Full Length Research Paper

ABSTRACT
To evaluate the effect of arbuscular mycorrhizal fungi and phosphorus levels on root traits of cucumber plants, a factorial experiment was carried out based on a randomized completely design pot culture. Four phosphorus fertilization treatments, including 2, 5, 10 and 15 mg P kg-1 soil possessed phosphorus fertilization levels as the first factor. At the second factor arranged Glomus mosseae, Glomus intraradices of mycorrhiza species and non-inoculum as a control with three replications were conducted in the greenhouse of agricultural research center of west Azarbaijan province Urmia, in 2013. Results show that above-ground dry matter of inoculated cucumber at both species with 155.00 and 160.83 mg/plant had the highest values. Both species had more root fresh and dry weight, root length and root volume than control. Colonization of G. mosseae and G. intraradices, with 53.20 and 44.59% had the highest values at the 2 mg P kg-1 soil. G. mosseae and G. intraradices had the highest leaf phosphorus with 486.06 and 477.60 mg/100 g of leaf dry weight at the 15 mg P kg-1 soil, respectively. Leaf phosphorus (r = 0.62**), root dry weight (r = 0.79**), root length (r=0.44**), root volume (r = 0.82**) and fresh root weight (r = 0.74**) had positive correlation coefficients with above-ground dry matter. Although application of phosphorus increased above-ground dry matter and root traits, but our study clearly demonstrates that mycorrhizal fungi play an important role in the enhancement of growth of cucumber plants under very low phosphorus conditions.
Key words: Colonization, cucumber, dry matter, insoluble phosphorus, mycorrhiza.
INTRODUCTION
Phosphorus is critical for plant growth, and is a component of the nucleic acid structure of plants and bio-membranes. Therefore, it is important in cell division and tissue development. Phosphorus is also involved in the energy metabolism of cells and is required for the biosynthesis of primary and secondary metabolites in plants. Consequently, plants have evolved a range of strategies to increase phosphorus uptake and mobility (Marschner, 1996), the most common among which is arbuscular mycorrhiza (AM) symbiosis. In AM fungi symbiosis with plant roots, the enhanced uptake of phosphorus is attributed to the fungal partner, and the increase in phosphorus uptake by the colonized roots in turn leads to increased plant growth (Burleigh et al., 2002).
AM fungi are one of the most widespread mycorrhizal associations between soil microorganisms and higher plants. The function of all mycorrhizal systems depends on the ability of the fungal symbiont to absorb inorganic and organic nutrients available in soil (Marschner and Dell, 1994). The AM fungi infect the roots of a susceptible plant, forming a mutually beneficial, symbiotic relationship. In exchange for carbohydrates from the host plant, AM fungi benefit the plant primarily through increased uptake of soil nutrients, such as phosphorus, zinc and copper (Miller et al., 1986). This enhanced mineral uptake is facilitated by external hyphae of the fungi, which exploit a greater volume of soil than roots, thus accessing nutrients (especially phosphorus) not normally accessible to the plant's root system (Hayman, 1983). Colonization of Cucumis sativus by AM fungi affects flowering, fruit production, photosynthesis rates, and disease resistance (Trimble and Knowles, 1995; Valentine et al., 2001; Hao et al., 2005; Kiers et al., 2010).
AM fungi are important due to their great capability to increase plant growth and yield under certain conditions. The major reason for this increase is the ability of plants in association with AM to take some nutrients such as phosphorus efficiently (Podila and Douds, 2001). AM fungi, as obligate symbionts, also depend for their growth and activity on the supply of carbon compounds from the photosynthetic partner (Ocampo and Azcon, 1985; Jennings, 1995). AM symbiosis can cause an important carbohydrate gain in the host plant and up to 20% of total photo-assimilate substances can be transferred to the fungal partner (Graham, 2000). Inoculation with AM fungi, in some vegetables, may improve growth performance (Temperini et al., 2009). The benefits of AM fungi inoculation depend on the genotypic host-fungus combinations and also the type of the inoculums used (Rouphael et al., 2010). The aim of this study was to evaluate the effect of AM fungi species, Glomus mosseae and Glomus intraradices, on dry matter production and root traits of cucumber plants under different levels of phosphorus.
MATERIALS AND METHODS
A experiment was conducted at greenhouse with a day/night cycle of 16 h at 22°C and 8 h at 19°C (relative humidity: 50 to 70%) in the agricultural research center of west Azarbaijan province, Urmi, Iran. The greenhouse located in longitude 37°, 35', 32'' north, latitude 45°, 3′, 39'' east and 1330m altitude. Some physicochemical properties of soil which is used to growth the cucumber plants were determined (Table 1). Soil was collected from a low P (2 ppm Olsen extractable P) field in the region of Shaharchay River around of Urmia.
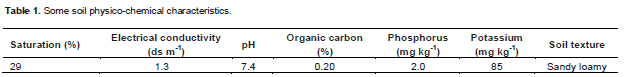
A factorial experiment based on a randomized completely design was carried out with three replications. Four phosphorus fertilization treatments in this study, including 2, 5, 10 and 15 mg P kg-1 soil (KH2PO4) were incorporated into the soil by hand and mycorrihzal treatments, a control with no inoculums and two mycorrhizal fungi inoculums, G. mosseae and G. intraradices arranged as the first and second factors, respectively.
Seeds of the cucumber cultivar were provided by the Agricultural Research Station of Urmia. The two species of AM fungi used in this study were G. mosseae and G. intraradices, which were produced on maize (Zea mays L.) host plants by Dr. E.M. Goltapeh at Tarbiat Modarres University, Tehran, Iran. The mycorrhizal inoculum was a mixture of sterile sand, mycorrihzal hyphae, spores (25 spores g–1 inoculums), and colonized root fragments. Seeds of cucumber were surface sterilized with 0.05% sodium hypo-chloride for 45 min before sowing them. Seeds were sown in sterilized soils in plastic pots (12 ×12 cm) at a depth of 3 cm on 25 June 2013. Thirty grams of the appropriate inoculums was placed into the hole below each seed, and then covered with sterile soil. For non-mycorrhizal control plants were sown with no inoculation. The plants were grown in a greenhouse under natural photoperiods for 6 weeks during which only distilled water was applied. In addition, twice a week, each pot was supplied with 100 ml of a nutrient solution containing 720 mg of MgSO4.7H2O, 295 mg of Ca (NO3)2.4H2O, 240 mg of KNO3, 0.75 mg of MnCl2.4H20, 0.75 mg of KI, 0.75 mg of ZnSO4.7H2O, 1.5 mg of H3BO3, 0.001 mg of CuSO4.5H20, 4.3 mg of FeNaEDTA and 0.00017 mg of Na2MoO4.2H2O supplemented without phosphorus (Vosatka and Gryndler, 1999).
The root fresh weights were measured before drying at 72°C for 24 h that leads to the weights of the dry matter for root. Length and volume of roots, root dry weights and above-ground dry matter of seedlings were determined after harvesting. At 6 weeks after planting, the percentage of colonization of cucumber roots by AM fungi was determined per experimental unit. Root colonization was measured in fresh roots cleared in 10% KOH for 10 min at 90°C and stained in 0.05% lactic acid-glycerol-Trypan Blue (Phillips and Hayman, 1970). The percentage of root colonization by AM fungi was microscopically determined using the gridline intersection method (Giovannetti and Mosse, 1980). To measure leaf phosphorus, dried leaves were milled, digested, and analyzed as described by Watanabe and Olsen (1965) and Ohnishi et al. (1975). The method described for phosphorus involves drying, homogenization, and combustion (4 h at 500°C) of the leaf sample. The plant ashes (5 mg) are digested in 1 ml of concentrated HCl. The samples are then filtered, and total phosphorus is quantified as PO4– using the ascorbic acid method (Watanabe and Olsen, 1965). The amount of PO4– in solution was determined calorimetrically at 882 nm (Graca et al., 2005).
Analysis variance of data was performed using MSTATC software. The effects of phosphorus, application of mycorrhizae, and the interactions of these two factors were analyzed by ANOVA and the means compared by the Duncan’s Multiple Range test (P ≤ 0.05). Also, correlation coefficients were calculated.
RESULTS AND DISCUSSION
Different levels of phosphorus and mycorrhizae for traits of root fresh weight, root dry weight, root volume, total dry weight and interaction between them for leaf phosphorous accumulation, root length and mycorrhiza colonization had significant differences (Table 2).

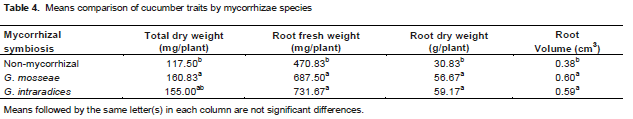
.png)
Colonization percentage of G. mosseae was more than G. intraradices and was less reduced with increasing phosphorus levels. Variations of this trait were for G. mosseae between 29.34 to 53.20 and G. intraradices 23.97 to 44.59. Colonization mycorrhiza was reduced due to increasing phosphorus fertilization (Table 3). Both species of mycorrhiza had more root fresh weight, root dry weight and root volume with 731.67 mg, 59.17 mg, 0.59 cm3, respectively than non-inoculated cucumber plants (Table 4). Root dry weight and root volume increased with improved phosphorus fertilization. Phosphorus fertilization treatments of 2 and 15 mg kg-1 soil were 36.11 mg and 0.42 cm3 and 65.56 mg and 0.64 cm3 values of them, respectively (Table 5). Expanded roots of mychorrhizal plants enhanced root area (Allen et al., 1981). Therefore, water and nutrient uptake in mycorrhizal plants was due to more root expansion than control (Huang et al., 1985).

Analysis of the phosphorus accumulation in leaves of the cucumber plants showed that the highest phosphorus accumulation in leaves (486.06 mg/100 g dry leaf) was obtained from the plants inoculated with G. mosseae and phosphorus treatment 15 Mg P kg-1 soil (Table 3). The minimum phosphorus accumulation in leaves (392.36 mg/100 g dry leaf) was obtained from the non-mycorrhizal and 2 mg P kg-1 soil, followed by the non-mycorrhizal plants 5 and 10 mg P kg-1 soil. The phosphorus concentration in the leaves of cucumber plants in each treatment was significantly higher than that in the control. Plants were more responsive to additional phosphorus in the low to medium phosphorus (2 to 15 mg phosphorus kg-1 soil) range, while AM infected plants were more responsive in the low phosphorus (2 and 5 mg phosphorus kg-1 soil) range, with increasing colonization and acquiring phosphorus (Table 3). The significance of sufficient phosphorus availability during early crop growth has been reported in different crop species (Grant et al., 2005). It has been reported that enhanced early-season phosphorus nutrition in maize increased dry matter at early stages partitioned to the grain at later development stages (Parewa et al., 2010). Likewise, in wheat and barley, phosphorus supply during earlier growth had superior effect on final grain yield than phosphorus supply in later stages (Smith and Smith, 2011).
Plenets et al. (2000) reported a greater difference in dry matter accumulation of maize under phosphorus deficiency during early stages of growth. The above ground dry matter accumulation was observed to be severely reduced (up to 60%) during early stages of maize growth, while there were only slight differences on dry matter accumulation at harvest and grain yield. The effect of early phosphorus deficiency on decline in shoot growth occurs because of slight stimulation of root growth (Mollier and Pellerin, 1999). The initial reduction in growth related to phosphorus deficiency has an ultimate effect on the final crop yield, which is experienced by the crop throughout the remaining of the growing period. Phosphorus is critical for plant growth and makes up about 0.2% of dry mass, but it is one of the most difficult nutrients for plants to acquire. In soil, it may be present in relatively large amounts, but much of it is poorly available because of the very low solubility of phosphates of iron, aluminum, and calcium, leading to soil solution concentrations of 10 mM or less and very low mobility (Ryan et al., 2005). The ability of AM fungi to enhance host-plant uptake of relatively immobile nutrients, in particular phosphorus and Zn (Balakrishnan and Subramanian, 2012), and their requirement for up to 20% of host-plant for establishment and maintenance, is well accepted (Subramanian et al., 2009).
Analysis of the root length of cucumber plants showed that the highest root length (20.33 cm) was obtained from the plants inoculated with G. intraradices and phosphorus treatment 10 mg P kg-1 soil (Table 3). The minimum root length (11.67 and 10.67 cm) was obtained from the non-mycorrhizal and 5, 10 mg P kg-1 soil (Table 3). Marulanda et al. (2007) reported that in lavender inoculated plants with G. mosseae and G. intraradices improved root growth 35 and 100%, respectively.
Between different levels of phosphorus application, 15 mg P kg-1 soil had the most above-ground dry matter with 187.78 mg/plant and the lowest phosphorus application (2, 5 mg P kg-1 soil) had above-ground dry matter with 114.44 and 131.11 mg/plant. Both Species with 160.83 and 155.00 mg/plant above-ground dry matters had the highest values than control (Tables 4 and 5). Subramanian et al. (2006) observed that root colonization by the AM fungus significantly increased dry matter yield and ultimate increased the production. Total dry weight differences in mycorrhizal treatments are related to water absorption and mineral nutrients (AL-Karaki et al., 2004; Demir, 2004; Kaya et al., 2003; Pelletier and Dione, 2004; Sanches-blanco et al., 2004).
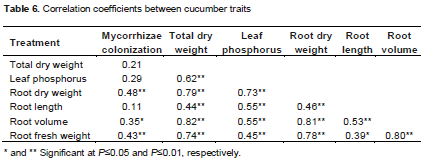
Correlation coefficients of traits showed that mycorrhizae colonization with root dry weight (r=0.48**), root volume (r=0.35*) and fresh root weight (r=0.43**) had significant differences (Table 6). In addition, leaf phosphorus (r = 0.62**), root dry weight (r = 0.79**), root length (r=0.44*), root volume (r = 0.82**) and root fresh weight (r=0.74*) had significant differences with above-ground dry matter. These observations indicate that plants having a higher leaf phosphorus, root dry weight and root volume produce higher total dry weight.
CONCLUSIONS
Inoculated plants with G. mosseae and G. intraradices showed more Leaf phosphorus, root fresh and dry weight, root length and volume than control. Root related traits such as root fresh and dry weight, root length and root volume increased in more phosphorus application and consequently will lead to increase above-ground dry matter. Relationships between traits showed that with increasing leaf phosphorus, root dry weight and root volume in inoculated mycorrhizal cucumber plants enhanced above-ground dry matter. Furthermore, since the formation of mycorrhizae often leads to increases in root traits and above-ground dry matter, the effect of mycorrhizae on Leaf phosphorus is also probably partly caused by the enhanced phosphorus nutrition. The overall results show that the use of mycorrhizal fungi is an essential element for the production of strong seedlings of cucumber and reduction in consumption of phosphate fertilizers.
CONFLICT OF INTEREST
The author did not declared any conflict of interests.
REFERENCES
|
Al-Karaki GN, McMichael B, Zak J (2004). Field response of wheat to arbuscular mycorrhizal fungi and drought stress. Mycorrhiza 14:263-269. Crossref |
||||
|
Allen MF, Smith WK, Moore TS, Christensen M (1981). Comparative water relations and photosynthesis of mycorrhizal and non-mycorrhizal Bouteloua gracilis H.B.K. Lag ex Steud. New Phytol. 88:683-693. Crossref |
||||
| Balakrishnan N, Subramanian KS (2012). Mycorrhizal symbiosis and bioavailability of micronutrients in maize grain. Maydica 57:129-138. | ||||
|
Burleigh SH, Cavagnaro T, Jakobsen I (2002). Functional diversity of arbuscular mycorrhizas extends to the expression of plant genes involved in P nutrition. J. Exp. Bot. 53:1593-1601. Crossref |
||||
| Demir S (2004). Influence of arbuscular mycorrhiza on some physiological, growth parameters of pepper. Turkish J. Biol. 28:85-90. | ||||
|
Giovannetti M, Mosse B (1980). An evaluation of techniques for measuring vesicular arbuscular mycorrhizal infection in roots. New Phytol. 84:489-500. Crossref |
||||
|
Graca MAS, Barlocher F, Gessner MO (2005). Methods to study litter decomposition: A practical guide. Springer-Verlag, Dordrecht, the Netherlands. 329 p. Crossref |
||||
| Graham JH (2000). Assessing cost of arbuscular mycorrhizal symbiosis in agroecosystems. In: Podila GK, Douds DD eds. Current Advances in Mycorrhizae Research pp. 127-140. APS Press, St Paul. | ||||
|
Grant C, Bittman S, Montrea M, Plenchette C, Morel C (2005). Soil and fertilizer phosphorus: Effects on plant P supply and mycorrhizal development. Canadian J. Plant Sci. 85:3-14. Crossref |
||||
|
Hao ZP, Christie P, Qin L, Wang CX, Li XL (2005). Control of fusarium wilt of cucumber seedlings by inoculation with an arbuscular mycorrhical fungus. J. Plant Nut. 28:1961-1974. Crossref |
||||
|
Hayman DS (1983). The physiology of vesicular-arbuscular endomycorrhizal symbiosis. Canadian J. Bot. 61:944-963. Crossref |
||||
|
Huang RS, Smith WK, Yost RS (1985). Influence of vesicular-arbuscular mycorrhiza on growth, water relations, and leaf orientation in Leucaena leucocephala Lam. De Wit. New Phytol. 99:229-243 Crossref |
||||
|
Jennings DH (1995). The Physiology of Fungal Nutrition. Cambridge: Cambridge University Press. Crossref |
||||
|
Kiers ET, Adler LS, Grman EL, Heijden MGA (2010). Manipulating the jasmonate response: How do methyl jasmonate additions mediate characteristics of aboveground and below-ground mutualisms? Func. Ecol. 24:434-443. Crossref |
||||
|
Marulanda A, Porcel R, Barea M, Azcon R (2007). Drought tolerance and antioxidant activities in laventies in lavender plants colonized by native drought-tolerant or drought-sensitive Glomus species. Mic. Ecol. 54:543-552. Crossref |
||||
| Marschner H (1996). Mineral Nutrition of Higher Plants. London: Academic Press. | ||||
| Marschner H, Dell B (1994). Nutrient uptake in mycorrhizal symbiosis. Plant and Soil 159:89-102. | ||||
| Miller JC, Jr, Rajapakse S, Garber RK (1986). Vesicular-arbuscular mycorrhizae in vegetable crops. Horti. Sci. 2l:974-984. | ||||
|
Mollier A., Pellerin S (1999). Maize root system growth and development as influenced by phosphorus deficiency. J. Exp. Bot. 50:487-497. Crossref |
||||
|
Ocampo JA, Azcon R (1985). Relationship between the concentration of sugars in the roots and VA mycorrhizal infection. Plant and Soil 86:95-100. Crossref |
||||
| Parewa HP, Rakshit A, Rao AM, Sarkar NC, Raha P (2010). Evaluation of maize cultivars for phosphorus use efficiency in an Inceptisol. Int. J. Agri. Environ. Biotech. 3:195-198. | ||||
| Pelletier S, Dionne J (2004). Inoculation rate of Arbuscular Mycorrhizal Fungi Glomus intraradices and Glomus etunicatum affects establishment of landscape Turf with no irrigation or fertilizer inputs. Crop Sci. 44:335-338. | ||||
|
Phillips JM, Hayman DS (1970). Improved procedures for clearing roots and staining parasitic and vesicular arbuscular mycorrhizal fungi for rapidassessment of infection. Trans. British Mycol. Soci. 55:158-161. Crossref |
||||
|
Plenets D, Mollier A, Pellerin SI (2000). Growth analysis of maize field crops under phosphorus deficiency. II. Radiation use efficiency, biomass accumulation and yield components. Plant and Soil 224:259-272. Crossref |
||||
| Podila GK, Douds DD (2001). Current Advances in Mycorrhizae Research. APS Press, St. Paul. | ||||
| Temperini O, Rouphael Y, Parrano L, Biagiola E, Colla G, Mariotti R, Rea E, Rivera CM (2009). Nursery inoculation of pepper with arbuscular mycorrhizal fungi: An effective tool to enhance transplant performance. Acta Horti. 807:591-596. | ||||
| Rakshit A, Bhadoria PBS, Das DK (2002). An overview of mycorrhizal symbioses. J. Interacademicia 6:570-581. | ||||
|
Rouphael Y, Cardarelli M, Mattia ED, Tulli M, Rea E, Colla G (2010). Enhancement of alkalinity tolerance in two cucumber genotypes inoculated with an arbuscular mycorrhizal biofertilizer containing Glomus intraradices. Biol. Fertil. Soils 46:499-509. Crossref |
||||
|
Ryan MH, Herwaarden AF, Angus JF, Kirkegaard JA (2005). Reduced growth of autumn-sown wheat in a low-P soil is associated with high colonisation by arbuscular mycorrhizal fungi. Plant and Soil 270:275-286. Crossref |
||||
|
Sanchez-Blanco MJ, Ferrandez T, Morales MA, Morte A, Alarcon JJ (2004). Variations in water status, gas exchange, and growth in Rosmarinus officinalis plants infected with Glomusdeserticola under drought conditions. J. Plant Physiol. 161:675-682. Crossref |
||||
|
Smith SE, Smith FA (2011). Roles of arbuscular mycorrhizas in plant nutrition and growth: new paradigms from cellular toecosystem scales. Ann. Rev. Plant Biol. 62:227-250. Crossref |
||||
|
Subramanian KS, Tenshia V, Jayalakshmi K (2009). Biochemical changes and zinc fractions in arbuscular mycorrhizal fungus (Glomus intraradices) inoculated and uninoculated soils under differential zinc fertilization. Appl. Soil Ecol. 43:32-39. Crossref |
||||
|
Subramanian KS, Santhanakrishnan P, Balasubramanian P (2006). Responses of field grown tomato plants to arbuscular mycorrhizal fungal colonization under varying intensities of drought stress. Sci. Horti. 107:245-253. Crossref |
||||
|
Trimble MR, Knowles NR (1995). Influence of vesicular–arbuscular mycorrhizal fungi and phosphorus on growth, carbohydrate partitioning and mineral nutrition of greenhouse cucumber (Cucumus sativus L.) plants during establishment. Canadian J. Plant Sci. 75:239-250. Crossref |
||||
|
Valentine AJ, Osborne BA, Mitchell DT (2001). Interactions between phosphorus supply and total nutrient availability on mycorrhizal colonization, growth and photosynthesis of cucumber. Scien. Horti. 88:177-189. Crossref |
||||
|
Vosatka M, Gryndler M (1999).Treatment with culture fractions from Pseudomonas putida modifies the development of Glomus fistulosum mycorrhiza and the response of potato and maize plants to inoculation. Appl. Soil Ecol. 11:245-251. Crossref |
||||
|
Watanabe FS, Olsen SR (1965). Test of an ascorbic acid method for determining phosphorus in water and NaHCO3 extracts from soil. Soil Sci. Soci. America 29:677-678. Crossref |
||||
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


