refusals. The chemical composition of the diets and mulberry leaf powder are shown in Table 1.
Samples collection and chemical analyses
Animals were weighed at the beginning of the trial and every week thereafter. Feed intake and uneaten feed were recorded weekly throughout the experimental period for determining growth performance. Daily weight gain and feed conversion ratio were calculated. At the end of the experiment, whole blood samples were collected from the marginal vein in the left ear of all rabbits and centrifuged at 3 000 × g for 10 min to collect plasma. Eight rabbits from each group were then deprived of feed for 12 h and slaughtered by jugular bleeding. Carcass parameters, including slaughter weight, dressing percentage, and organ percentage were determined according to Abu Hafsa et al. (2017); also, cecal contents were collected in a pre-warmed thermos and sampled for fermentation parameter and micro-flora population analyses. Subsequently, the right-side longissimus dorsi muscles were individually packed, weighed and frozen at -20°C until further analysis of chemical composition.
The chemical composition of mulberry leaf powder and meat samples, including dry matter, crude protein, crude fiber, ash, ether extract, nitrogen-free extract, and moisture content, were determined according to the Association of Official Analytical Chemists (AOAC, 2000). Total phenolic acids, flavonoids, and total alkaloids in mulberry leaf powder and diets were determined without fixation using the method described by Zhang et al. (2014).
Fasting glucose, cholesterol, low-density lipoprotein (LDL), high-density lipoprotein (HDL), triglyceride total protein, albumin and globulin in plasma were determined using a Pentra 400 analyzer (HORIBA ABX). The content of thiobarbituric acid-reactive substances (TBARS) and the antioxidant activities of glutathione peroxidase (GPx), glutathione S-transferase (GST), catalase (CAT), and superoxide dismutase (SOD) were determined using commercial kits obtained from Jiancheng Bio. Tech., Nanjing, China, according to the procedures outlined by the manufacturer. TBARS was measured using ELISA method and examined at 450 nm wavelength; GPx and CAT were measured at 470 and 240 nm wavelength, respectively with a spectrophotometer; GST was measured using fluorescence method and examined at 340 nm wavelength; SOD was measured using hydroxylamine method and examined at the 560 nm wavelength.
The pH of the cecal contents was determined by diluting 10 g of the contents with 15 ml distilled water and measured using a pH meter (PHSJ-4F; Shanghai Electric Science Instrument Company Limited). After centrifugation at 5 000 × g for 15 min, the supernatant was used for estimation of total N, trichloroacetic acid perceptible N (TCA-N), and total volatile fatty acids (VFAs) using a distillation method (Barnett and Reid, 1957). Aliquots of approximately 1 g of the cecal contents and 3 mL of a solution of 2% sulfuric acid or 2 mL of 2% ortho-phosphoric acid were mixed well for the analysis of ammonia nitrogen (NH3-N) and VFAs, respectively. The NH3-N concentration in the cecal contents was determined according to AOAC (2000). VFA analysis was conducted according to Alagón et al. (2014) using a gas chromatograph (Agilent 6890 GC) equipped with a 63 Ni electron capture detector (IECD: Agilent Technologies, Wilmington, DE, USA) and a 7694E automatic injector. Major bacterial species in the fresh cecal contents of rabbits were counted. In particular, aerobic and facultative anaerobic bacteria grown on tryptic glucose yeast agar, Escherichia coli on chromogenic coliform agar, Lactobacillus spp. on De Man-Rogosa-Sharpe agar, along with Clostridium spp., Bacteroides spp., and Bacillus spp. on PEA agar were determined and counted on plates using a Reichert Quebec® Darkfield Colony Counter according to Maturin and Peeler (2001).
Statistical analysis
Data were analyzed with one-way ANOVA using SPSS16.0 software (SPSS Inc., Chicago, IL, USA), followed by Duncan’s multiple range test. Results were expressed as means and the standard error of the means (SEM). Differences were considered significant at P ≤ 0.05. Dose-response curves were estimated using regression analysis available in SPSS (linear, quadratic, and cubic).
RESULTS
Chemical composition of diets
The chemical composition of the five levels of mulberry leaf inclusion diets were shown in Table 1. The nutritional composition of the dry matter was similar among the five treatments. In Table 2, the phenolic acid, flavonoid, and alkaloid contents of diets increased with an increase in the levels of supplemented mulberry leaves.
Growth performance, carcass, and meat quality analyses
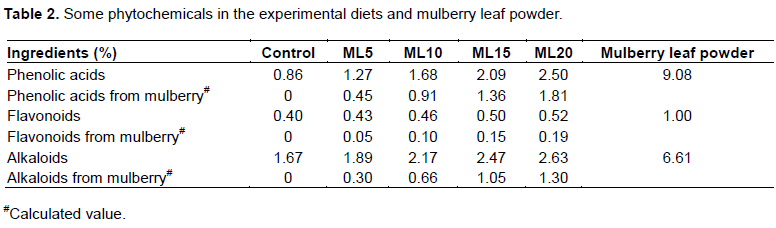
Rabbits in the ML10, ML15, and ML20 groups had a lower (P < 0.05) weight gain and feed intake than those in the control and ML5 groups (Table 3). The feed conversion ratio of the ML20 group was higher (P < 0.05) than that of the control and other mulberry groups; whereas there were no significant differences among the control, ML5, ML10, and ML15 groups. The slaughter weight of rabbits in the ML20 group was significantly lower (P < 0.05) compared with that in the control and other mulberry groups. Mulberry inclusion diets (≤20%) had no significant (P > 0.05) effect on the dressing percentage, weight percentage of organs (liver, kidney, and heart), meat moisture, or protein and ash contents, whereas the ether extract for ML15 and ML20 group rabbits decreased significantly (P < 0.05) compared with that of the other treatments.
Biochemical analysis of plasma
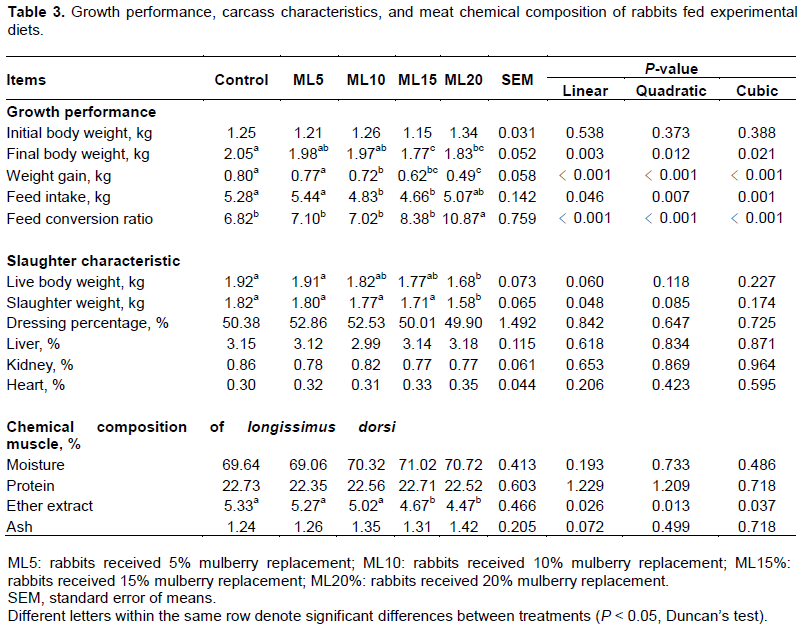
The results presented in Table 4 indicate that the plasma TBARS content in rabbits of the ML15 and ML20 groups was lower than that in rabbits in the control group, whereas the activities of SOD, GPx, GST, and CAT showed significant (P < 0.05) dose-dependent increases in rabbits treated with mulberry inclusion diets. However, there were no significant effects of mulberry leaf powder (P > 0.05) on the fasting glucose, cholesterol, HDL, LDL, triglycerides, protein, and albumin/globubin contents in the plasma of rabbits.
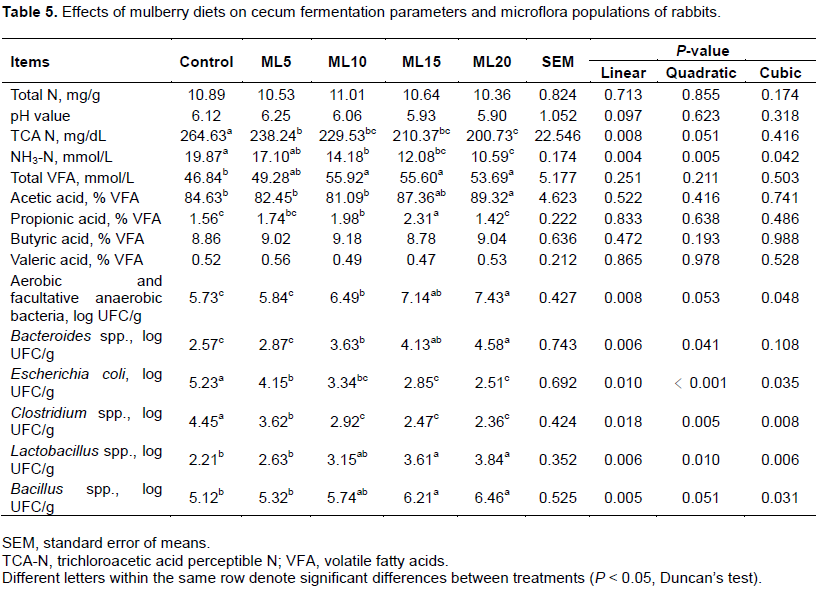
Cecal fermentation and micro-flora
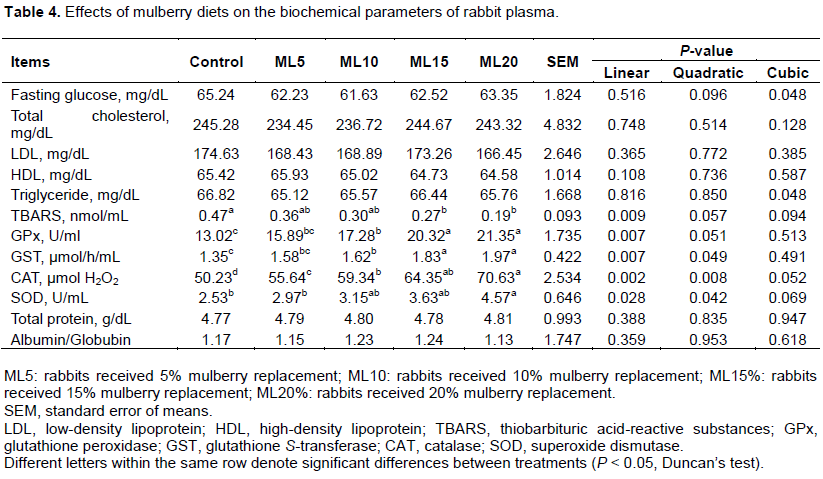
As indicated in Table 5, increasing levels of mulberry leaf powder in diets did not result in substantial changes in the total N or pH of cecal content. The rabbits fed on the mulberry inclusion diets did, however, show a marked reduction in the TCA-N level in cecum liquor at the end of the 42-day experimental period. A similar result was also obtained for the NH3-N level, which, as expected, decreased significantly (P < 0.01). In contrast, total VFA in the cecum liquor of rabbits in the ML10, ML15, and ML20 groups increased significantly (P < 0.05) compared with the control group. In particular, animals fed the ML20 diet showed a significantly (P < 0.05) higher acetic acid concentration compared with those fed low mulberry leaf powder diets (ML5 and ML10) and the control. An increasing proportion of mulberry leaf powder in diets did not result in a similar trend in prop ionic acid concentration in cecal contents, as the data indicated a sudden fall in the high mulberry diet group (ML20). However, butyric acid and valeric acid did not differ significantly between the control and mulberry inclusion diet groups. The data obtained from bacteriological analysis showed high variability, which tended to obscure differences among the experimental groups. However, the E. coli and Clostridium spp. counts were statistically lower in the mulberry leaf powder-treated groups than in the control group. Furthermore, an increase in the proportion of dietary mulberry leaf powder significantly (P < 0.05) increased Bacteroides spp., Lactobacillus spp., and Bacillus spp. in the high mulberry leaf powder inclusion groups (ML20 and/or ML15).
DISCUSSION
The selection of the percentages of mulberry leaves in rabbit diets (5, 10, 15 and 20%) used in the present study was based on reference to previous results. The data obtained indicated that the utilization of mulberry leaf in rabbit feed at levels of 5 and 10% in the ML5 and ML10 groups maintained growth performance normally during the entire experimental period, whereas levels of 15 and 20% in the ML15 and ML20 groups resulted in a significant reduction in growth. However, although the final weight and average daily gain of the rabbits were lower in rabbits fed the ML15 diet compared with those fed the control and ML5 diets, the feed conversion ratio and carcass yield were still satisfactory from a production point of view. Rabbits in mulberry treated groups were all healthy, which was indicated by the little variation in selected health markers (organ weight, plasma total protein, and albumin/globubin) between the control group and the mulberry treated groups (Huang et al., 2017), and through this it can be inferred that mulberry leaf is non-toxic, which is in accordance with Yang et al. (2014), who examined the toxicity of mulberry leaf ethanol extract and found no toxic effect on adipocytes. Hence, the level of mulberry leaf powder included in the diet of growing rabbits can be up to 15% without having an adverse effect. Prasad et al. (2003), reported that mulberry leaves can completely replace lucerne hay in the diet of growing rabbits, that was 15%, and that even higher levels (that is, 30 and 45% of the diet) could also promote growth of rabbits, . Bamikole et al. (2005), suggested that 50% of concentrate could be replaced by mulberry leaves. The study results do not recommend the dosage of more than 15% of mulberry, for the feed conversion ratio may rise.
The chemical compositions were similar among treatments, indicating energy supply did not differ substantially among the control and mulberry leaf inclusion diets, and thus are probably not the main cause of the observed differences in growth rates. The primary difference among the different levels of mulberry inclusion diets may have been the contents of phytochemicals. Mulberry leaf ethanol extracts contain polyphenols and alkaloids (Jeszka-Skowron et al., 2014). It has been reported that mulberry leaf ethanol extracts inhibit fat accumulation in adipocytes by altering the protein expression levels of adipogenesis-related factors (Yang et al., 2014). Mulberry leaf polyphenols have been shown to inhibit hepatic lipogenesis, promote lipolysis, and lower high-fat diet-induced body weight in hamsters (Peng et al., 2011). The anti-lipogenesis and lipolysis-enhancement effect of mulberry leaf phytochemicals may result in lean meat and less weight gain in non-obese animals fed with normal feed, which was confirmed by the results obtained for the high mulberry inclusion treatment ML20. To analyze the dose-response effect of the mulberry leaf phytochemicals in detail, the content of supplemented phenolic acids, flavonoids, and alkaloids in the ML15 diet were calculated (1.36, 0.15 and 1.05%, respectively), and in terms of animal weight ratio, the data were 1034, 114, and 798 mg/kg of body weight [calculated using the middle weight (1.46 kg) and feed intake (111 g/day) in the ML15 group], respectively. It has previously been reported that an intake of 25 mg/L mulberry leaf phenolic-rich water extract reduces fatty acid storage in Caenorhabditis elegans in vivo (Zheng et al., 2014), and that intake of 1.0% mulberry leaf polyphenols (phenolic acids + flavonoids) or 2 mg/kg of body weight of alkaloids from curry leaves per day results in markedly less weight gain in murines and provided a high-fat diet (Peng et al., 2011; Jagtap et al., 2017). Although the influence of phytochemicals from different sources varies in different animals, their efficacy may be the same. The dosages of phenolic acids, flavonoids, and alkaloids in the ML15 treatment of the present study were higher than those used in the above-mentioned studies, and in this regard, the anti-lipogenesis and lipolysis-enhancement effect of the ML15 diet may be stronger, indicating that the content of phytochemicals (phenolic acids, flavonoids, and alkaloids) in mulberry leaves may be the main factor responsible for the low growth of rabbits.
To determine the health status of the experimental rabbits, biochemical indices in plasma were examined. On the basis of the data presented in Table 3, there was no significant change in fasting glucose, cholesterol, HDL, LDL, or triglycerides of rabbits in the mulberry leaf treatment groups. Mulberry leaves are used to prevent postprandial hyperglycemia and hyperlipidemia in type-2 diabetes mellitus treatment to promote glycolipid metabolism (Andallu et al., 2001; Tsuduki et al., 2013). For disease-free animals, glycolipid metabolism improves, and the levels of glucose and lipid metabolites can return to the normal levels a short time after feeding. Mulberry leaf ethanol extract has the effect of accelerating the reduction rate of postprandial hyperglycemia and hyperlipidemia in normal rats (Miyahara et al., 2004), illustrating that mulberry leaves can maintain blood glucose concentrations at normal levels, which prevents excessive amounts of glucose from circulating in the blood or converts it into fat distributed throughout the body.
In the present study, plasma thiobarbituric acid-reactive substances (TBARS), which are considered early biomarkers of oxidative damage, were discovered in all rabbits groups, thereby indicating that the experimental animals were suffering from oxidative stress (Ghani et al., 2017). For animals, many factors, including environmental change, negative energy balance, consumption of oxidized diets, weaning, and infection, can lead to oxidative stress (Yin et al., 2013; Celi and Gabai, 2015). Mulberry leaves appear to have a significant inhibitory effect against the formation of TBARS, as the content of TBARS in rabbits fed on the ML15 and ML20 mulberry leaf diets were lower than in the control group animals, which in turn indicates that oxidative damage was reduced by consuming mulberry leaves. A reduction in oxidative damage was mediated through an increase in the activity of the antioxidant enzymes SOD, GPx, GST, and CAT in plasma (Table 3). These results are consistent with the findings of Cheong et al. (2012) obtained from the muscle of beef cattle fed by mulberry leaf silage. Mulberry leaf phytochemicals such as phenolic acids, flavonoids, and 1-deoxynojirimycin (DNJ, an important alkaloid in mulberry leaf) are strong antioxidants (Jeszka-Skowron et al., 2014; Pham et al., 2017). The increased activity of antioxidant enzymes in rabbits was stimulated by these antioxidants, which can be inferred from the study of Andallu and Nch (2003), in which it was demonstrated that the activities of antioxidant enzymes in uncontrolled diabetes were improved efficiently by mulberry leaf powder treatment in streptozotocin-induced diabetic rats.
In the present study, the use of mulberry leaves altered the concentrations of cecal metabolites in rabbits. The increase in total VFAs and the slight (non-significant) decrease in pH suggests that fermentation activity in the cecum was higher in the mulberry leaf-supplemented groups ML10, ML15, and ML20 (Table 3), which is consistent with the observations of Prasad et al. (2003), who reported a significant decrease in NH3-N in the cecum of rabbits fed with 15% and 20% mulberry leaves. Studies on finishing steers revealed similar NH3-N and VFA concentration changes in rumen fluid, although no differences were detected in pH values between mulberry leaf and non-mulberry leaf groups (Zhou et al., 2014). Different levels of NH3-N and VFAs could be related to changes in the composition of commensal cecal and intestinal micro-flora, as some of the microbes in the cecum originate from gastrointestinal tract. An effect of mulberry leaf on modulating intestinal micro-flora was also identified in the cure of intestinal flora disorder in streptozotocin-induced diabetic rats (Sheng et al., 2017).
In ewes, supplementation of mulberry flavonoids has been shown to improve the digestibility of organic matter and reduce CH4 output by inhibiting the populations of microbes involved in methanogenesis (Ma et al., 2016). As indicated by the findings of the present and previous studies, 10~20% mulberry leaves in the diet can effectively optimize the cecum micro-flora as well as the intestinal micro-flora of rabbits, and this capacity of mulberry leaf can be attributed to the constituent phytochemicals.
CONCLUSION
The results of this study indicate that inclusion of mulberry leaves in the diet was suitable for raising rabbits. Inclusion of ≤15% mulberry leaves in the diet could promote performance from a production point of view. Phytochemicals (phenolic acids, flavonoids, and alkaloids), which may be the main factor restricting the acceptable amount of mulberry leaves that can effectively be included in rabbit feed, also contribute to the enhancement of antioxidant activity in rabbit blood and optimization of the micro-flora population in the rabbit cecum. For the future research, how to reduce the restricting factors would be a significant direction, and that would be helpful to increase the usage of mulberry in animal feed.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This research was supported by the Construction of Modern Agricultural Industry Technology System of China (CARS-18-ZJ0207), the Overseas Study Program 2016 of Colleges and Universities in Jiangsu Province for Outstanding Young and Middle-aged Teachers, Jiangsu Science and Technology Project (BZ2016055), China-Cuba Sericulture Technology Cooperation Center Construction and International Technology Transfer Service,Zhenjiang Agricultural Technology support project (NY2018013) and Public Industry (Agriculture) Specific Research Program (201403049).
REFERENCES
|
Abu Hafsa SH, Ibrahim SA, Hassan AA (2017). Carob pods (Ceratonia siliqua L.) improve growth performance, antioxidant status and caecal characteristics in growing rabbits. Journal of animal physiology and animal nutrition 6:1307-1315. |
|
|
Alagón G, Arce ON, Martínez-Paredes E, Ródenas L, Cervera C, Pascual JJ (2014). Effect of inclusion of distillers dried grains and solubles from barley, wheat and corn in isonutritive diets on the performance and caecal environment of growing rabbits. World Rabbit Science 22:195-205. |
|
|
Andallu B, Nch V (2003). Antioxidant role of mulberry (Morus indica L. cv. Anantha) leaves in streptozotocin-diabetic rats. Clinica Chimica Acta 338:3-10. |
|
|
Andallu B, Suryakantham V, Lakshmi BS, Reddy GK (2001). Effect of mulberry (Morus indica L.) therapy on plasma and erythrocyte membrane lipids in patients with type 2 diabetes. Clinica Chimica Acta 314:47-53. |
|
|
AOAC (2000) Official methods of analysis of the Association of Official Analytical Chemists International, 17th revised edition. AOAC, Maryland, USA. |
|
|
Bamikole MA, Ikhatua MI, Ikhatua UJ, Ezenwa IV (2005). Nutritive value of mulberry (Morus Spp.) leaves in the growing rabbits in Nigeria. Pakistan Journal of Nutrition 4:231-236. |
|
|
Barnett AJG, Reid RL (1957). Studies on the production of volatile fatty acids from grass by rumen liquor in an artificial rumen: I. The volatile acid production from fresh grass. The Journal of Agricultural Science 48:315-321. |
|
|
Celi P, Gabai G (2015) Oxidant/Antioxidant Balance in Animal Nutrition and Health: The Role of Protein Oxidation. Frontiers in Veterinary Science 2:48. |
|
|
Chang YC, Yang MY, Chen SC, Wang CJ (2016). Mulberry leaf polyphenol extract improves obesity by inducing adipocyte apoptosis and inhibiting preadipocyte differentiation and hepatic lipogenesis. Journal of Functional Foods 21:249-262. |
|
|
Cheong SH, Kim KH, Jeon BT, Park PJ, Hwang IH, Choi NJ, Kim ET, Hong SK, Park JH, Sung SH, Thomas DG, Moon SH (2012). Effect of mulberry silage supplementation during late fattening stage of Hanwoo (Bos taurus coreanae) steer on antioxidative enzyme activity within the longissimus muscle. Animal Production Science 52:240-247. |
|
|
Deshmukh SV, Pathak NN, Takalikar DA, Digraskar SU (1993). Nutritional effect of mulberry (Morus alba) leaves as sole ration of adult rabbits. World Rabbit Science 1:67-69. |
|
|
Ghani MA, Barril C, Bedgood Jr DR, Prenzler PD (2017). Measurement of antioxidant activity with the thiobarbituric acid reactive substances assay. Food Chemistry 230:195-207. |
|
|
Huang Q, Gao X, Liu P, Lin H, Liu W, Liu G, Zhang J, Deng G, Cao H, Guo X, Hu G (2017). The relationship between liver-kidney impairment and viral load after nephropathogenic infectious bronchitis virus infection in embryonic chickens. Poultry Science 96:1589-1597 |
|
|
Islam MR, Siddiqui MN, Khatun A, Siddiky MNA, Rahman MZ, Bostami ABMR, Selim ASM (2015). Dietary effect of Mulberry leaf (Morus alba) meal on growth performance and serum cholesterol level of broiler chickens. Dissertations and Theses - Gradworks 12:79-89. |
|
|
Jagtap S, Khare P, Mangal P, Kondepudi KK, Bishnoi M, Bhutani KK (2017). Effect of mahanimbine, an alkaloid from curry leaves, on high-fat diet-induced adiposity, insulin resistance, and inflammatory alterations. Bio factors 43:220-231. |
|
|
Jeszka-Skowron M, Flaczyk E, Jeszka J, Krejpcio Z, Król E, Buchowski MS (2014). Mulberry leaf extract intake reduces hyperglycaemia in streptozotocin (STZ)-induced diabetic rats fed high-fat diet. Journal of Functional Foods 8:9-17. |
|
|
Klinger ACK, de Silva LP, de Toledo GSP, Galarreta B, Goulart F (2017). Effect of sweet potato vines on performance parameters and some carcass characteristics of rabbits. Journal of Animal Physiology and Animal Nutrition 102:152-156. |
|
|
Ma T, Chen DD, Tu Y, Zhang NF, Si BW, Diao QY (2016). Dietary supplementation with mulberry leaf flavonoids inhibits methanogenesis in sheep. Animal Science Journal 88:72-78. |
|
|
Maturin L, Peeler JT (2001). Aerobic plate count. Bacteriological analytical manual. Silver Spring, MD: U.S. Food and Drug Administration, Chapter 3. |
|
|
Miyahara C, Miyazawa M, Satoh S, Sakai A, Mizusaki S (2004). Inhibitory Effects of Mulberry Leaf Extract on Postprandial Hyperglycemia in Normal Rats. Journal of Nutritional Science and Vitaminology 50:161-164. |
|
|
Peng CH, Liu LK, Chuang CM, Chyau CC, Huang CN, Wang CJ (2011). Mulberry water extracts possess an anti-obesity effect and ability to inhibit hepatic lipogenesis and promote lipolysis. Journal of Agricultural and Food Chemistry 59:2663-2671. |
|
|
Pham PP, Morales NP, Pitaksuteepong T, Hemstapat W (2017) Antioxidant activity of mulberry stem extract: A potential used as supplement for oxidative stress-related diseases. Songklanakarin Journal of Science and Technology 39:407-414. |
|
|
Prasad R, Misra AK, Sankhyan SK, Mishra AS, Tripathi MK, Karim SA, Jakhmola RC (2003). Growth performance and caecal fermentation in growing rabbits fed on diets containing graded levels of mulberry (Morus alba) leaves. Asian-Australasian Journal of Animal Sciences 16:1309-1314. |
|
|
Premalatha N, Saravanakumar VR, Sivakumar K, Jagatheesan PNR, Ramesh V (2012). Growth and carcass characteristics of Soviet Chinchilla rabbits fed mulberry and hedge lucerne leaves. Indian Journal of Small Ruminants 18:85-89. |
|
|
Sánchez MD (2000). Mulberry: an exceptional forage available almost worldwide. FAO Electron. Conf. "Mulberry for Animal Production". FAO, Rome, Italy. World Animal Review 93:1. |
|
|
Sánchez MD (2002). World distribution and utilization ofmulberry and its potential for animal feeding. FAO Electronic Conference on Mulberry for Animal Production. FAO Animal Production and Health Paper 147:1-9. |
|
|
Sheng Y, Zheng S, Ma T, Zhang C, Ou X, He X, Xu W, Huang K (2017). Mulberry leaf alleviates streptozotocin-induced diabetic rats by attenuating NEFA signaling and modulating intestinal microflora. Scientific Reports 7:12041. |
|
|
Smith J (2017). Mulberry leaf protects the brain from glyphosate toxicity. Botanicals, Brain Health, Environmental Health. |
|
|
Tsuduki T, Kikuchi I, Kimura T, Nakagawa K, Miyazawa T (2013). Intake of mulberry 1-deoxynojirimycin prevents diet-induced obesity through increase in adiponectin in mice. Food Chemistry 139:16-23. |
|
|
Uribe TF, Sanchez MD (2001). Mulberry for animal production: animal production and health paper. Rome: FAO 33:199-202. |
|
|
Wu T, Qi X, Liu Y, Guo J, Zhu R, Chen W, Zheng X, Yu T (2017). Dietary supplementation with purified mulberry (Morus australis Poir) anthocyanins suppresses body weight gain in high-fat diet fed C57BL/6 mice. Food Chemistry 141:482-487. |
|
|
Yang SJ, Park NY, Lim Y (2014). Anti-adipogenic effect of mulberry leaf ethanol extract in 3T3-L1 adipocytes. Nutrition Research and Practice 8:613-617. |
|
|
Yin J, Wu MM, Xiao H, Ren WK, Duan JL, Yang G, Li TJ, Yin YL (2013). Development of an antioxidant system after early weaning in piglets. Journal of Animal Science 92:612-619. |
|
|
Yupakarn W, Pattarajinda V, Lowilai P, Priprem S (2015). Effects of using Indian mulberry leaves as feed additives on feed digestion, ruminal fermentation and milk production in dairy cattle. Pakistan Journal of Nutrition 14:620-624. |
|
|
Zhang XM, Ma TJ, Sun Y, Bi QL, Zhang JQ (2014). Determination of flavonoids, phenolic acids and alkaloids in different varieties of mulberry leaves. Advanced Materials Research 962-965:1248-1251. |
|
|
Zhou Z, Zhou, B, Ren L, Meng Q (2014). Effect of ensiled mulberry leaves and sun-dried mulberry fruit pomace on finishing steer growth performance, blood biochemical parameters, and carcass characteristics. PLoS One 9:e85406. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


