Full Length Research Paper

ABSTRACT
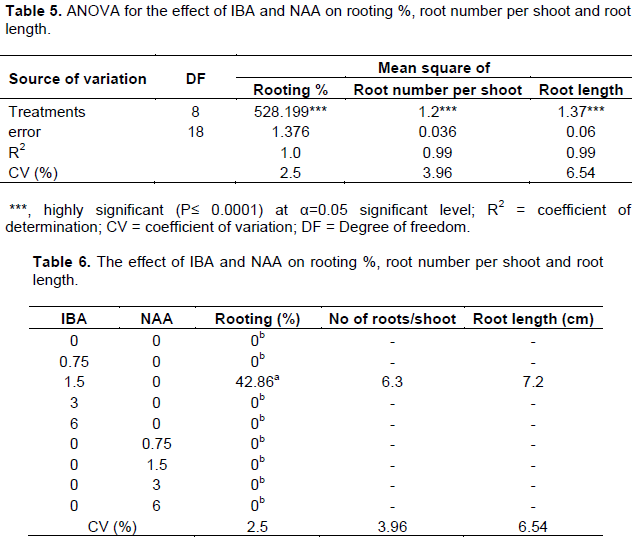
The present study aimed to develop micropropagation procedure for Prunus persica cv. Garnem. Shoot induction and multiplication was found from nodal explants cultured on MS medium fortified with different concentrations of two cytokinins (6-benzyladeninepourine and Kinetin) alone and in combination with 0.5 mg/l gibberellic acid and 0.01 mg/l indole-3-butyric acid. Microplantlets were cultured to root on ½ strength MS medium supplemented with different concentrations of indole-3-butyric acid and α-naphthalene-acetic acid alone. 6-Benzyladeninepourine (0.5 mg/l) with indole-3-butyric acid (0.01 mg/l) and gibberellic acid (0.5 mg/l) was found to be the optimum combination treatment for shoot initiation (100%). 6-Benzyladeninepourine (2.0 mg/l) with indole-3-butyric acid (0.01 mg/l) and gibberellic acid (0.5 mg/l) was found to be optimum for maximum shoot number per explant (7.67). The maximum rooting (42.86%), maximum root number/shoot (6.33) and longest rooting (7.17 cm) were found at 1.5 mg/l indole-3-butyric acid. α-naphthalene-acetic acid was found to be not effective. The plantlets were acclimatized in the glasshouse and survival percentage was 73.3% at potting mixture of autoclaved river sand, forest soil and manure in a 2:1:1 (v/v/v) ratio.
Key words: Acclimatization, auxin, cytokinin, micropropagation, Prunus persica.
INTRODUCTION
MATERIALS AND METHODS
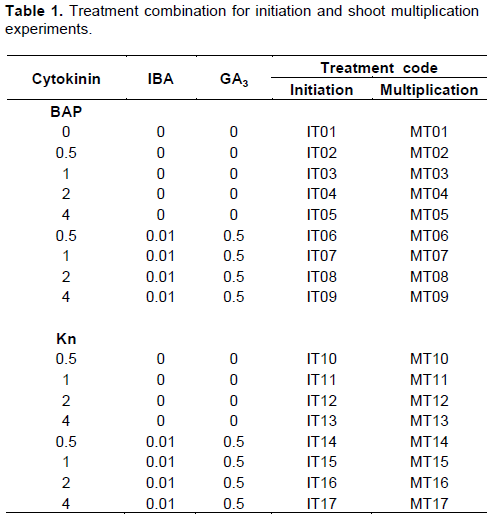
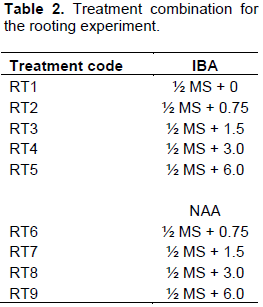
RESULTS AND DISCUSSION
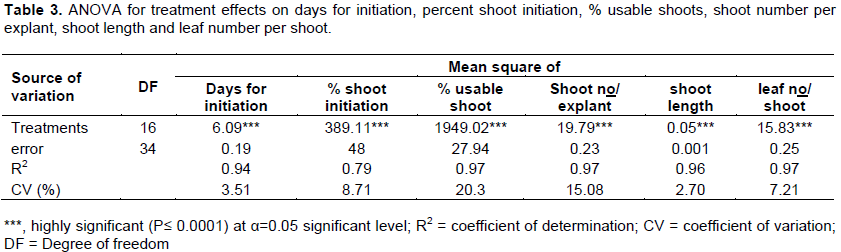
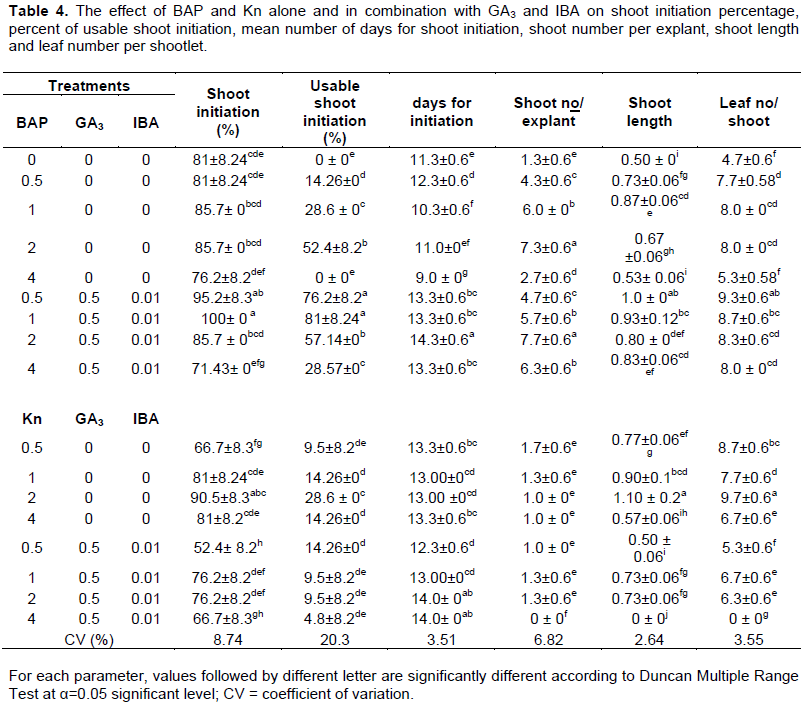


CONFLICT OF INTERESTS
REFERENCES
|
Ainsley PJ, Collins GG, Sedgley M (2000) Adventitious shoot regeneration from leaf explants of almond (Prunus dulcis Mill.). In vitro Cell. Dev. Biol-Plant 36:470-474. |
|
|
Alanagh EN, Garoosi GA, Haddad R (2010). The effect of PGRs on in vitro shoot multiplication of GF677 hybrid (Prunus persica x P. amygdalus) rootstock on GNH medium. Iranian J. Genet. Plant Breeding 1(1):34-43. |
|
|
Ammer M (1999). Performance of Hansen, GF 655 and GF677 peach rootstocks for rooting with the use of IBA under greenhouse condition. M.Sc. thesis. Univ. Arid. Agri. Rawalpindi, Pakistan, P 65. |
|
|
Ansar A, Touqeer A, Nadeem AA, Ishfaq, AH (2009). Effect of different concentrations of auxins on in vitro rooting of olive cultivar 'Moraiolo'. Pak. J. Bot. 41(3):1223-1231. |
|
|
Aranzana MJ, Abbassi El-K, Howad W, Arus P (2010). Genetic variation, population structure and linkage disequilibrium peach commercial varieties. Bio-Med. Cen. Genet. 11(69):1-12. |
|
|
Benelli C, Fabbri A, Grassi S, Lambardi M, Rugini E (2001). Histology of somatic embryogenesis in mature tissues of olive (Olea europaea L.). J. Hortic. Sci. Biotechnol. 76(1):112-119. |
|
|
Deepa VS, Rajaram K, Kumar MA, Das S, Kumar PS (2011). High frequency regeneration and shoot multiplication in Andrographis lineate wall. ex. nees: an endemic medicinal plant of south India. J. Med. Plant Res. 5(20):5044-5049. |
|
|
Dejampour J, Grigourian V, Majidi E, Asgharzade N (2007). Sterilization, establishment and proliferation of some Prunus interspecific hybrids for in vitro culture. J. Hortic. Sci. Technol. 8:165 - 174. |
|
|
Demsachew G (2011). Micropropagation of two apple (Malus domestica Borkh) varieties from shoot tip explants. An M.Sc Thesis presented to the school of Graduate Studies of Haramaya University, Haramaya, Ethiopia. P 60. |
|
|
Felipe AJ (2009). 'Felinem', Garnem' and 'Monegro' almond x peach hybrid rootstocks. Hortic. Sci. 44:196-197. |
|
|
Fotopoulos S, Sotiropoulos TE (2005). In vitro rooting of PR 204/84 rootstock (Prunus persica x P. amygdalus) as influenced by mineral concentration of the culture medium and exposure to darkness for a period. Agron. Res. 3(1):3-8. |
|
|
Gentile A, Monticelli S, and Damiano C (2002). Adventitous shoot regeneration in peach (Prunus persica (L.) Batsch). Plant Cell Rep. 20:1011-1016. |
|
|
Gomez KA, Gomez AA (1984). Statistical Procedure for Agricultural Research. New York, John Wiley and Sons. |
|
|
Hammatt N, Grant NJ (1997). Micropropagation of mature British wild cherry. Plant Cell Tissue, Organ. Cult. 47:103-110. |
|
|
Hammatt N, Grant NJ (1998). Shoot regeneration from leaves of Prunus serotina Ehrh. (black cherry) and P.avium (wild cherry). Plant Cell Rep. 17:526-530. |
|
|
Hesse CO (1975). Peaches. In: Janick J, Moore J (eds.). Advances in fruit breeding. Purdue Univ. Press, West Lafayette, Ind. Pp.285-335. |
|
|
Jimenez S, Pinochet J, Romero J, Gogorcena Y, Moreno MA, Espada JL (2011). Performance of peach and plum based rootstocks of different vigour on a late peach cultivar in replant and calcareous conditions. Sci. Hortic. 129(1):58-63. |
|
|
Kalinina A, Brown DCW (2007). Micropropagation of ornamental Prunus spp. and GF305 peach, a Prunus viral indicator. Plant Cell Rep. 26:927-935. |
|
|
Krieken WVD, Breteler H, Visser MHM and Mavridou D (1993). The role of the conversion of IBA into IAA on root regeneration in apple. Introduction of test system. Plant Cell Rep. 12:203-206. |
|
|
Krussmann G (1986). Manual of cultivated broad leaved trees and shrubs. Vol. 3. Timber Press, Portland, Oregon. pp. 18-58. |
|
|
Mansseri-Lamrioui A, Louerguioui A, Bonaly J, Yakoub-Bougdal S, Allili N, Gana-Kebbouche S (2011). Proliferation and rooting of wild cherry: the influence of cytokinin and auxin types and their concentration. Afr. J. Biotechnol. 10(43):8613-8624. |
|
|
Martinez-Gomez P, Sanchez-Perez R, Rubiio M, Dicenta F, Gradziel T M, Sozzi GO (2005). Application of recent biotechnologies to Prunus tree crop genetic improvement. Clen. Inv. Agric. 32(2):73-96. |
|
|
Muna AS, Ahmad AK, Mahmoud K, Abdul-Rahman K (1999). In vitro propagation of a semi-dwarfing cherry rootstock. Plant Cell Tiss. Org. Cult. 59:203-208. |
|
|
Murashige CT, Skoog F (1962). A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 15:473-497. |
|
|
Nagaty MA (2012). Establishment of regeneration system for Taif peach (Prunus persica L. Batsch) cultivar (Balady cultivar) in Taif, KSA. J. Am. Sci. 8(4):232-239. |
|
|
Naghmouchi S, Khouja ML, Rejeb MN, Boussaid M (2008). Effect of growth regulators and explant origin on in vitro propagation of Ceratonia siliqua L. via cuttings. Biotechnol. Agron. Soc. Environ. 12(3):251-258. |
|
|
Ndoye M, Diallo I, Gassama YK (2003). In vitro multiplication of the semi-arid forest tree, Balanites aegyptiaca (L.) Del. Afr. J. Biotechnol. 2(11):421-424. |
|
|
Nissen SJ, Sutter EG (1990). Stability of IAA and IBA in nutrient medium of several tissue culture procedures. Hort. Sci. 25(7):800-802. |
|
|
Perez-Tornero O, Egea J, Vanoostende A, Burgos L (2000). Assessment of factors affecting adventitious shoot regeneration from in vitro cultured leaves of apricot. Plant Sci. 158:61-70. |
|
|
Pinochet J, Calvet C, Dorrego AH, Bonet A, Felipe A, Moreno M (1999). Resistance of peach and plum rootstocks from Spain, France, and Italy to root-knot nematode Meloidogyne javanica. Hort. Sci. 34(7): 1259-1262. |
|
|
Pruski KW, Lewis T, Astatkie T, Nowak J (2000). Micropropagation of Chokecherry and Pincherry cultivars. Plant Cell Tissue Organ. Cult. 63:93-100. |
|
|
Ramage CM, Williams RR (2004). Cytokinin-induced abnormal shoot organogenesis is associated with elevated knotted 1-type homeobox gene expression in tobacco. Plant Cell Rep. 22:919-924. |
|
|
Reighard G (2011). Peach rootstocks. NC-140 Regional Rootstock Research Project. Utah State Hort Association, Clemson University. |
|
|
Scorza R., Sherman WB (1996). Peachs. In: Janick J, Moore JN, (eds.). Fruit breeding: tree and tropical fruits 1. Wiley, New York, NY, USA. pp. 325-440. |
|
|
Smulders MJM, Van De ETWM, Croes AF, Wullems GJ (1990). Metabolism of 1-naphthaleneacetic acid in explants of tobacco: evidence for release of free hormone from conjugates. J. Plant Growth Regul. 9:27-34. |
|
|
Stylianides DC, Tsipouridis C, Michailidis Z (1989). Resistance to iron deficiency of five peach rootstocks. International Peach Symposium. Acta Hortic. 254:185-188. |
|
|
Subbu RR, Chandraprabha A, Sevugaperumal R (2008). In vitro clonal propagation of vulnerable medicinal plant, Saraca asoca (Roxb.) De Wilde. Nat. Prod. Rad. 7(4):338-341. |
|
|
Tanimoto E (2005). Regulation of root growth by plant hormones: Roles for auxin and gibberellin. Crit. Rev. Plant Sci. 24(4):249-265. |
|
|
Tiwari SK, Tiwari KP, Siril EA (2002). An improved micropropagation protocol for teak. Plant Cell Tissue Organ Cult. 71:1-6. |
|
|
Tsipouridis C, Thomidis T (2003). Methods to improve the in vitro culture of GF677 (peach x almond) peach rootstock. New Zealand J. Crop Hort. Sci. 31:361-364. |
|
|
Vaghari-Azar E, Vatapour-Azghandi A, Majidi-Heravan E, Dejampour J, Habashi AA (2012). Micropropagation of two apricot x plum inter-specific hybrid rootstocks (HS405 and HS706). Iranian J. Genet. Plant Breeding 1(2):9-15. |
|
|
Van Bragt J, Pierik RLM (1971). The effect of autoclaving on the GA3. In Van Bragt J, Mossel DAA, Pierik RLM, Veldstra H (Eds.), Effects of sterilization on components in nutrient media, Wageningen, The Netherlands: Kniphorts Scientific. pp. 133-137. |
|
|
Zaerr JB, Mapes MO (1982). Action of growth regulatorsIn: Bonga JM, Durzan DJ (Eds.). Tissue Culture in Forestry. Martinus Nijhoff/Dr. W. Junk Publishers, The Hague/Boston/London. pp. 231-255. |
|
|
Zarrouk O, Gogorcena Y, Gomez-Aparisi J, Betran JA, Moreno MA (2005). Influence of almond x peach hybrids rootstocks on flower and leaf mineral concentration, yield and vigour of two peach cultivars. Hortic. Sci. 106(4):502-514. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


