Full Length Research Paper

ABSTRACT
Medicago truncatula is an omni-Mediterranean species grown as an annual forage legume. In addition to its small genome size and simple genetics, M. truncatula harbors several attributes which make it an attractive model legume. In this study, we investigated morphological and photosynthetic responses to water deficit in parental genotypes of M. truncatula recombinant inbred lines. Ten parental lines were cultivated under three water regimes (100% of field capacity (FC), 50% FC and 30% FC) and were harvested at flowering time and at the end of their lifecycle. Results from ANOVA showed that variability of measured parameters was explained by the effects of line, treatment and their interaction with treatment factor recorded the highest values. Out of the 27 traits, 14 were influenced by the line x treatment interaction. High to moderate broad-sense heritability (H²) were observed for most of the traits under control treatment and drought stress. Most of the correlations between measured traits were positive under the three water regimes. The flowering time was positively correlated with aerial and root growth rate. The tolerance of lines to water deficit seems to depend, in particular, on their ability to maintain higher photosynthetic activity. In 30% FC, principal component analysis clustered lines into two groups. The Jemalong A17 line was the least affected for most phenotypic parameters. Hence, all populations of recombinant inbred lines derived from crosses between Jemalong A17 and the remaining lines were useful for the identification of the genetic determinants for tolerance to water deficit in M. truncatula.
Key words: M. truncatula, parental lines, recombinant inbred lines, water deficit, morphological traits, photosynthetic parameters.
INTRODUCTION
Environmental constraints are the main factors limiting agricultural productivity and play a major role in the distribution of plant species. Among the abiotic stresses, water deficit is considered the most important factor in limiting crop production (Harb et al., 2010). Some adaptive responses to water deficit correspond to changes in the structure of plants (Shao et al., 2008), the growth rate and gas exchange at the leaf level (Slama et al., 2007), and overproduction of compatible compounds (Nunes et al., 2008). One of the first responses to drought stress is stomatal closure (Anjum et al., 2011). As a result, photosynthetic assimilation is unavoidably reduced due to decreased CO2 concentrations at the chloroplast level (Cornic, 1994). Furthermore, a reduced leaf area and decreased transpiration rate is often the result of drought stress (Dong and Zhang, 2000). Consequently, gas exchange characteristics and chlorophyll concen-trations are reduced as stress intensity increases.
Legumes are very important sources of vegetable protein for human food and animal feed and these plants do not require nitrogen fertilization. Drought represents one of the main factors which limits legume productivity in the Mediterranean basin. A better understanding of the mechanisms for adaptation to water deficit in legumes is a prerequisite for any research aiming to improve legume yields. The slowness of genetic improvements in most legumes of economic interests is due to some unfavorable traits such as their large genome, their complex ploidy, and the difficulties of their transformation and regeneration. M. truncatula thus emerged as a model legume because of several unique characteristics (Barker et al., 1990). It is diploid (2n=16), self-pollinating, has a low DNA content per haploid genome (~465 Mbp), is suitable for cultivation in the laboratory (size, growth habit, flowering, etc.) and has a rapid lifecycle. Furthermore, several studies on M. truncatula have contributed to an expanded genetic and genomics tools (Tang et al., 2014). It is also an important forage crop species in several countries especially Australia.
In recent years, molecular mechanisms for tolerance to drought stress in plants have become an active area of investigation and many genes regulated by drought stress have been reported in a variety of plants (Sehgal et al., 2012; Thudi et al., 2014; Xu et al., 2014). Very little is known about the physiological and molecular mechanisms by which the model legume M. truncatula responds to drought stress. In most of the previous experiments plants were terminated at the vegetative stage (Badri et al., 2011; Wang, 2014; Zhang et al., 2014). Furthermore, these previous studies have used only one population of RILs to analyze the genetic determinants for tolerance to drought stress. Therefore, it is important to analyze the genetic bases of tolerance to this constraint by using more than one population of RILs in addition to studying their responses at both the vegetative and reproductive stages.
The objective of this study was to evaluate the effects of water deficit on photosynthetic and growth charac-teristics in parental genotypes of RILs of M. truncatula at flowering and end of lifecycle stages. This as well as similar investigations can lead to the selection of more than one appropriate RIL population useful for the analysis of genetic determinism to drought. Furthermore, knowledge obtained by studying M. truncatula can be transferred to other legumes.
MATERIALS AND METHODS
Plant material and experimental conditions
Ten parental lines of M. truncatula recombinant inbred lines (RILs) collected in different regions that varied in soil composition, salinity and water availability were used in this study. These lines were intercrossed in order to provide useful material for the analysis of genetic determinism of traits of agronomical interest (Figure 1). These lines are DZA315.16 and DZA45.5 from Algerian populations, Jemalong A17 from the Australian collection, F83005.5 line from the Var region in France, A20 line of Moroccan origin and Tunisian lines TN1.11, TN1.21, TN8.3, TN8.20 and TN6.18 collected respectively from Enfidha (TN1), Soliman (TN8) and Thala (TN6) sites. The nomenclature of local lines includes “TN” to signal their Tunisian origin, the first number to refer to their collection site, and the second number is added to indicate the number of the pod.
.png)
The experiment was performed under greenhouse conditions at the Centre of Biotechnology of Borj Cedria (CBBC) in the Tunisian winter of 2012. Because the seeds of M. truncatula are dormant, a mechanical scarification was carried out by using sandpaper Q600. Once the seeds were scarified, they were directly transplanted into black pots (diameter = 17 cm; deep = 13cm) containing two liters of sandy loam soil (Rouached et al., 2013) consisting of (cmol kg-1 dry soil) 0.23 Na+, 0.94 K+, 0.64 Ca2+, and 0.05 Cl–, and (g kg–1 dry soil) 0.23 P2O5 and 0.44 total nitrogen. The pH and the electrical conductivity of the aqueous extract (1/10) were 6.68 and 0.05 dS m–1 respectively, with three seeds per pot. At the stage of the sixth leaf all the plants were removed while only a single plant per pot was kept for treatments. Lines were grown in a randomized complete block design. Eight replicates per line and per treatment were used. Plants were submitted to three water regimes (i) control treatment without water deficit where the soil is maintained at 100% field capacity (FC), (ii) moderate (50% FC) and severe (30% FC) water deficit which were applied 21 days after seed germination. The equivalent soil moisture was determined by the method of Bouyoucos (1983). Watering of plants was done every two days using a nutrient solution (Hewitt, 1966). To maintain the constant water regime during the culture period, the amount of water absorbed by the plant or lost by evaporation was compensated regularly every two days for each level of irrigation. This quantity was determined in advance, for each pot and weighed to calculate the weight lost from the initial weight (soil in pot without any plants) for each treatment, and this difference corresponded to the amount of water lost. Eight replicates of each genotype were harvested at flowering time and eight replicates at the end of their lifecycle.
Morphological measured traits
Twenty three morphological traits related to aerial and root growth were measured for the 10 lines of M. truncatula at flowering and end of lifecycle stages (Table 1). These traits showed high levels of polymorphism and broad-sense heritability (H2) as observed in previous studies (Badri et al., 2007, 2011; Arraouadi et al., 2011). Plants were harvested at flowering stage and at the end of their lifecycle; plants were divided into leaves, stems and roots. Dry weight was obtained after oven-drying leaf, stem and root samples at 60ºC for 48 h. For each water regime, the leaf water content (LWC), the aerial water content (AWC), and the root water content (RWC) were estimated as follows:
LWC = 100(FWL-LDW)/FWL
AWC = 100(AFW-ADW)/AFW
RWC = 100(RFW-RDW)/RFW
Where FWL, AFW and RWC are the fresh weights of leaves, aerial part and roots, respectively, while LDW, ADW and RDW are the dry weights of leaves, aerial part and roots, respectively. The aerial part is the leaves plus stems.
The leaf water potential (Ψh) was measured using a pressure chamber (Soil Moisture Equipment, Santa Barbara, CA, USA) (Scholander et al., 1965).
.png)
Photosynthesis measurements
Gas exchange measurements (net CO2 assimilation rate (A), transpiration rate (E), stomatal conductance (gs), and instantaneous water-use efficiency (WUEi) as the ratio A/E were done on mature leaves using a CO2 and H2O infrared gas analyzer (Li-Cor 6200, Li-Cor Nebraska, USA). Measurements were performed between 10 and 12 am on the leaves of plants subjected to various treatments as described by Hessini et al. (2013).
Statistical analyses
A three-way analysis of variance was used to test for line, drought treatment differences, and line x treatment interaction effects. Only characters that showed a significant line x treatment interaction were used for further analysis. Means were compared using Duncan's multiple range test at 5%. Correlations between parameters were calculated using Pearson’s correlation method. The significance level of associations between morphological traits and photosynthetic parameters was set to 0.05, and adjusted for multiple comparisons by Bonferroni corrections. All analyses were performed using SPSS software (SPSS Inc. Released 2007 SPSS for Windows, Version 16.0. Chicago, SPSS Inc.). Drought response index (DRI) is considered as an indicator of tolerance to water deficit (Chen et al., 2007) and is estimated as the ratio between the values ​​observed under water deficit conditions and those obtained in the control treatment.
Broad-sense heritability (H²) for each trait was estimated as:
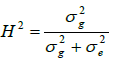
Where, σ²g is the genetic variance observed between the lines and σ²e is the environmental variance corresponding to the residual error between the eight replicates of the same genotype (=line).
Drought response indices (DRI) for measured traits in lines of M. truncatula grown under 50 and 30% of field capacity were subjected to principal component analysis (PCA). This analysis was carried out using the XLSTAT software version 7.5 (Addinsoft, 2007, New York, USA).
RESULTS
Morph-physiological variation among lines
Results from ANOVA showed that variation of morph-physiological traits was explained by the effects of line, treatment and their interaction (Table 1). The parameters were mostly influenced by the treatment factor. Out of the 27 measured characters, 14 were significantly explained by the line x treatment interaction. They include the flowering time (FLOR), number of leaves (NL), leaf area (LA), leaf fresh weight (FWL), root fresh weight (RFW), root dry weight (RDW), number of nodules (NbNOD), number of pods (NPOD), weight of pods (WPOD), weight of 100 pods (W100P), harvest index (HI), net CO2 assimilation (A) and transpiration (E), and stomatal conductance (gs).
In the control treatment, all the 14 parameters showed significant difference between lines (Table 2). The A20 line was the earliest flowering genotype while DZA45.5 was the latest flowering genotype. The largest number of pods (NPOD) was observed for TN6.18 while the lowest value was registered for A20. The net CO2 assimilation (A), transpiration (E), and stomatal conductance (gs) showed significant difference between lines. The highest net CO2 assimilation (A) and transpiration (T) were noted for the F83005.5 line while the lowest values ​​were observed for DZA45.5.
.png)
Of the 14 parameters, 11 and 12 showed significant reductions between lines in 50 and 30% FC respectively. Reductions were more significant for most traits in lines under severe (30% FC) than moderate (50% FC) water deficit. In 50% FC, the TN6.18 line showed the greatest reduction for NPOD and WPOD while TN1.11 and TN8.20 exhibited the highest levels for leaf area (LA). Furthermore, the largest reductions for A, E and gs were recorded for the A20 line. In 30% FC, the highest reductions for A and E were registered for DZA315.16 and F83005.5.
The broad-sense heritability (H2) of traits ranged from 0.13 to 0.93, from 0 to 0.92, and from 0.09 to 0.97 under the control treatment as well as for 50 and 30% FC respectively (Table 3). Of the 14 traits, 9, 11 and 10 showed high heritability values ​​under control treatment, 50 and 30% FC respectively. In the control treatment, high heritability (H2>0.4) values were recorded for FLOR, NL, LA, RFW, NbNOD, NPOD, W100P, A, and E. Moderate values (0.2≤H2≤0.4) were recorded for RDW, HI and gs, and low levels of heritability (H²<0.2) were registered for FWL and WPOD.
.png)
On the other hand, high broad-sense heritability (H²>0.4) under moderate water deficit (50% FC) were observed for FLOR, NL, LA, RFW, RDW, NbNOD, NPOD, W100P, A, E, and gs. Moderate value ​​was recorded for FWL while low levels of heritability were registered for WPOD and HI characters. Lastly, under severe water stress (30% FC), high heritability levels were observed for FLOR, NL, LA, FWL, RFW, NPOD, W100P, A, E, and gs. Moderate value was recorded for RDW while low values were registered for NbNOD, WPOD and HI traits.
Of the 252 possible correlations between measured parameters, 34, 41 and 33 were significant for the control treatment, as well as for 50 and 30% FC respectively (Table 4). Among these correlations, 23, 30, and 23 are positive in control treatment, 50 and 30% FC, respectively. Flowering time is positively correlated with NL and NbNOD, while it is negatively correlated with LA and A. The NL is positively correlated with root growth and pods production. Photosynthetic parameters (A, E and gs) are positively correlated with LA in 50% FC while they are negatively correlated with RFW in 30% FC. Comparison between the three matrices between parameters measured in lines of M. truncatula under control treatment, 50% FC and 30% FC (Table 4) showed specific correlations. In the control treatment, the NbNOD is negatively correlated with HI and A. In 50% FC, the LA is negatively correlated with NbNOD and W100P while the NPOD is positively correlated with E and gs, and the W100P is negatively correlated with A and gs. In 30% FC, the FLOR is positively correlated with FWL and RDW while NL is negatively correlated with LA as well as the RFW is negatively correlated with A and gs. Furthermore, the RDW is negatively correlated with HI.
.png)
Principal component analysis (PCA)
Principal component analysis showed that the first three principal components with eigenvalues >1 explained 72.40 and 71.31% of the total variability among genotypes grown under 50 and 30% FC, respectively, for the 15 measured parameters (Table 5). In 50% FC, the relative magnitude of the eigenvectors from the first principal component (28.15%) indicated that the flowering time, the number and weight of pods and the harvest index were the most important contributing traits. For the second principal component which explained 24.01% of the total variation, the most contributing characters were the root fresh and dry weight. The third principal component explained 20.25% of the total variation with weight of 100 pods and stomata conductance as the major contributing characters. In PCA three-dimensional graph, the 10 lines were clustered into three groups(Figure 2). The first group consisted of TN8.20, TN1.21, TN1.11, TN8.3 and DZA45.5. The second group was formed by Jemalong A17, A20, and F83005.5. The third group composed of DZA315.16 and TN6.18. The A20, TN6.18 and TN8.3 are positively associated with PC1 (Table 5). Jemalong A17, TN8.20 and F83005.5 are negatively correlated with PC1. TN1.11, TN1.21 and TN8.20 are positively correlated with PC2. A20 and DZA45.5 are positively associated with PC3. F83005.5 is negatively correlated with PC1 and PC2, Jemalong A17 is negatively associated with PC2, and TN6.18 and TN8.20 are negatively associated with PC3.
.png)
.png)
In 30% FC, the first component accounted for 29.58% of the total variation between studied lines while the most contributing parameters are the root fresh and dry weights, the number of pods and the harvest index. The second component explained 24.47% of the total variation among the 10 lines while the most contributing parameters are the photosynthetic rate, the transpiration rate and the stomata conductance. The third component accounted for 17.26% of the total variation among lines with the number and area of leaves being the most contributing characters. The scatter diagram of studied lines defined by the three first principal components shows three groups (Figure 2). The first group consisted of Jemalong A17 while the second group was formed by DZA45.5 and F83005.5, and the third group was composed by the remaining lines. The lines DZA45.5 and Jemalong A17 are negatively associated with PC1 while TN1.21 and TN8.3 are positively associated with PC1(Table 5). DZA45.5, TN8.20 and TN8.3 are positively correlated with PC2 while DZA315.15, F83005.5 are negatively associated with PC2. Jemalong A17 and TN6.18 are positively associated with PC3 while DZA45.5 and F83005.5 are negatively correlated with PC3. Overall, our results showed that similarities between lines of M. truncatula were dependent on the treatment effects (50 and 30% FC).
DISCUSSION
Lines of M. truncatula are potentially subjected to different selection pressures (Badri et al., 2007; Lazrek et al., 2009) in their natural areas of distribution therefore variability is expected in response to water deficit in the different genotypes used in this study. Lines may have developed an adaptive response to drought that can be exploited later to identify the genetic bases of tolerance to drought. The analysis of 27 morph-physiological characters showed significant variation among the 10 lines of M. truncatula. The treatment factor explains the maximum of variation of measured traits. Out of the 27 characters, 14 were explained by the interaction treatment x line indicating an involvement of these parameters in the adaptation of lines to water stress.
Positive correlations between flowering time and growth traits showed that late flowering lines have well-developed vegetative organs. The same observation was noted in chickpea (Soltani et al., 2006) and populations of M. laciniata and M. truncatula (Badri et al., 2007). Indeed, it has been shown that the genotype of plants, temperature and photoperiod factors affect flowering time in chickpea (Berger et al., 2006; Soltani et al., 2006) and rice (Ouk et al., 2006). For some accessions of M. truncatula, vernalization is a significant factor in reducing time to flowering when days are long and temperatures are moderate (e.g. 16 h/8 h and 21-25°C) (Chabaud et al., 2006). Under greenhouse condition, with seasonal changes in day-length and temperature, the effect of vernalization on time to flowering is secondary to the photoperiodic response. Annual Medicago grow best under long days and warm temperatures (De Ruiter and Taylor, 1979) and on neutral to alkaline soils (Nichols et al., 2010). Early flowering is an adaptation to escape adverse conditions, as observed in populations of chickpea in India (Berger et al., 2006). This adaptive characteristic allows lines of M. truncatula to survive in unfavorable regions for growth by restricting their development to short rainy periods of the year. This result is consistent with results observed in Algerian populations of M. truncatula (Si Ziani and Abdelguerfi​, 1995), where flowering time was correlated with altitude and annual rainfall of collection sites. A high growth rate can potentially compensate for low plant vigor at flowering, but this reduces their tolerance to drought (Mitchell-Olds, 1996).Our results showed that DZA315.16 line allocates more effort to root growth under both irrigation regimes (50 and 30% FC). The importance of the root system in the acquisition of water has long been recognized (Lynch et al., 2014). The development of the root system increases the water absorption and maintains the osmotic pressure. An increase in the root system under water stress has been reported in sunflower (Tahir et al., 2002), and in Sesuvium portulacastrum (Slama et al., 2007). Water deficit reduces the growth of leaves more compared to roots because the growth of roots and leaves exhibits different sensitivities to reduced water availability (Verslues et al., 2006). This implies a reallocation of carbon (C) to the roots, even if the rate of net assimilation of C is reduced due to the low diffusion of CO2 caused by the limited stomata opening under impeding water loss (Cornic, 2000). The water deficit potentially increased availability of C for roots (Hummel et al., 2010).
In the current study, the reduction of growth is associated with a decrease in the number of leaves. Strong positive correlations were recorded between the biomass of whole plant and number of leaves (NL) for studied lines under control treatment and drought stress. Indeed, plants reduce water loss by restricting their leaf area transpiration.
The decrease of plants growth under water deficit is also due to an inhibition of photosynthesis (Cornic, 2000). In this study, lines of M. truncatula showed a decrease in net CO2 assimilation under water deficit, which is the highest value recorded for F83005.05. This decrease is mainly due to the closure of stomata and fewer leaves and/or leaf area as part of a strategy adopted by plants to reduce their transpiring surface. The reduction in the number as well as the biomass of leaves suggest that net CO2 assimilation (A) per unit of leaf area is higher in plants subjected to drought stress than those grown in non-limiting conditions. Accordingly, Slama et al. (2007) reported a decrease of number and size of leaves and an increase in net CO2 assimilation in leaf growth rate per unit under water deficit in S. portulacastrum. This reduction saves water for the plant and is therefore considered as a criterion for adaptation to drought. Our results showed negative correlations between net CO2 assimilation (A) and flowering time (FLOR), indicating that plant growth was affected under water deficit due to reduced capacity of photosynthetic organs (that is, the amount of assimilates). This reduction can be harmful, from an agronomic point of view, because it is irreversible when the water conditions become favorable after droughts (Flexas and Medrano, 2002). According to Chaves et al. (2003), most plants tend to increase their water use efficiency (WUE) when they are subjected to moderate drought stress. This increase is due to a non-linear relationship between carbon assimilation and stomata conductance. Stomata conductance is one of the main factors affecting photosynthesis of plants (Medrano et al., 2002). It appears that plants use efficient mechanisms to limit the loss of water before the inhibition photosynthesis.
High to moderate values ​​of heritability (H2) were registered for most measured parameters in the control treatment, 50 and 30% FC, indicating that much of the variation of these traits is under genetic control. The spatial structure of studied lines in the three first principal component based on their responses to water deficit in 50 and 30% FC showed that lines were differently affected by drought stress (Figure 2). In 30% FC, Jemalong A17 exhibited the lowest reductions for FWL, NPOD, W100P, and HI while it showed the highest reduction for LA. Hence, all populations of recombinant inbred lines derived from crosses between Jemalong A17 and the remaining lines (TN1.21, TN1.11, A20, DZA315, DZA45 and F83) (Figure 1) are useful for the identification of the genetic determinants for tolerance to water deficit in M. truncatula.
Overall, the variability of measured parameters was mainly explained by the treatment factor. High to moderate broad-sense heritability (H²) were registered for most traits under control treatment and drought stress. The high heritability of most traits indicates that genetic factors are most likely controlling these characteristics even in water deficit conditions. Further genetic studies will be required to dissect the specific genetic factors that are responsible for these traits. Most of the correlations between the characters are positive under the three water regimes. The tolerance of lines to water deficit seems to depend, in particular, on their aptitude to maintain a good photosynthetic activity. Knowledge obtained by studying this model plant can be transferred to other crop and forage legumes.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors thank Ken Moriuchi and Marshall Keyster for the helpful comments on the manuscript, and Kamel Hessini and Fethia Zribi for the photosynthesis measurements. They are grateful to Mohamed Elarbi Aouani for the useful discussion, Mohamed Salhi and Najah Ben Cheikh for assistance in the greenhouse. This work was supported by the Tunisian Ministry of Higher Education and Scientific Research (LR10 CBBC 02), and the FP6 European Grain Legume Integrated Project.
REFERENCES
|
Anjum SA, Xie XY, Wang LC, Saleem MF, Man C, Lei W (2011). Morphological, physiological and biochemical responses of plants to drought stress. Afr. J. Agric. Res. 6:2026-2032. |
|
|
Arraouadi S, Chardon F, Huguet T, Aouani ME, Badri M (2011). QTLs mapping of morphological traits related to salt tolerance in Medicago truncatula. Acta Physiol. Plant. 33:917-929. |
|
|
Badri M, Chardon F, Huguet T, Aouani ME (2011). Quantitative Trait Loci associated with drought tolerance in the model legume Medicago truncatula. Euphytica 181:415-428. |
|
|
Badri M, Ilahi H, Huguet T, Aouani ME (2007). Quantitative and molecular genetic variation in sympatric populations of Medicago laciniata and M. truncatula (Fabaceae): Relationships with ecogeographical factors. Genet. Res. 89:107-122. |
|
|
Barker DG, Bianchi S, Blondon F, Datté Y, Duc G, Essad S, Fament P, Gallusci P, Génier G, Guy P, Muel X, Tourneur J, Dénarié J, Huguet T (1990). Medicago truncatula, a model for studying the molecular genetics of the Rhizobium-legume symbiosis. Plant Mol. Biol. Rep. 8:40-49. |
|
|
Berger JD, Ali M, Basu PS, Chaudhary BD, Chaturvedi SK, Deshmukh PS, Dharmaraj PS, Dwivedi SK, Gangadhar GC, Gaur PM, Kumar J, Pannu RK, Siddique KHM, Singh DN, Singh DP, Singh SJ, Turner NC, Yadava HS, Yadav SS (2006). Genotype by environment studies demonstrate the critical role of phenology in adaptation of chickpea (Cicer arietinum L.) to high and low yielding environments of India. Field Crop Res. 98:230-244. |
|
|
Bouyoucos C (1983). Les propriétés physiques du sol dépendent de sa texture et de sa structure. In Les bases de la production végétale. Tome 1. Collection Sciences et Technique agricoles, pp67-87. |
|
|
Chabaud M, Lichtenzveig J, Ellwood S, Pfaff T, Journet EP (2006). Vernalization, crossings and testing for pollen viability. In The Medicago truncatula handbook. Mathesius U, Journet EP, Sumner LW (eds). ISBN 0-9754303-1-9. |
|
|
Chaves MM, Maroco JP, Pereira JS (2003). Understanding plant responses to drought - from genes to the whole plant. Funct. Plant Biol. 30: 239-264. |
|
|
Chen C, Tao C, Peng H, Ding Y (2007). Genetic analysis of salt stress responses in Asparagus Bean (Vigna unguiculata (L) ssp Sesquipedalis Verdc). J. Hered. 98(7):655-665. |
|
|
Cornic G (1994). Drought stress and high light effects on leaf photosynthesis. Photoinhibition of photosynthesis 17:297-313. |
|
|
Cornic G (2000). Drought stress inhibits photosynthesis by decreasing stomatal aperture–not affecting ATP synthesis. Trends Plant Sci. 5:187-188. |
|
|
De Ruiter JM, Taylor AO (1979). Effects of temperature, photoperiod, and vernalization on flowering. New Zeal. J. Exp. Agr. 7(2): 153-156. |
|
|
Dong XJ, Zhang XS (2000). Special stomatal distribution in Sabina vulgaris in relation to its survival in a desert environment. Trees-Struct. Funct. 14:369-375. |
|
|
Flexas J, Medrano H (2002). Drought-inhibition of photosynthesis in C3 plants: Stomatal and non-stomatal limitations revisited. Ann. Bot. 89(2):183-189. |
|
|
Harb A, Krishnan A, Ambavaram MMR, Pereira A (2010). Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiol. 154(3):1254-1271. |
|
|
Hessini K, Ben Hamed K, Gandour M, Mejri M, Abdelly C, Cruz C(2013). Ammonium nutrition in the halophyte Spartina alterniflora under salt stress: Evidence for a priming effect of ammonium: Plant Soil 370:163-173. |
|
|
Hewitt EJ (1966). Sand and water culture methods used in the study of plant nutrition. Commonw. Agric. Bur. 22:431-446. |
|
|
Hummel I, Pantin F, Sulpice R, Piques M, Rolland G, Dauzat M, Christophe A, Pervent M, Bouteillé M, Stitt M, Gibon Y, Muller B (2010). Arabidopsis plants acclimate to water deficit at low cost through changes of carbon usage: an integrated perspective using growth, metabolite, enzyme, and gene expression analysis. Plant Physiol. 154(1):357-372. |
|
|
Lazrek F, Roussel V, Ronfort J, Cardinet G, Chardon F, Aouani ME, Huguet T (2009). The use of neutral and non-neutral SSRs to analyze the genetic structure of a Tunisian collection of Medicago truncatula lines and to reveal associations with eco-environmental variables. Genetica 135:391-402. |
|
|
Lynch JP, Chimungu JG, Brown KM (2014). Root anatomical phenes associated with water acquisition from drying soil: targets for crop improvement. J. Exp. Bot. |
|
|
Medrano H, Escalona JM, Bota J, Gulias J, Flexas J (2002). Regulation of photosynthesis of C-3 plants in response to progressive drought: Stomatal conductance as a reference parameter. Ann. Bot. (Lond) 89:895-905. |
|
|
Mitchell-Olds T (1996). Genetic constraints on life-history evolution: Quantitative-trait loci influencing growth and flowering in Arabidopsis thaliana. Evolution 50:140-145. |
|
|
Nichols P, Loi A, Nutt B, Snowball R, Revell C (2010). Domestication of new Mediterranean annual pasture legumes. In Sustainable use of genetic diversity in forage and turf breeding, Huyghe C (ed.). pp. 137- |
|
|
Nunes C, Araújo SS, da Silva JM, Fevereiro MPS, da Silva AB (2008). Physiological responses of the legume model Medicago truncatula cv. Jemalong to water deficit. Environ. Exp. Bot. 63:289-296. |
|
|
Ouk M, Basnayake J, Tsubo M, Fukai S, Fischer K, Cooper M, Nesbitt H (2006). Use of drought response index for identification of drought tolerant genotypes in rainfed lowland rice. Field Crop. Res. 99:48-58. |
|
|
Rouached A, Slama I, Zorrig W, Jdey A, Cukier C, Rabhi M, Talbi O, Limami AM, Abdelly C (2013). Differential performance of two forage species: Medicago truncatula and Sulla carnosa under water deficit stress and recovery. Crop Pasture Sci. 64(3):254-264. |
|
|
Si Ziani Y, Abdelguerfi A (1995). Comportement de 54 populations de Medicago truncatula Gaertn. dans deux sites différents; relation avec les facteurs du milieu d'origine. Meeting of FAO European sub-Network on Mediterranean Pastures and fodder Crops, Avignon (Fr.). Cah. Opt. Méditerranéennes 72:29-32. |
|
|
Slama I, Ghnaya T, Messedi D, Hessini K, Labidi N, Savouré A, Abdelly C (2007). Effect of sodium chloride on the response of the halophyte species Sesuvium portulacastrum grown in mannitol-induced water stress. J. Plant Res. 120:291-299. |
|
|
Soltani A, Hammer GL, Torabi B, Robertson MJ, Zeinali E (2006). Modeling chickpea growth and development: Nitrogen accumulation and use. Field Crop. Res. 99:24-34. |
|
|
Tahir MHN, Imran M, Hussain MK (2002). Evaluation of sunflower (Helianthus annuus L) inbred lines for drought tolerance. Int. J. Agric. Biol. 3:398-400. |
|
|
Tang H, Krishnakumar V, Bidwell S, Rosen B, Chan A, Zhou S, Gentzbittel L, Childs KL, Yandell M, Gundlach H, Mayer KFX, Schwartz DC, Town CD (2014). An improved genome release (version Mt4.0) for the model legume Medicago truncatula. BMC Genom. 15:312. |
|
|
Thudi M, Upadhyaya HD, Rathore A, Gaur PM, Krishnamurthy L, Roorkiwal M, Nayak SN, Chaturvedi SK, Basu PS, Gangarao NVPR, Fikre A, Kimurto P, Sharma PC, Sheshashayee MS, Tobita S, Kashiwagi J, Ito O, Killian A, Varshney RK (2014). Genetic dissection of drought and heat tolerance in chickpea through genome-wide and candidate gene based association mapping approaches. PloS one 9(5):1-12. |
|
|
Wang TZ (2014). Recent research progress on MicroRNA's in Medicago truncatula. Clon. Transgen. 3:2. |
|
|
Verslues PE, Agarwal M, Katiyar-Agarwal S, Zhu J, Zhu JK (2006). Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 45(4):523-539. |
|
|
Xu J, Yuan Y, Xu Y, Zhang G, Guo X, Wu F, Wang Q, Rong T, Pan G, Cao M, Tang Q, Gao S, Liu Y, Wang J, Lan H, Lu Y (2014). Identification of candidate genes for drought tolerance by whole-genome resequencing in maize. BMC Plant Biol. 14:83. |
|
|
Zhang J-Y, Cruz de Carvalho MH, Torres-Jerez I, Kang Y, Allen SN, Huhman DV, Tang Y, Murray J, Sumner LW, Udvardi MK (2014). Global reprogramming of transcription and metabolism in Medicago truncatula during progressive drought and after rewatering. Plant Cell Environ. 37(11):2553-2576. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


