ABSTRACT
Taro (Colocasia esculenta L. Schott) is an important crop for food and nutrition security, incomes, and livelihood of people in developing countries. Its cultivation and productivity have been limited mainly due to a lack of quality planting materials. Therefore, this study developed an effective micropropagation method for two common taro varieties in Kenya (Purple Wild and Dasheen) using apical meristem derived from portions of the corm and base of leaf petioles. This study optimized the 6-benzylaminopurine (BAP) and indole-3-butyric acid (IBA) levels for a better shoot and root development. The highest shoot induction was observed in both varieties when Murashige & Skoog (MS) media was supplemented with 2 mg/L of BAP. Similarly, the maximum rooting response was achieved in both varieties on half-strength MS media supplemented with 0.5 mg/L of IBA. The study also revealed significant interactions between variety × BAP levels and variety × IBA levels (P<0.0001) for shoot and root development, respectively. This apical meristem-based micropropagation method developed in this study can be used for rapid multiplication of genetically clean planting materials for commercial uses and to establish taro transformation protocols.
Key words: Taro, 6-benzylaminopurine (BAP), indole-3-butyric acid (IBA), shoot induction, root induction, regeneration.
Abbreviation:
MS, Murashige and Skoog media; BAP, 6-benzylaminopurine; IBA, indole-3-butyric acid.
Taro (Colocasia esculenta L. Schott) is one of the edible members of the family Araceae distributed across the world, particularly in the tropics (Okonkwo, 1993). It is native to Southeast Asia (Matthews, 1991; Yoshino, 2002) and widely cultivated as a staple food crop in the Pacific Islands, Asia, and Africa (Onwueme, 1999). The global acreage under taro production for 2019 was 1.957 million hectares, with a total production of 10.54 million tons. Nigeria, China, Cameroon, Ghana, and Papua New Guinea were the top five producers of taro in the world (FAO, 2021). Taro is mainly cultivated for corm production, but the pseudo-stems and leaves are also edible and used to prepare various traditional food dishes. Taro corms are a good source of digestible starch, crude fiber, minerals, vitamin C, thiamin, riboflavin, and niacin (Boampong et al., 2019; Jane et al., 1992; Kidanemariam, 2018). Both corms and leaves are rich in crude proteins (Temesgen and Retta, 2015). Besides nutritional values, taro is an important food security crop in developing countries because of its high yield potential, that is, as high as 110 t/ha (Lebot, 2009). Although taro is primarily grown for domestic consumption, the production of taro for cash generation has been a rewarding business. Moreover, taro is adapted to a wide range of environments and agricultural practices (Kantaka, 2004), making taro a crop of choice under changing climatic conditions.
Despite the importance of taro in food and nutrition security, incomes, and livelihood of rural people, the current global productivity of taro is estimated around 5.39 t/ha, which is about 5% of its experimental yield (Lebot, 2009). Low productivity and limited cultivation of taro are attributed to several factors, including a high incidence of pest and diseases; scarcity of quality planting materials; labour-intensive traditional production system; difficulties in post-harvest handling and marketing; and low investment in taro research and extension (Onwueme, 1999). Taro is commonly propagated through vegetative means like side suckers, small corms, and corm pieces. These propagation materials are bulky in nature and low throughput, and the availability of these planting materials is seasonal. Moreover, these vegetative planting materials often serve as vehicles for various pests and diseases from one taro crop to the next crop and subsequently compromise the quality and quantity of taro production. Therefore, the availability of quality planting material in sufficient quantities has been a major challenge among taro producers. More importantly, the taro seeds system lacks an efficient and high throughput micropropagation system that helps in the mass propagation of quality planting materials.
The tissue culture or micropropagation technique provides a sustainable solution to the problems associated with conventional propagation by enabling rapid production of high-quality, disease-free, and uniform planting materials (IAEA, 2004). Since tissue culture is performed in a controlled laboratory environment, the multiplication of planting materials could be achieved all year-round. The tissue culture technique has been successfully applied on thousands of plant species (Fay, 1992; Villalobos and Engelmann, 1995; Jackson et al., 2001; Sarasan et al., 2006), but the use of this technique for in-vitro regeneration and mass production of taro planting material has been limited in Kenya primarily due to the lack of efficient regeneration and mass multiplication protocols. Several protocols for taro tissue culture are available (Yam et al., 1990; Tuia, 1997; Minas, 2002; Hossain, 2012), including the one that uses locally available nutrients to substitute MS medium (Ngetich et al., 2015). All these efforts to develop taro micropropagation protocols could be due to genotype dependent response of taro to micropropagation methods, variations in growth media and culture conditions, types of explants used in micropropagation, and to reduce the costs.
The direct and indirect shoot organogenesis are considered the best micropropagation method due to a low cost, minimal soma-clonal variations, and high throughput production system (Mukami et al., 2018; Burner and Grisham, 1995). Apical meristem culture eliminates viruses in many plant species, thus helps in the production of disease-free planting materials and achieving better yields than conventional planting materials (Wang and Valkonen, 2008, 2009). In this study, organogenesis-based regeneration protocols was developed for two common taro varieties of Kenya, that is, Purple Wild and Dasheen, through optimizing cytokinin and auxin levels for the shoot and root regeneration, respectively.
Collection and management of planting materials
Two taro varieties, Purple Wild and Dasheen were collected from Busia (0.4347 °N, 34.2422 °E), Kakamega (0.2827 °N, 34.7519 °E), Kiambu (1.0314 °S, 36.8681 °E), Kisii (0.8067 °S, 34.7741 °E), Machakos (1.5177 °S, 37.2634 °E), Meru (0.3557 °N, 37.8088 °E), Murang’a (0.7839 °S, 37.0400 °E), Nyeri (0.4197 °S, 37.0400 °E), and Siaya (0.0617 °S, 34.2422 °E) counties of Kenya in August 2017. Taro plantlets were collected in polythene bags, labelled, and transported to the greenhouse of International Livestock Research Institute, Nairobi, Kenya. The taro plantlets were established in 4 L pots (Planter 1 H200 × Dia268 mm) with autoclaved soil consisting of forest soil and manure in the ratio of 2:1 by volume. Notably, a hand full of gravel was dispensed at the bottom of each for drainage of excess water. These planets were maintained in a screen house at ambient conditions for four months before using these stock plants in subsequent experiments.
Media and explants preparation
Murashige and Skoog (MS) media was prepared using 4.4 g/L of MS media with basal salts, 30 g/L of sucrose dissolved in distilled water, and pH was adjusted to 5.8 using 0.1 N HCl and 0.1 N NaOH, and 3 g of Gelrite added to the bottle. All the media, glassware, and metallic equipment, e.g., forceps and blade holders used in experiments, were autoclaved at 121°C at 15 psi for 20 min. The explants comprising the corm top attached to the base of the petioles were harvested from stock plants in the greenhouse, trimmed to a length of 2 cm, and taken to the laboratory. The explants were thoroughly washed using running tap water for 30 min and immersed in a 1 L glass beaker filled with tap water containing soap antibacterial (Dettol) and two drops (100 µl/L) of tween 20 and swirled gently by hand for 30 min. The explants were rinsed twice, then transferred to another beaker containing 3 g/L Redomil fungicide (Metalaxyl 8% + Mancozeb 64% WP) an hour with gentle swirling at intervals, and then rinsed thrice using autoclaved-distilled water. After that, the explants were immersed in 70% ethanol for 1 min and rinsed thrice using autoclaved distilled water in a laminar flow chamber. The explants were subjected to 40% JIK regular bleach (added with 100 µl tween-20 L-1) for 20 min, rinsed thrice in autoclaved distilled water, and trimmed to a length of about 1 cm in a laminar flow chamber.
Shoot induction, multiplication, and elongation
The trimmed explants were transferred to shoot induction medium (SIM) comprising MS basal supplemented with 30 g/L sucrose and five 6-benzyl aminopurine (BAP) hormone levels (0, 0.5, 1.0, 2.0, and 3.0 mg/L). For each variety, ten explants were subjected to each BAP level and replicated three times, and the experiment was arranged in a completely randomized design. The cultured explants were maintained in the growth room with the set temperature at 24 ± 2°C for 16/8 h light/dark cycle for 30 days. The induced shoots at 15 days were transferred to fresh SIM and grown for a further 15 days at similar growth conditions to enhance shoot elongation and multiplication. Data on shoot numbers were recorded at fortnightly intervals.
Root induction
The elongated shoots of ~7 cm length generated in the SIM comprising MS basal supplemented with 30 g/L sucrose were transferred to rooting media consisting of MS basal salt supplemented with 30 mg/L sucrose and five IBA concentrations (0, 0.25, 0.5, 0.75, and 1 mg/L). For each variety five explants were subjected to each IBA level and replicated three times, and this experiment was arranged in a completely randomized design. The plantlets were then incubated in the growth room at 24±2°C under 16/8 h light/dark cycle to induce rooting. Data on the total number of roots initiated per shoot and their length was recorded after 25 days.
Hardening and acclimatization
The rooted plantlets were removed from the growth room, immersed in distilled water to remove media, and transferred individually to a disposable plastic cup that acted as a potting container. The container was first filled with gravel and then topped with forest soil and manure in the ratio of 1:2:1 by volume. The plants were labeled according to the variety and the concentration of BAP/IBA. They were grown for 25 days, transferred to the glasshouse, and placed under an improvised humidity chamber for eight days, after which they were removed and exposed to ambient temperature conditions in the greenhouse. The plants were watered daily, and plant survival rates were recorded for each treatment.
Data collection and data analysis
Data on shoots and roots development was recorded after 15 days. The number of induced roots and their length after culturing them for 15 days on the rooting medium was recorded. The percentage survival of the plantlets transferred for acclimatization was recorded after 15 days. Data were subjected to analysis of variance (ANOVA) and treatment means were compared using Duncan Multiple Range Test (DMRT) using the XLSTAT 2020 software (Addinsoft, 2021), and the graphs were plotted using R statistical software (Wickham, 2016).
Effect of BAP concentrations on shooting
The effect of different BAP concentrations on shoot induction and shoot multiplication in two taro varieties are shown in Table 1. The BAP concentrations evaluated in this study were significantly different for shoot induction and multiplication in two taro varieties (P<0.001). At 15 days, the number of shoot induced was the highest at a BAP concentration of 2.0 mg/L for both varieties (P <0.001), but varieties were indifferent for shoot induction in this concentration. The same concentration was the best performer for shoot multiplication in both genotypes, and two varieties were similar for shoot multiplication. When the BAP concentration was 3.0 mg/L, there was a significant decrease in shoot induction and shoot multiplication in both varieties (P <0.0001). A significant interaction was also noted between lower BAP concentration (0.5 mg/l) and varieties e.g., Dasheen had significantly less shoot induction and shoot multiplication than Purple Wild. A schematic of organogenesis in taro in this study is presented in Figure 1.
Effect of IBA concentration rooting
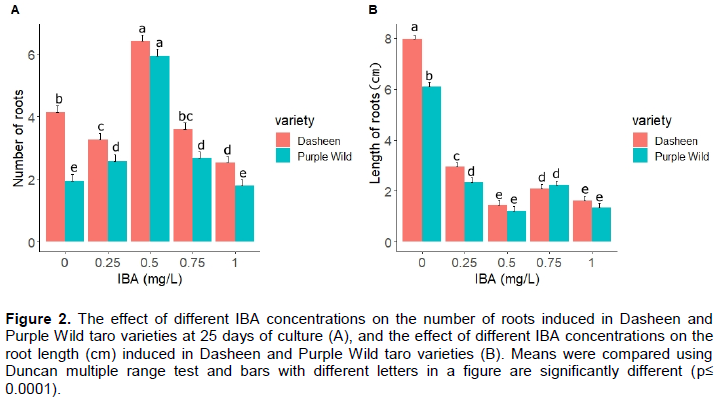
The effect of different IBA concentrations on number of roots and root length in two taro varieties are presented in Figure 2. The different IBA concentrations evaluated in this study deferred significantly for number of root production and root length (P < 0.0001). The IBA concentration of 0.5 mg/L induced the highest number of roots in both Dasheen and Purple Wild varieties compared to other treatments and number of root induction in both varieties were similar for this concentration. A gradual decline was observed in the number of root inductions in both taro varieties when IBA concentration was increased to 0.75 and 1 mg/L. Dasheen performed better than Purple Wild for root induction in all levels of IBA concentrations and control treatment, indicating significant interactions among treatments evaluated in this study for root induction (P<0.0001).
Micropropagation is the in-vitro clonal propagation of plants (Bhojwani and Dantu, 2013). Since food insecurity is still a serious global challenge, it is imperative todevelop a direct shoot organogenesis protocol for crops of economic importance as these protocols enable us to produce large numbers of new high-quality plantlets in a relatively short time and space. Micropropagation is a low-cost technology, and plantlet derived from micropropagation helps in plants germplasm conservation (Gupta et al., 2020). In this study, micropropagation method was developed for two common varieties of taro in Kenya, that is, Dasheen and Purple Wild.
Benzylaminopurine (BAP), an adenine derivative, is an important cytokinin used in shoot induction (Chand et al., 1999). BAP hormone is mainly preferred for in-vitro regeneration of monocots (Ramakrishnan et al., 2014). Results generated from shoot proliferation media showed a significant difference in the number of shoots induction between the two taro varieties. Reports by Toledo et al. (1998) state that various varieties of potatoes respond to shoot induction differently because of different genetic background. In this study, shoots and leaves were best produced on the two varieties when cultured on a BAP level of 2 mg/L. Here, we noted that this regime gave the best shoot induction in both Dasheen and Purple Wild varieties. BAP enhanced growth and development of axillary buds in C. esculenta var. esculenta, and this has been documented in a previous study (Chung and Goh, 1994). Seetohul et al. (2008) report the highest in-vitro multiplication rates of taro shoot tips in MS medium supplemented with 2 mg/L of BAP, which is agreeing with our findings. As observed in the current study, a higher concentration of BAP (6 mg/L) produced a fewer shoot compared to a lower concentration (El-Sayed et al., 2016). This could be attributed to the toxicity caused by high cytokinin concentrations, which causes a delay in shoot formation (Manju et al., 2017). During the multiplication stage, the requirement of cytokinin differs depending on the type of crop, explant, developmental phase, growth regulator concentration, and the interaction between growth regulators and the environment (Yokoya et al., 1999).
Both taro varieties used in this study successfully formed root when grown in MS media supplemented with different concentrations of IBA as well as in the absence of IBA. Roots are essential to plants as they supply water and nutrients to plants (Schiefelbein et al., 1997). An earlier study reports better rooting in taro shoots in half-strength MS medium supplemented with 1.5 or 2.0 mg/L of naphthaleneacetic acid (NAA) than in half-strength MS medium supplemented with 1.5 or 2.0 mg/L IBA at 15 days of culture (Behera and Sahoo, 2008). The present study showed that MS (4.4 g/L) premix supplemented with 0.5 mg/L of IBA was the best concentration for rooting in both taro varieties evaluated. The application of auxins to micro-propagated shoots could intensify the production of the root by increasing the endogenous contents of enzymes (Asghar et al., 2011). Another study reports auxins induced complication in lateral root formation through repetitive cell division (Liu et al., 2002). In the present study, the roots were of a shorter length in IBA supplemented medium than those induced in media with no IBA, possibly due to inhibition of shoot bud formation at elevated IBA concentration and arrest of root production as the auxin in the root primordial is shifted from the apex shoot (Ozel et al., 2006).
A previous study established that plants could be transplanted when they have rooted, but good results are achieved if the plants do not have too many long roots but have a greater number of roots (Singh et al., 2012). Despite initial difference in root length all the plants developed well during the acclimatization. The present study demonstrates that supplementation of MS media (4.4 g/L) with 2.0 mg/L BAP and 0.5 mg/L IBA enhance better shoot and root induction, respectively.
This study has developed an efficient regeneration protocol for Dasheen and Purple Wild, two common taro varieties in Kenya. The upper part of the corm and the base of the petioles were used as the explants. This protocol is a doorway for future studies on taro in tissue culture and facilitates rapid multiplication of disease-free genetically uniform taro varieties for commercial uses. The protocol can also be useful in the taro transformation process, a means for rapid multiplication of genetically engineered plants improved for different traits of agronomic importance.
MS, Murashige and Skoog media; BAP, 6-benzylaminopurine; IBA, indole-3-butyric acid.
The authors have not declared any conflict of interests.
The authors, with much appreciation, acknowledge the support of BecA-ILRI Hub through the Africa Biosciences Challenge Fund (ABCF) program, which is funded by the Australian Department for Foreign Affairs and Trade (DFAT) through the BecA-CSIRO partnership; the Syngenta Foundation for Sustainable Agriculture (SFSA); the Bill & Melinda Gates Foundation (BMGF); the UK Department for International Development (DFID) and the Swedish International Development Cooperation Agency (Sida). The authors appreciate the support of the International Institute of Tropical Agriculture (IITA) and the farmers in Busia, Kakamega, Kiambu, Kisii, Machakos, Meru, Murang’a, Nyeri, and Siaya who provided them with the germplasm.
REFERENCES
|
Addinsoft (2021). XLSTAT statistical and data analysis solution. New York, USA.
View
|
|
|
|
Asghar S, Ahmad T, Hafiz IA, Yaseen M (2011). In vitro propagation of orchid (Dendrobium nobile) var.Emma white. African Journal of Biotechnology 10(16):3097-3103.
Crossref
|
|
|
|
|
Behera KK, Sahoo S (2008). In vitro micropropagation of Colocasia esculenta (L.) Schott. (cv local-jhankhri) through corm sprouts. The Orissa Journal of Horticulture 36:50-54.
|
|
|
|
|
Boampong R, Aboagye LM, Nyadanu D, Essilfie ME, Adu Amoah R (2019). Biochemical characterization of some Taro (Colocasia esculenta L. Schott) Germplasm in Ghana. Advances in Agriculture.
Crossref
|
|
|
|
|
Burner DM, Grisham MP (1995). Induction and stability of phenotypic variation in sugarcane as affected by propagation procedure. Crop Science 35(3):875-880.
Crossref
|
|
|
|
|
Bhojwani SS, Dantu PK (2013). Micropropagation. In: Plant Tissue Culture: An Introductory Text. Springer, India.
Crossref
|
|
|
|
|
Chand H, Pearson MN, Lovell PH (1999). Rapid vegetative multiplication in Colocasia esculenta (L.) Schott (Taro). Plant Cell, Tissue and Organ Culture 55(3):223-226.
Crossref
|
|
|
|
|
Chung RC, Goh CJ (1994). High frequency direct shoot regeneration from corm axillary buds and rapid clonal propagation of Taro, Colocasia esculenta var. esculenta (L.) Schott (Araceae). Plant Science 104(1):93-100.
Crossref
|
|
|
|
|
El-sayed SE, Gharib AA, El-Sawy AM, Omaima SD (2016). Micropropagation protocol of Egyptian native cultivar of Taro, Colocasia esculenta var. esculenta. International Journal of Advanced Research in Biological Sciences 3(1):17-26.
|
|
|
|
|
Fay MF (1992). Conservation of rare and endangered plants using In vitro methods. In Vitro Cellular and Developmental Biology-Plant 28(1):1-4.
Crossref
|
|
|
|
|
Food and Agriculture Organization of the United Nations (2021). FAOSTAT statistical database. Rome: FAO.
|
|
|
|
|
Gupta N, Jain V, Rosy M, Dev S (2020). A Review on Micropropagation Culture Method, Asian Journal of Pharmaceutical Research and Development 8(1):86-93.
Crossref
|
|
|
|
|
Hossain MJ (2012). In vitro organogenesis of Colocasia esculenta cv. Antiquorum L. American Journal of Plant Sciences 3:709-713.
Crossref
|
|
|
|
|
International Atomic Energy Agency (IAEA) (2004). Low cost options fortissue culturetechnology in developing countries. Proceedings of a Technical Meeting organized by theJoint FAO/IAEA Division of Nuclear Techniques in Food and Agriculture, 26-30 August 2002, Vienna. IAEA-TECDOC-1384,.102p.
|
|
|
|
|
Jackson GV, Vilsoni F, Kumar J, Pearson MN, Morton JR (2001). Comparison of the growth of In vitro produced pathogen-tested Colocasia taro and field-collected planting material. New Zealand Journal of Crop and Horticultural Science 29:171-176.
Crossref
|
|
|
|
|
Jane J, Shen L, Lim S, Kasemsuwantt T, Nip K (1992). Physical and chemical studies of taro starches and flours. Cereal Chemistry 69(5):528-535.
|
|
|
|
|
Kantaka S (2004). Colocasia esculenta (L.). Schott. Grubbrn GJH, Denton OA (Eds). PROTA (Plant resources of Tropical Africa / Ressources vegetales de l' Afrique tropicale). Netherlands, Wageningen.
|
|
|
|
|
Kidanemariam DB (2018). Viruses of taro and other edible aroids in east Africa (Doctoral dissertation, Queensland University of Technology).
|
|
|
|
|
Lebot V (2009). Tropical root and tuber crops: cassava, sweet potato, yams and aroids. UK, MPG Biddles Ltd.
|
|
|
|
|
Liu C, Zhu J, Liu Z, Li L, Pan R, Jin L (2002). Exogenous auxin effects on growth and phenotype of normal and hairy roots of Pueraria lobata (Wild.) Ohwi. Plant Growth Regulation 38:37-43.
Crossref
|
|
|
|
|
Manju B, Evelyn, Mbong AG, Fokunang N, Charles Tembe-Fokunang, Estella A, Rachid H (2017). Application of In vitro micropropagation technique for sustainable production of four local taro cultivars in Cameroon. African Journal of Biotechnology 16(30):1638-1645.
Crossref
|
|
|
|
|
Matthews PJ (1991). A possible tropical wild type taro: Colocasia esculenta var. equatilis. Indo-Pacific Prehistory Association Bulletin 11:69-81.
Crossref
|
|
|
|
|
Minas GJ (2002). In vitro micropropagation of kolokasi from apical meristem. Agricultural Research Institute, Nicosia, Cyprus. Miscellaneous Reports 84. 8 p.
|
|
|
|
|
Mukami A, Ngetich A, Mweu C, Muthangya M, Oduor RO, Ngugi M, Mbinda W (2018). "Rapid and Efficient Plant Regeneration from Shoot Apical Meristems of Finger Millet [ Eleusine Coracana (L .) Gaertn]. African Journal of Biotechnology 17(29):898-905.
Crossref
|
|
|
|
|
Ngetich A, Runo S, Omwoyo O, Ngugi M (2015). "Low-Cost Micropropagation of Local Varieties of Taro (Colocasia esculenta Spp .)". Biotechnology Journal International pp. 136-145.
Crossref
|
|
|
|
|
Okonkwo CAC (1993). Taro: Colocasia spp. In: G Kalloo, BO Bergh (eds.) Genetic Improvement of Vegetable Crops pp. 709-715. Pergamon, Amsterdam.
Crossref
|
|
|
|
|
Onwueme IC (1999). Taro cultivation in Asia and the Pacific. Bangkok, Thailand, Food and Agriculture Organization of the United Nations Regional Office for Asia and the Pacific.
|
|
|
|
|
Ozel CA, Khawar KH, Mirici S, Arslan O, Sebahattin O (2006). Induction of Ex Vitro Adventitious Roots on Soft Wood Cuttings of Centaurea tchihatcheffii tchihatcheffii Fisch et. Mey using Indole 3Butyric Acid and α-Naphthalene Acetic Acid. International Journal of Agriculture and Biology 8(1):66-69.
|
|
|
|
|
Ramakrishnan M, Ceasar SA, Duraipandiyan V, Ignacimuthu S (2014). Efficient plant regeneration from shoot apex explants of maize (Zea mays) and analysis of genetic fidelity of regenerated plants by ISSR markers. Plant Cell, Tissue, and Organ Culture 119(1):183-196.
Crossref
|
|
|
|
|
Sarasan V, Cripps R, Ramsay MM, Atherton C, McMichen M, Prendergast G, Rowntree JK (2006). Conservation in vitro of threatened plants -Progress in the past decade. In Vitro Cellular and Developmental Biology-Plant 42(3):206-214.
Crossref
|
|
|
|
|
Schiefelbein JW, Masucci JD, Wang H (1997). Building a root: The control of patterning and morphogenesis during root development. Plant Cell 9(7):1089-1098.
Crossref
|
|
|
|
|
Seetohul S, Puchooa D, Ranghoo-Sanmukhiya VM (2008). Genetic improvement of Taro (Colocasia esculenta var esculenta) through In vitro mutagenesis. University of Mauritius Research Journal 13:79-89.
|
|
|
|
|
Singh D, Jackson G, Hunter D, Fullerton R,Lebot V, Taylor M, Iosefa T, Okpul T, Tyson J (2012). "Taro Leaf Blight-A Threat to Food Security. Agriculture 2(4):182-203.
Crossref
|
|
|
|
|
Temesgen M, Retta N (2015). Nutritional potential, health and food security benefits of taro Colocasia esculenta (L.): A review. Food Science and Quality Management 36:23-30.
|
|
|
|
|
Toledo J, Espinoza N, Golmirzaie A (1998). Tissue culture management of in-vitro plantlets in potato seed production. Training Manual. International Potato Centre.
|
|
|
|
|
Tuia VS (1997). In vitro multiplication of taro (Colocasia esculenta var.esculenta L. Schott), M.Agric. Thesis, University of the South Pacific, Fiji.
|
|
|
|
|
Villalobos VM, Engelmann F (1995). Ex situ conservation of plant germplasm using biotechnology. World Journal of Microbiology and Biotechnology 11:375-382.
Crossref
|
|
|
|
|
Wang QC, Valkonen JP (2008). Efficient elimination of sweetpotato little leaf phytoplasma from sweetpotato by cryotherapy of shoot tips. Plant Pathology 57(2):338-347.
Crossref
|
|
|
|
|
Wang QC, Valkonen JP (2009). Cryotherapy of shoot tips: Novel pathogen eradication method. Trends in Plant Science 14(3):199-122.
Crossref
|
|
|
|
|
Wickham H (2016). ggplot2: Elegant Graphics for Data Analysis: Springer-Verlag New York.
Crossref
|
|
|
|
|
Yam TW, Hsu GI, Arditti J (1990). Plant regeneration in vitro of South Pacific taro (Colocasia esculenta var. esculenta cv. Akalomamale, Aracea) Plant Cell Reports 9:229-232.
Crossref
|
|
|
|
|
Yokoya NS, Hirotaka K, Hideki O, Takao K (1999). Effects of environmental factors and plant growth regulators on growth of the red alga Gracilaria vermiculophylla from Shikoku Island, Japan. Hydrobiologia 398:339-347.
Crossref
|
|
|
|
|
Yoshino H (2002). Morphological and genetic variation in cultivated and wild taro. In S Yoshida, PJ Matthews (Eds.), Vegeculture in east-ern Asia and Oceania (pp. 95-116). JCAS Symposium Series 16. Japan Center for Area Studies.
|
|