ABSTRACT
Khat (Catha edulis Vahl) is an evergreen shrub habitually ingested for its euphoric and stimulatory effects. The crop is grown in the Middle East, Somalia, East Africa and Ethiopia. It is referred to as Miraa in Kenya, while in Yemen it has several names like Qat, Kat, Kath, Gat, Chat and Tschat. It belongs to the sub-order Rosidae, family Celastraceae and characterized by astringent taste. Despite the daily use and consumption of khat by millions of people in Kenya, little is known about its phenotypic. Phenotypic characterization is an essential approach for assessment of khat diversity; however, it is limited by morphological plasticity and multiple lineage evolution. The study aimed at evaluating the phenotypic diversity of selected khat cultivars grown in Embu and Meru Counties. Ninety samples from 18 cultivars were collected for phenotypic characterization. MINITAB 17 Software was used for description of principal component and construction of dendrogram using the Euclidean distance tool where 58.7% variability was observed among 13 traits studied in 90 samples of khat. Phenotypes grouped into 2 clusters phenotypic diversity showed considerable variability based on 13 khat traits. This will be useful in breeding and characterization programmes of khat cultivars.
Key words: Phenotypic, khat, diversity, cultivar.
Khat (Catha edulis Vahl) is ever green and an edible plant (Ngari et al., 2018). It is classified in the kingdom Plantae, class Magnoliopsida, order Celastrales, family Celastraceae, genus Catha, and species edulis (Sikiru, 2012). The plant was first described a by Swedish botanist called Peter Forskal. He encountered the plant as he travelled to Yemen through Egypt in an expedition that was paid for by King of Denmark Friederick, who wanted all the natural collections (Al Motarreb et al., 2002). Countries and communities have different names for the plant such as Qat and Chat in Yemen and Ethiopia, Jaad and Qaad in Somalia, Muguka and Miraa in Kenya, and Jimma in the Oromo language. In most western countries, it is recognized as khat (Ngari et al., 2018).
Khat is said to have originated from Ethiopia and then spread to East Africa and Yemen (El-Menyar et al., 2015). In Kenya, khat is grown in Meru and Embu Counties for commercial purposes. In Meru County, it is grown around Nyambene hills in Meru North, 320 km North East of Nairobi and the main outlet is Chyulu Maua Town (Nyongesa and Onyango, 2010). In Embu county, khat is mainly found in lowland area of Mbeere region
which are usually dry (Kiunga et al., 2016).
Khat also flourishes in arid and semi-arid environments where temperatures range from 5 to 35°C with free draining soil (New Agriculturalist, 2007; Nyongesa and Onyango, 2010). Khat leaves are harvested in the morning then covered with fresh banana leaves and polythene bags to maintain freshness (Nyongesa and Onyango, 2010). Khat is more potent during the dry and sunny season of the year (Ng’ethe, 2012). Harvesting is done throughout the year and planting is spread over a period of time to obtain a continuous supply. Khat is usually consumed by chewing the leaves while fresh, although occasionally, leaves are dried, then consumed as a stimulating juice (Wabe, 2011).
Mainly, khat chewing is a male habit though it has gained popularity among women (Mwenda et al., 2003; Kiunga et al., 2016). More than 10 million people worldwide use khat for psychostimulating effect (Gitonga et al., 2017). In many countries, khat is chewed for social and psychological reasons (Al-Kholani, 2010). Current trends indicate that khat is used by all societal groups regardless of age, gender, affluence, class, education and occupation (Gesese, 2013).
Various khat cultivars are identified based on the communities that consume them. In Yemen, local cultivars of khat are described according to geographical location, growth habit and physical appearance (colour of the leaf, stem sizes and potency) (Al-Thobhani et al., 2008). It is documented that different kinds of khat vary in the extent of their pharmacological activity. Farmers in Yemen have four cultivars, namely, Abyadh, Azraq, Aswad and Ahmar categorized according to the colour of shoots and growing twig (Ngari et al., 2018). Also forty kinds of khat cultivar were recognized according to geographical origin. In Ethiopia, two major cultivars have been described as Dimma (red) or Ahde (white). Getahun and Krikorian, classified khat into three types, namely ‘madness-causing, intoxicating-like spirit and insomnia-causing based on their effects (Al-Thobhani et al., 2008; Ayana and Mekonen, 2004). Little is known of how C. edulis differs morphologically, therefore, in this study the plant was characterized morphologically.
Morphological traits have been used for classification of variety duplicates, determination of genetic diversity and correlation with characteristics of agronomic significance (Zeng, 2015). Morphological characterization of khat cultivars in Embu and Meru counties was done by assessing variations in 13 khat traits. This has been traditionally used for classification of khat cultivars. It is clear that most users’ knowledge is limited to only the narcotic effects of the plant (Al-Thobhani et al., 2008).
Khat plant cultivars are notable by the level of cathionone (or the narcotic effect of the plant) present in the plant material and also their morphological differences. The varying morphological features and cathionone levels within khat may also be as a result of genetic variations. Morphologic studies have not been conducted for this crop in Mt. Kenya region where it has become a major income earner. As a traditional method, morphological traits are used to assess genetic divergence and classify existing germplasm materials. In addition, this technique is easier, cost effective, and easy to score and requires less time and finally it does not need any technical knowledge (Malek et al., 2014).
Collection sites
The germplasm was collected from 12 major khat producing wards (Figures 1 and 2). The wards included: Maua, Kianjai, Gaiti, Kangeta, Muthaara in Meru County and Kaaga South, Kithimu, Kaaga North, Mbeti south, Mbeti North, Mavuria, and Muminji in Embu County. Meru County is found in eastern region of Kenya approximately 225 km northeast of Nairobi, it covers an area of 6,936 km2. The area receives about 1366 mm per annum. The climate of Meru is described as cool and warm with temperature ranging between 16°C during cold season and 23°C during hot warm season. Embu County is located approximately at 120 km northeast of Nairobi towards Mt. Kenya. It covers an area of 2,818 km2 and lies between latitude 0° 8” and 0° 35” South and longitude 37° 40” East.
Collection of plant materials
Khat fruits, flowers and fully grown leaves were harvested from selected khat plants for morphological characterization. A total of 90 samples were collected from locally available cultivars and three replicates picked randomly from each of the sample giving a total number of 90 khat samples where different local khat names were given.
For morphological characterization, a small branch of the khat plant was cut aseptically and each packed in between a newspaper and later transported to National Museums Herbarium Department, where identification and morphological measurements were carried out. All the information on these plants was recorded based on local names given by Meru and Embu communities as well as geographic distribution (Appendix 1). The local names given by farmers included: Kira Kieru-1, Kira Kieru-2, Kira Kieru-3, Kira Kiiru-1, Kira Kiiru-2, Kira Gitune-1, Kira Gitune-2, Muchuri, Kithara, Mutimutiri, Mugiza-1, Mugiza-2, Mugumo-1, Mugumo-2, Mugumo-3, Mugumo-4, Mugumo-5, Muguka-1, Muguka-2, Muguka-3, Muguka-4, Muguka-5, Mugukawakarimi, Gitu,Mutamucii, Mukurukuru, Muruti, Muceke, Mitune, and Mumbu.
A total of 13 morphologic traits of 90 khat samples were evaluated according to guidelines provided by Robson (1994). The traits studied were leaf length, leaf width, leaf margin, petiole length, inflorescence length, peduncle length, sepal length, petal length, and diameter of the stamen, diameter of the filament, ovary length, capsule length and the length of the wing (Appendix 2). For each of the khat sample, 3 leaf samples were randomly selected and their measurement taken using a digital vernier caliper. Flowers and fruits measurements were taken using wild MSA Switzerland dissecting microscope at magnification of ×10.
Data management and statistical analysis
Raw data was entered into Microsoft excel spreadsheet which was then imported to Minitab software version 17.0 (State College Pennsylvania-USA) software. Data was analyzed statistically for the differences in means for 13 khat traits, through one-way ANOVA followed by Tukey’s post hoc test. Statistical significant differences were set at p≤0.05. Cluster analysis yielded a dendrogram that was used to examine the morphological relatedness among the 90 khat samples while Principal Component Analysis (PCA) was used to assess the underlying source of variation in morphology.
The measurements of the 13 phenotypic traits studied were found to vary across the 90 khat samples, Muguka-4 had the highest mean leaf length of 99 mm, while Mutimutitiri had the lowest mean leaf length of 53.67 mm. The mean leaf length of the 90 khat samples had significant differences. Regarding leaf width, Muguka-4 had the highest mean value of 45.67 mm, while Kira Kieru-1 had the lowest mean leaf length of 19.33 mm (Appendix 2).
The mean sepal length of the studied samples did not show significant differences among themselves with the highest mean length of 1.47 mm being recorded in Mugumo-1, Mugiza-1, and Kithara. The lowest mean sepal length of 1.03 mm was observed in Mugumo-3 and Mugumo-4, Mugukawakarimi, Mukurukuru, Mumbu, Muruti, Mutamucii and Mutimutiri. The highest mean petal length of 1.67 mm was observed in Kira Gitune-1, Kira Kieru-1 and Kira Kiiru-1 cultivar from Meru, while the lowest mean petal length of 1.00 mm was observed in Muguka-2. The mean petal length of the 90 khat samples did not show significant differences among themselves. The mean stamen diameter of the studied khat samples did not show significant differences with the highest diameter of 2.37 mm being shown in Kira Gitune-1 and the lowest mean diameter being 1.07 mm in Kira Kieru-2 (Appendix 2).
The mean filament diameter of all samples ranged from 1.53 to 1.07 mm. The highest mean filament diameter was observed in Muchuri and Kira Kiiru-1, while the lowest filament diameter was shown in Gitu, Kira Kieru-2, Muguka-4, Muguka-5, Muguka-1, Muguka-2, Mugukawakarimi, Mugumo-3, Mukurukuru, Muruti and Mutimutiri cultivars. There were no significant differences in ovary length mean among all cultivar with Mugiza-1 showing the highest length of 1.57 mm, while Kira Kieru-3 and Kira Kiiru-2 showed the lowest mean ovary length of 1.10 mm. There were significant differences in mean capsule length among the 90 khat samples with the highest capsule length of 8.43 mm being observed in Gitu and lowest mean capsule length of 5.1 mm in Kira Gitune-1, and Kira Kieru-3. The mean wing length varied significantly among various cultivar with the highest mean wing length of 3.5 mm in Muguka-1 and lowest wing length of 2.07 mm in Kira Gitune-1.
The leaf margin had wide variations ranging from a mean of 107.33 mm (Muguka-4) to a mean of 55.00 mm (Mutimutiri). The mean leaf margin of Mutimutiri cultivar was significantly lower than the other cultivars (p≤0.05).
Besides, the mean petiole length did not show significant differences among the cultivar with the highest mean petiole length being 5.67 mm (Muguka-1 and Muguka-2) and the minimum mean petiole length being 2.33 mm observed in Mugiza-1 and Muguka-3 cultivars (p˃0.05). The mean inflorescence length of 90 khat samples did not show significant difference among the 90 samples. Kira Kieru-1 had the highest mean inflorescence length of 21.00 mm, while Mutimutiri had the lowest inflorescence length of 14.33 mm. The mean peduncle length of 30 samples was not significantly different with Kira Gitune-1 having the highest mean of 7.33 mm, and Mugumo-1, Mugumo-2 and Mugiza-1 having lowest mean peduncle length of 4.00 mm (p˃0.05).
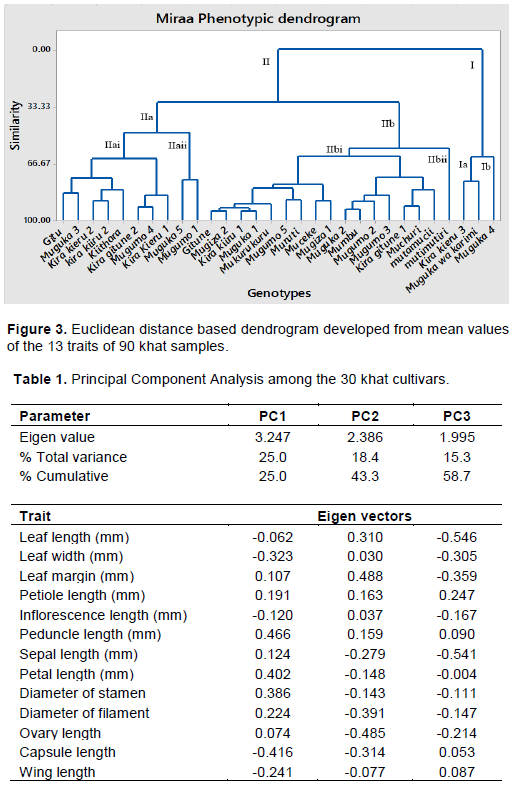
A dendrogram constructed from data set of mean values of the 13 khat traits showed 2 super clusters namely I and II. Super cluster I had two sub clusters Ia and Ib. The sub cluster Ia comprised Mugukawakarimi and Kira kieru-3 cultivars from Embu and Meru counties, respectively, while sub cluster Ib had Muguka-4 cultivars from Embu county, clustering independently. The sub cluster II was more diverse and clustered into two sub clusters IIa and IIb. The sub cluster IIa was divided into IIai and IIaii sub clusters. The sub cluster IIai had three groups where the first group comprised Gitu and Muguka-3 cultivars from Embu County while the second group comprised Kira kiiru-2, Kira kieru-2 and Kiithara cultivars from Meru County. The third group comprised Kira gitune-2 and Kira kieru-1 cultivars from Meru county and Mugumo-4 cultivar from Embu county. The sub cluster llaii had one group which comprised Muguka -5 and Mugumo-1 genotypes both from Embu County.
The sub cluster IIbi had four groups. The first group comprised of Gitune, Muruti, Mukurukuru Mugumo-5, Mugiza-2 and Muguka-1 cultivar from Embu County and Kira kiiru-1 cultivar from Meru County. The second group comprised Muceke and Mugiza-1 from Embu County. The third group comprised Mugumo-2, Muguka-2, Mugumo-3 and Mumbu from Embu County, while the fourth group comprised Kira gitune-1 and Muchuri cultivar from Meru county, and Mutamucii from Embu county. The sub cluster IIbii had Mutimutiri cultivar which originated from Embu County (Figure 3).
Principal component analysis (PCA)
The PCA was performed for all the 13 traits in the 90 khat samples as indicated in Table 1. Out of the 13 traits, three principal components (pc1, pc2 and pc3) exhibited more than one Eigen value and showed about 58.7% variability among the 13 studied traits. The three principal components: pc1, pc2 and pc3 had 25.0, 18.4 and 15.3% variability, respectively among the cultivars for the traits under study. The pc1, pc2 and pc3 had Eigen values of 3.247, 2.386, and 1.995, respectively.
The pc1 was positively correlated to leaf margin, petiole length, peduncle length, sepal length, petal length, stamen diameter, filament diameter and ovary length. However, it was negatively correlated to leaf length, leaf width, inflorescence length, capsule length and wing length. The pc2 showed a highly positive correlation to leaf length, leaf margin, petiole length, inflorescence length and peduncle length. However, it was negatively correlated to sepal length, petal length, stamen diameter, filament diameter, ovary length and wing length. The pc3 showed a positive correlation to petiole length, capsule length, peduncle length and wing length but negatively correlated to leaf length, leaf width, leaf margin, inflorescence length, sepal length, petal length, stamen diameter, filament diameter and ovary length (Table 1).
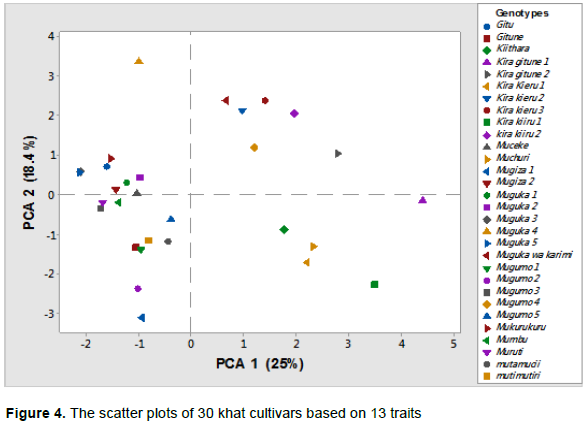
The scatter plot of the 30 khat samples based on the 13 studied traits was also drawn in this study. It was observed that the first quadrant comprised of Muguka-1, Muguka-2, Muguka-3 and Mugiza-2, Muguka wa karimi, muguka-4, Muguka-5 and Gitu cultivars, which were from Embu county. The second quadrant had six cultivars, comprising Kira kieru-2, Kira kieru-3, Kira kiiru-2, Kira gitune-2, from Meru County, as well as Mugumo-4 and Mukurukuru cultivar from Embu County. The third quadrant comprised Mugumo-3, Mugiza-1, Mugumo-1, Mugumo-2, Gitune, Mutimutiri, Muruti, Mumbu, Muceke and Mugumo-5 cultivar from Embu County. The fourth quadrant comprised five cultivar, which originated from Meru county and they included Kira gitune-1, Kiithara, Muchuri, Kira kieru-1 and Kira kiiru-1 (Figure 4).
The mean leaf length of different cultivars indicated significant differences, whereby Muguka-4 cultivar recorded the highest mean leaf length while Mutimutiri cultivar showed the lowest mean leaf length. Both cultivars were collected from the same geographical location and environmental conditions. The lowest leaf length of Mutimutiri cultivar could be due to the ability of the cultivar to adapt to relatively drier and saline environment. Plants growing in such conditions usually have reduced leaf area in order to minimise loss of water during evaporation (Deblonde and Ledent, 2001).
The mean leaf width of different cultivar was significantly different, Muguka-4 cultivar recorded the highest mean leaf width while Kira Kieru-1 recorded the lowest leaf width. The lowest leaf width of Kira Kieru-1 could be as a result of adaptations to conserve water. Kira Kieru-1 had narrow leaves and less total surface area than Muguka-4 and, therefore, loses more water compared to Kira Kieru-1. Studies have shown that broad leaves heat up more than narrow leaves of the same length, hence narrow leaf plants are well adapted to dry and hot environments. The cultivar with the highest mean leaf width (Muguka-4) could have been in moist and shady environments, which enhanced their ability to absorb sunlight. This was confirmed in tropical vines (Tardieu, 2013).
Cultivars with long inflorescence produce more flowers, fruits and seeds compared to cultivars with short inflorescence (Rahman et al., 2009). This suggests that both the number of inflorescences and the number of female flowers are the main factors in determining yield. On the other hand, determining the number of inflorescences per plant and the number of female flowers both depend on environmental factors (Sangoi, 2001; Domiciano et al., 2014). Flowering and determination of the flower type are influenced by the occurrence of low temperatures and high rainfall (Inouye, 2008; Prasad et al., 2001). Therefore, the highest inflorescence length in Kira Kieru-1 could have resulted from high rainfall and low temperature, while the lowest inflorescence length in Mutimutiri could be due to low rainfall and high temperature.
The mean peduncle length showed significant differences among different cultivars. Drought and stress cause reduction in peduncle length. From this study Kira Gitune-1, which had the highest peduncle length was not drought stressed while Mugumo-1, Mugumo-2 and Mugiza-1 which had the lowest peduncle length were drought stressed. This was confirmed by Amiri et al. (2013) on 80 bread wheat genotypes.
The mean sepal length of different genotypes showed significant differences among different cultivars. The difference in sepal and petal length could be related to availability of water (Kwak et al., 2007). Therefore, the longer the petal and sepal the higher the water availability. Hence cultivars Mugumo-1, Mugiza-1, Kiithara, Kira Gitune-1, Kira Kieru-1 and Kira Kiiru-1 were in place with enough water. The mean stamen length of many cultivars showed significant differences with the highest diameter of 2.37 mm being recorded in Kira gitune-1 cultivar from Meru and the lowest mean diameter of 1.07 mm was recorded in Kira kieru-2 cultivar from Meru. Availability of water results to increased flowers size and stamen length. Consequently, the traits usually related to floral attractiveness are increased (Natalia et al., 2015). However, fecundity potential is not increased. These results suggest that population differentiation in floral characters could be a caused by random genetic drift that occur in relatively small or isolated populations. It may also be attributed to restricted gene flow (Medrano et al., 2005).
The mean filament diameter showed significant difference among the cultivar with mean filament diameter ranging between 1.53 and 1.07 mm. Kira Kiiru-4 had the highest, while Gitu, Muguka-4 and Muguka-5 had the least 1.07. The floral parts of a plant are highly affected by the environment and this could be reasons as to why significant difference was recorded in filament diameter. The mean ovary length of all cultivars indicated no significant difference. This could be due to similar ancestry of all the cultivars studied. The mean capsule length showed significant difference among 30 khat samples, such could be used to discriminate cultivars. Gitu had the highest capsule length while Kira gitune-1 and Kira kieru-3 cultivar had the lowest. The difference in position of the inflorescence or the fruit could account for the variation in the capsule length (Buide, 2008). From the results it indicates that Gitu fruit was far from the stalk, while Kira Gitune-1 and Kira Kieru-1 fruit was near the stalk.
The mean length of the wing was significantly different among the various cultivars. Muguka-1 cultivar from Embu had the highest and Kira Gitune-1 had the lowest. This could be explained by the possibility that the age of the plant could have had an impact on the length of the seed wing (Georg-kraemer et al., 2004). The one with high mean wing length could be older compared to the one with short wing length. From the results this could mean Muguka-1 cultivar was older than Kira Gitune-1 cultivar.
Principal Component Analysis (PCA) is a method that summarizes the data without much loss of information based on the similarities and the differences of the data. The PCA results of 13 traits among 30 khat samples indicated that the first principal component had a higher contribution to the total variation compared to the second and third principal components. The traits which contributed the highest variability value in PC1 included leaf margin, petiole length, peduncle length, sepal length, petal length, diameter of stamen, diameter of filament and ovary length (Table 1). This showed that these traits were responsible for the most of the diversity exhibited in the first principal component. The cumulative value of the first three principal components (58.7%) was slightly above the first principal component obtained by Mawia et al. (2015) (53.8%) in 13 rice genotypes. The principal component two showed variability of six traits while principal component three showed variability of four traits (Table 1). Leaf margin, petiole length and peduncle length contributed to variation across all the three principal components, indicating that they were most important agronomic traits in C. edulis. The first Eigen value (3.247) was close to that found by Asudi et al. (2010) who reported a value of 3.147 on phenotypic features of Kenyan papaya. The second Eigen value of 2.386 was lower than that obtained by Mawia et al. (2015) in 13 rice genotypes. Conversely, the third Eigen value of 1.995 was higher than that obtained by Mawia et al. (2015) (0.38) in 13 rice genotypes.
The distribution of the cultivars on the scatter plot indicated the existence of wide variations among the studied cultivars. Cultivars that were close to each other on the same scatter plot showed phenotypic relatedness, while cultivars that were far away from each other were regarded as phenotypically distant. For example, the graphical closeness of Mugumo-5 and Muguka-3 indicated morphological relatedness. Most of the cultivars, from the same quadrant were collected from the same geographical region. For example Mugumo-5, Mugumo-2, Mugiza-2 and Muguka-3 were found in the same quadrant and were collected from the same geographical region. Gitune, Muceke and Mumbu were from the same county. However, cultivars from different geographical regions were found clustering together for instance Kirakiiru from Meru clustered together with Muguka-1, Mutimutiri and Muguka-4 which were cultivars from Embu county, while Mugumo-4,Gitu,Mugumo-1,Muruti, MugukawaKarimi clustered together with Kira Gitune-2, Kira Kieru-1 which were cultivars from Meru county.
The dendrogram generated from 13 phenotypic traits showed two main clusters (clusters I and II), which indicated an existing variation in the cultivars. Cluster II showed high phenotypic variation, indicating the enormous phylogenetic divergence among cultivars in this cluster. Cluster II had two sub-clusters, IIa and IIb each of these sub-cluster, clustered into two subcluster again, IIai, IIaii and IIbi and IIbii. Cultivars in these cluster I included Kira Kieru-3, Mugukawakarimi and Muguka-4 cultivars where 2 cultivars were from Embu and 1 from Meru County. Most of the cultivars clustered in cluster II, some of the cultivars included Kira Kiiru-2 and Kira Kieru-2 which were collected from Meru in cluster IIai, Muguka-2, Mumbu, Mugumo-2, and Mugumo-3 cultivars from Embu was found in IIbi. In these findings, those cultivars which clustered together were collected from the different geographical location. For the scatter plot, for example Kira Kieru-2 and Kira Kiiru-2 cultivars which were on the same scatter plot showed little variation in dendrogram clustering. Further studies are recommended due to the limitations associated with morphological traits as a means of determining variations, the diversity of these khat cultivars should be validated using superior traits such molecular markers.
From this study, it was concluded that phenotypic diversity showed considerable variability; clustering of the phenotypes was not based on the geographical origin of the plant.
The authors have not declares any conflict of interests in phenotypic variations in Catha edulis along different growth intervals should be determined.
REFERENCES
|
Al Motarreb A, Baker K, Broadley KJ (2002). Khat: Pharmacological and medical aspects and its social use in Yemen. Journal of Phytotherapy Research 16(5):403-413.
Crossref
|
|
|
|
Al-Kholani AI (2010). Influence of Khat Chewing on Periodontal Tissues and Oral Hygiene Status among Yemenis. Dental Research Journal 7(1):1-6.
|
|
|
|
|
Al-Thobhani MAH, Sathyanarayana BN, Simon L, Sondur SN (2008). First comparative genotypic study on khat (Catha edulis Forsk) genotypes from Yemen.Middle East. Russian Journal of Plant Science and Biotechnology 2:1-8.
|
|
|
|
|
Amiri R, Bahraminejad S, Jalali-Honarmand S (2013). Effect of terminal drought stress on grain yield and some morphological traits in 80 bread wheat genotypes. International Journal of Agriculture and Crop Science 5(10):1145-1153.
Crossref
|
|
|
|
|
Asudi GO, Ombwara FK, Rimberia FK, Nyende AB, Ateka EM, Wamocho LS, Shitanda D, Onyango A (2010). Morphological diversity of Kenyan papaya germplasm. African Journal of Biotechnology 9(51):8754-8762.
|
|
|
|
|
Ayana AM, Mekonen Z (2004). Khat (Catha edulis Forsk). Chewing, socio-demographic description and its effects on academic performance, Jimma University Students.
|
|
|
|
|
Buide ML (2008). Disentangling the causes of intrainflorescence variation in floral traits and fecundity in the hermaphrodite Silene acutifolia. American Journal of Botany 95(4):490-497.
Crossref
|
|
|
|
|
Deblonde PMK, Ledent JF (2001). Effects of moderate drought conditions on green leaf number, stem height, leaf length and tuber yield of potato cultivars. European Journal of Agronomy 14(1):31-41
Crossref
|
|
|
|
|
Domiciano GP, Alves AA, Laviola BG, Albrecht JC (2014). Analysis of morpho-agronomic and climatic variables in successive agricultural years provides novel information regarding the phenological cycle of Jatropha in conditions of the Brazilian cerrado. Anais da Academia Brasileira de Ciências Journal 86(4):1985-1998.
Crossref
|
|
|
|
|
El-Menyar A, Mekkodathil A, Al-Thani H, Al-Motarreb A (2015). Khat use: history and heart failure. Oman Medical Journal 30(2):77-82.
Crossref
|
|
|
|
|
Georg-kraemer JE, Caierão E, Minella E, Barbosa-Neto JF, Cavalli SS (2004). The glutamates in malting barley: enzyme survival and genetic and environmental effects. Journal of the Institute of Brewing 110:303-308.
Crossref
|
|
|
|
|
Gesese D (2013). Is khat a social ill? Ethical arguments about 'stimulant' among learned Ethiopians, ASC Working Paper 108.
|
|
|
|
|
Gitonga GM, Ngeranwa J, Machocho AK, Muthee DG, Kimutai R, Gitonga AW(2017). Nephrotoxicity effects of Khat (Catha edulis) on mice when administered orally. Journal of Phytopharmacology 6(1):27-33.
|
|
|
|
|
Inouye DW (2008). Effects of climate change on phenology, frost damage, and floral abundance of montane wildflowers. Journal of Ecology 89 (2):353-362.
Crossref
|
|
|
|
|
Kiunga JL, Dossaji C, Yenesew AS (2016). A Survey of Traditional Medicinal Uses of Cathaedulis (Celastraceae) in Meru and Embu Counties of Kenya. International Journal of Ethnobiology and Ethnomedicine 3(1):1-12.
|
|
|
|
|
Kwak MS, Min SR, Lee SM, Kim KN, Liu JR, Paek KH, Shin JS, Bae JM (2007). A sepal-expressed ADP-glucose pyrophosphorylase gene (NtAGP) is required for petal expansion growth in 'Xanthi' tobacco. Plant Physiology 145(1):277-89.
Crossref
|
|
|
|
|
Malek MA, Rafii MY, Afroz SS, Nath UK, Mondal M (2014). Morphological characterization and assessment of genetic variability, character association, and divergence in soybean mutants. The Scientific World Journal pp. 1-12.
Crossref
|
|
|
|
|
Mawia MA, Wambua KF, Daniel A, Nyamai WD, Felix M (2015). Phenotypic Diversity Studies on Selected Kenyan and Tanzanian rice (Oryza sativa L): Genotypes Based on Grain and Kernel Traits. Journal of Rice Research 3(4):150-161.
|
|
|
|
|
Medrano M, Herrera CM, Barrett SCH (2005). Herkogamy and mating patterns in the self compatible daffodil (Narcissus longispathus). Annals of Botany Journal 95:1105-1111.
Crossref
|
|
|
|
|
Mwenda JM, Arimi MM, Kyama MC, Langat DK (2003). Effects of khat (Catha edulis) consumption on reproductive functions: a review. East African Medical Journal 80:318-323.
Crossref
|
|
|
|
|
Natalia SL, Jamie O, David P, Rachel C, Adams, Katarina K, Frederick V (2015). Training response inhibition to food is associated with weight loss and reduced energy intake, Journal of Appetite 95:17-28.
Crossref
|
|
|
|
|
New Agriculturalist (2007). Growing demand for Kenyan khat,Wren Media, Suffolk, UK.
|
|
|
|
|
Ng'ethe JW (2012). Factors influencing the consumption and control of Khat among the youth in Igembe South District, Meru County, Kenya (Doctoral dissertation, University of Nairobi, Kenya).
|
|
|
|
|
Ngari LN, Njagi SN, Kamau JK, Lagat RC, Musyoki AM (2018). Genotypic Characterization of Catha edulis in Mt. Kenya Region, Kenya. Journal of Phylogenetics Evolutionary Biology 6:203.
Crossref
|
|
|
|
|
Nyongesa AW, Onyango DW (2010). Khat (Catha edulis Forsk): A bane or boon to humanity. Journal of Ethnomedicine 28:187-209.
|
|
|
|
|
Prasad PVV, Craufurd PQ, Kakani VG, Wheeler TR, Boote KJ (2001). Influence of high temperature during pre-and post-anthesis stages of floral development on fruit-set and pollen germination in peanut. Functional Plant Biology Journal 28(3):233-240.
Crossref
|
|
|
|
|
Rahman I, Ahmad H, Sirajuddin I, Ahmad I, Abbasi FM, Islam M, Ghafoor S (2009).Valuation of rapeseed genotypes for yield and oil quality under rainfed conditions of district Mansehra. African Journal of Biotechnology 8(24):6844-6849.
|
|
|
|
|
Robson NKB (1994). Flora of Tropical East Africa-Celastraceae.CRC Press.
|
|
|
|
|
Sangoi L (2001). Understanding plant density effects on maize growth and development: an important issue to maximize grain yield. Ciência Rural Journal 31(1):159-168.
Crossref
|
|
|
|
|
Sikiru L (2012). Flower of paradise (Khat: Catha edulis): Psychosocial, health and sports perspectives. African Journal of Health Sciences 22:61-175.
|
|
|
|
|
Tardieu F (2013). Plant response to environmental conditions: assessing potential production, water demand, and negative effects of water deficit. Frontiers in physiology 4:17.
Crossref
|
|
|
|
|
Wabe NT (2011). Chemistry, pharmacology, and toxicology of khat (catha edulis forsk): a review. Addiction and Health 3(3-4):137-49.
|
|
|
|
|
Zeng X (2015). Genetic variability in agronomic traits of a germplasm collection of hulless barley, Journal of genetics and molecular Research: Genetics and Molecular Research 14(4):18356-18369.
Crossref
|
|