Full Length Research Paper

ABSTRACT
Biochemical changes of Atlantic salmon (Salmo salar) ï¬llets during superchilled storage were evaluated. Due to the signiï¬cant differences in ice crystal sizes observed, the biochemical changes were evaluated both at the top and centre parts of the superchilled samples. No signiï¬cant differences were found in biochemical changes between top and centre parts of the superchilled samples. The amount of cell tissue fluid (CTF) increased significantly from day 1 to 3 in the top and centre parts of the superchilled samples. The amount of CTF was stable between day 3 and 14 but increased significantly from day 21 to 28. A significant increase in the activity of β-N-acetyl-glucosaminidase in the CTF at the top was observed between day 3 and 7, while at the centre at day 7 of storage. There was also a significant increase in β-N-acetyl-glucosaminidase both at the centre and the surface of the superchilled samples between day 21 and 28 of storage whereby the activity in these samples was on the same level as in the frozen samples. In the superchilled samples, the cathepsin B activity in CTF and homogenates were stable for the first one week of storage. There was significant increase in activity of cathepsin B in cell tissue fluid and homogenates between day 21 and 28 for both chilled and superchilled samples. These ï¬ndings provides valuable information on the quality of food products for the food industry in relation to ice crystallisation/recrystallisation during superchilled storage.
Key words: Partial freezing, biochemical changes, superchilled storage, salmon fish (Salmo salar).
INTRODUCTION
Partial freezing of food is an effective way of preservation and it has been shown to double shelf-life of foods (Duun and Rustad, 2008; Duun et al., 2008). Superchilling and superchilled storage of food may, however, lead to quality changes and protein denaturation, especially of the myofibrillar proteins. These changes may result in altered functional properties, such as changes in texture, reduced water holding capacity and juiciness. These properties are related to food quality and are important for maintaining high quality.
Quality is an arbitrary term and one which causes confusion among consumers, processors and researchers. Product quality is a very complex concept (Gao, 2007) which included: Nutritional, microbiological, biochemical and physiochemical attributes. Quality not only relates to the freshness of food, how old it is and the appearance, it also related to the service received by the consumers and the consumer’s expectations and specifications (Gao, 2007). Microbial growth, colour, taste, texture, water holding capacity and juiciness, off-flavour, biochemical properties and oxidation are important factors both for the safety and the quality of food products. The deterioration in the food quality during superchilled storage is due to undesirable processes taking place in lipids and proteins (Duun and Rustad, 2008; Jiang and Lee, 1985; Ocano-Higuera et al., 2009). The degree of deterioration is influenced by many factors such as freshness and treatment before partial freezing, state of rigor, partial freezing rate, ultimate partial freezing temperature, storage temperature, storage period, fluctuation of storage temperature, and thawing methods (Benjakul and Visessanguan, 2010; Blond and Meste, 2004; Chevalier et al., 2001; Duun and Rustad, 2008; Duun et al., 2008; Hagiwara et al., 2002; Jiang and Lee, 1985; Kiani and Sun, 2011; Martino et al., 1998; Mittal and Griffiths, 2005; Petzold and Aguilera, 2009).
There is growing demand for fresh and high quality foods worldwide. In recent years, consumers are increasingly concerned with their health, and are demanding foods that are beneficial to their health and help prevent diseases. This leads to an increased need for technologies to preserve the freshness of foods. Superchilled storage is an efficient way of preserving the quality of food (Kaale et al., 2013b). With good handling and safety practices both before and after partial freezing, superchilled foods can retain high quality. To retain good quality, the superchilled foods should be kept at stable superchilled storage temperatures (Banerjee and Maheswarappa, 2017). At these temperatures, growth of micro-organisms is very low and deteriorative reactions take place at very slow rates (Duun and Rustad, 2008).
Nevertheless, studies by Kaale et al. (2013b) and Kaale et al. (2013c) reported that a high superchilling rate results in a high rate of heat removal, leading to the formation of a large number of small nuclei and thus a large number of small ice crystals that grow both within and outside cells. As a result, the cells maintain their integrity which in turn minimises drip loss; maintain water holding capacity and textural changes during thawing (Smith, 2011). However, this advantage was reduced during superchilled storage by the rapid growth of ice crystal sizes in the salmon fillets (Kaale et al., 2013b). Moreover, there was a large difference between the ice crystal sizes at the surface and centre of the superchilled salmon fillets. The increase in ice crystal sizes during superchilled storage may impart mechanical damage by physically rupturing cell walls, which may result in an increase in drip loss, a reduction of the water-holding capacity and textural changes (Smith, 2011).
Therefore, there is a need to study the damage which may take place during superchilled storage of salmon fillets. When muscle foods are partially frozen, cell and organelle membranes are destroyed leading to leakage of lysosomal enzymes into the cell (Nilsson and Ekstrand, 1993). The degree of membrane destruction can be measured as an increase in amount of cell tissue fluid (CTF). Increase in lysosomal enzyme activity in the CTF is also a measure of membrane destruction. Increased amount of CTF is closely related to loss of water holding capacity and increase with reduced ability of the tissue to hold water (Duun and Rustad, 2008; Johansen, 2013). Hence, the objective of this study was to assess cell tissue fluid and enzyme activity changes both at the surface and centre of the salmon fillets in order to clarify the effect of the ice crystal development during superchilled storage.
MATERIALS AND METHODS
Salmon fillets (Salmon salar) stored in ice (0.9 to 1 kg), were delivered by Lerøy Midnor (Hitra, Norway). The fillets were vacuum-packed and stored at 4°C for 24 h before the superchilling process to ensure a constant temperature in all samples. Superchilling was performed in an Impingement Advantec Lab Freezer (JBT Food-Tech, Rusthållsgatan 21, SE-251 09, Helsingborg, Sweden) at NTNU Energy’s laboratory in Trondheim, Norway. The samples were superchilled (partially frozen) at -30°C and 227 W/m2.K (at 2.5 kPa pressure differences of the fan at the impingement freezer) for 2.1 min to achieve an ice content of 20% (Kaale et al., 2013c).
Temperature trend during storage
The temperature, as one of the critical parameters during superchilled storage, was strictly controlled during this study. The storage box was designed (92 × 73 × 54.5 cm) with a heating element inside to ensure adequate temperature regulation. Three Pt100 temperature sensors were inserted in the storage box: One was used to measure the air temperature, and the other two were used to measure the surface and centre temperatures of the superchilled salmon fillets. The set-point temperature was -1.7°C. The box was placed inside the storage room, which was at a temperature of approximately -5°C (temperature outside the storage box).
Cell tissue fluid (CTF)
Approximately 20 g of coarsely cut muscle was weighed out in a centrifuge tube and centrifuged at 4°C for 30 min at ~28 000 x g in SS-34 rotor (Sorvall centrifuge, The Netherlands) (Nilsson and Ekstrand, 1994). The supernatant was pipetted off, and this is the CTF. Quantification of the CTF was made by weighing directly into Eppendorf tubes. Samples were kept at -80°C until analysis. Two parallels were made from each sample.
Protein extraction (homogenate preparation)
Extraction of water soluble protein was performed as described by Nilsson and Ekstrand (1994)at +4°C, and samples were kept on ice before, under and after homogenization. Each sample was extracted once. For protein fractionation, approximately 5 g of muscle was cut from the loin side and weighed out in a centrifuge tube (50 mL), and 15 mL of 50 mM phosphate buffer, pH 7.0 was added before homogenization (Ultra Turrax). The homogenate was left in a cold room for 30 min, occasionally stirred or shaken before centrifugation at 4°C for 30 min at ~14 700 x g in a SS-34 rotor (Sorvall centrifuge). The supernatant was decanted into a volumetric flask, through a funnel with glass wool. The glass wool was rinsed with buffer, and the volume was made up to 25 ml. Aliquots of the extracts were frozen at -80°C until analyses.
Enzymatic activity
Activity of the lysosomal enzymes cathepsin B and N-acetylaminidase were determined in CTF and extract of water soluble proteins. The cathepsin B activity was determined with a synthetic fluorogenic substrate, Nα-carbobenoxy-L-arginyl-L-7-amido-4-methylcoumarin (Barrett and Kirschke, 1981). Activity of β-N-acetylglycosaminidase (NAG) was determined using ρ-nitrophenyl-N-acetyl-b-D-glucose amide as a substrate. The activity was determined as described by Milanesi and Bird (1972).
Statistical analysis
The observations of the CTF at the two locations with respect to storage days were analysed by one- and two-way analyses of variance using Minitab 16 software (Minitab, Inc., USA). A general linear model, (post- hoc test) under Tukey’s simultaneous test was applied whenever the ANOVA results were significant (p < 0.05).
RESULTS AND DISCUSSION
Ice crystal evolution during the superchilling process and the superchilled storage
Figure 1 showed the micrographs of superchilled and without superchilled salmon tissues (Control). The study by Kaale et al. (2013a) reported that the ice crystal size in zero-time (superchilling process) was significantly smaller (23 ± 3 µm) compared to that during storage of superchilled samples (92 µm), which is 4 folds larger than ice crystals at zero time. This is because the superchilling process was performed at a very low temperature (-30°C) and the samples were stored at -1.7 ± 0.3°C.
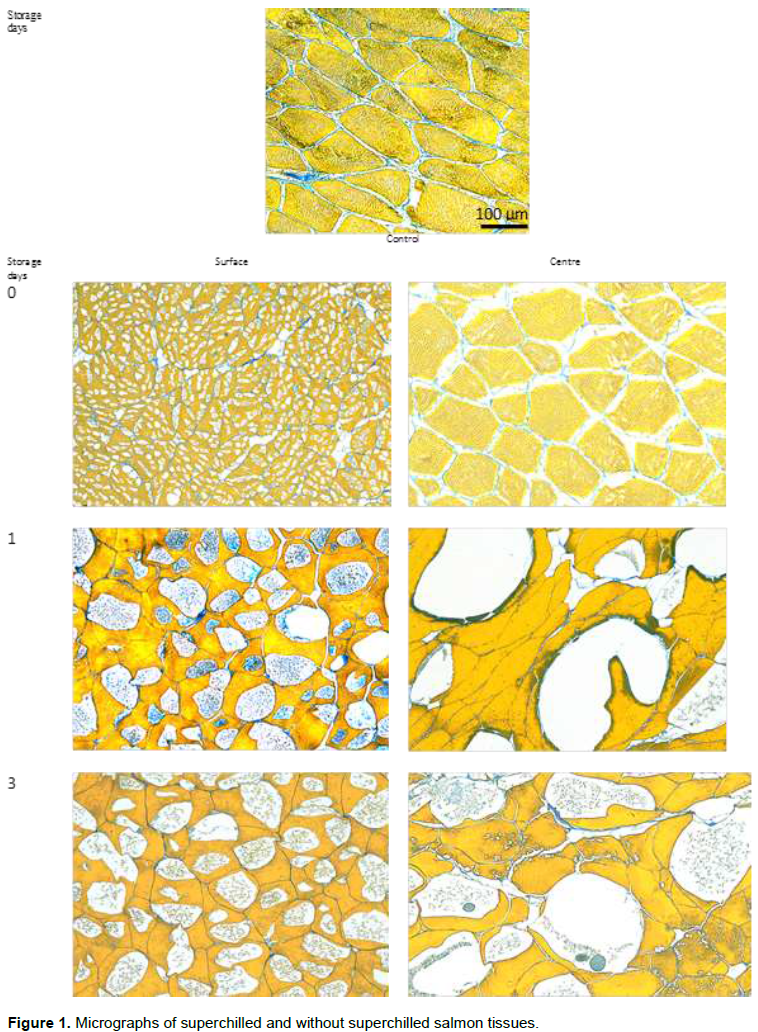
In addition, thermal gradients which developed during the superchilling process in the product near the surface were reported to contribute to the increase of ice crystal size during superchilled storage. When temperature equalisation was achieved within the samples, the growth of the intracellular ice crystals at the surface layer was not significant (p < 0.05) (Kaale et al., 2013a). Prior to temperature equalisation, ice crystal growth progresses from the surface to the centre of the superchilled food products (Figure 1). No ice crystals were formed at the centre during the superchilling process (zero time). The size of ice crystal that are formed at the centre was significantly larger than that at the surface (318 ± 4 µm_ after only 1 day of storage, which is 3-folds larger than ice crystals at surface. The large size is due to slow superchilling rate, and these crystals may damage the integrity of the superchilled product. Due to the significant difference in ice crystal sizes (surfaces vs centres) within the superchilled salmon samples, cell tissue fluid and enzyme activity were analysed separately at the surfaces and centres of the fillets during superchilled storage.
Cell tissue fluid (CTF)
Figures 2 and 3 showed the amount of CTF at the centre and surface of the superchilled samples. The amount of CTF increased during storage. No signiï¬cant differences were found in CTF between the top and centre parts of the superchilled samples. Kaale et al. (2013a) reported that the temperature equalization had taken place at day 1 and there was no significant growth of ice crystal size after this time. In this study, there was a significant increase in amount of CTF from day 1 to 3 at the top and the centre parts of the superchilled samples. The amount of CTF was stable between day 3 and 14 but increased significantly from day 21 to 28.
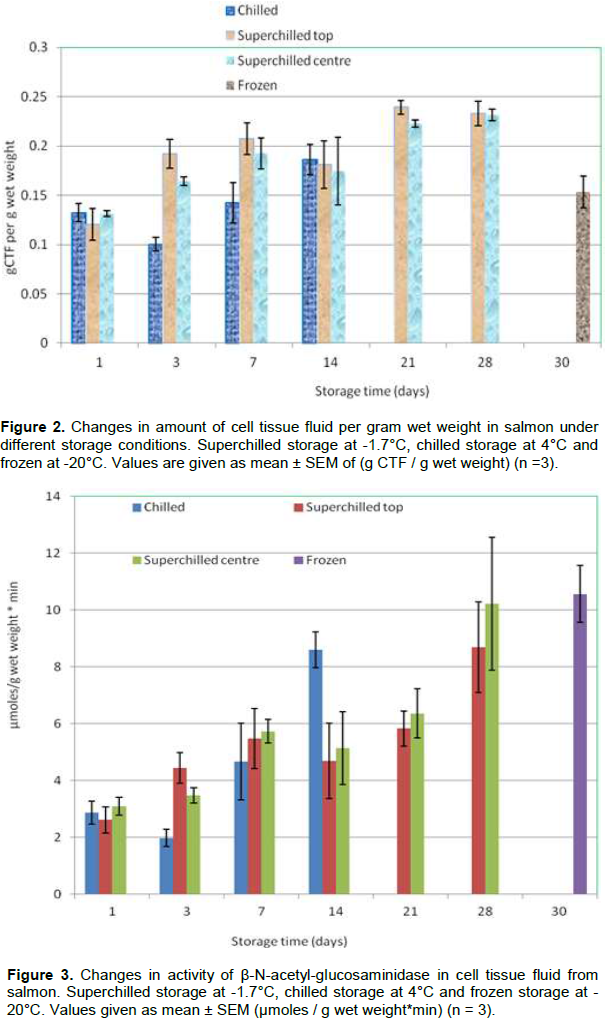
The changes in CTF showed a slight damage of cell membranes between day 1 and 3 where temperature equalization takes place and between day 21 and 28 (Figure 2). These results are according to that reported in previous study (Kaale et al., 2014). The type of samples and storage procedures in this study are the same as that used in Kaale et al. (2014).
Enzyme activity
Leakage of lysosomal enzymes such as different cathepsins, β-N-acetyl-glucosaminidase and a-glucosidase have been used as an indicator of membrane damage due to freezing (Nilsson and Ekstrand, 1993; 1994, Bahuaud et al., 2008). The significant increase in the activity of β-N-acetyl-glucosaminidase in the CTF at the top was observed between day 3 and 7, while at the centre on day 7. In contrast, the β-N-acetyl-glucosaminidase in the CTF increased signiï¬cantly between day 21 and 28 at the centre and the surface of the superchilled samples and the activity in these samples (day 21 and 28) was on the same level as in the frozen samples. No signiï¬cant differences were found in the β-N-acetyl-glucosaminidase in the CTF between top and centre parts of the superchilled samples.
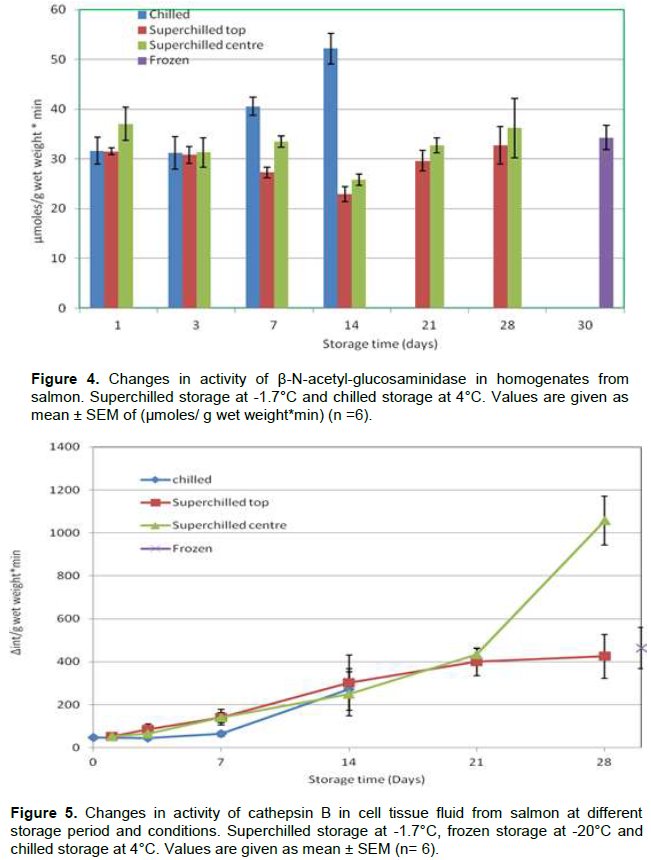
The activity of β-N-acetyl-glucosaminidase in extracts of water soluble proteins (Figure 4) from the chilled samples increases between day 7 and 14 of storage while in the superchilled samples there is a decrease between day 7 and 14 for the top samples and at the centre was observed at day 14 of storage, followed by an increase to the same level as in the samples from day one. This could imply that there are inhibitors that are inactivated during chilled storage while this does not take place till after 14 days of superchilled storage.
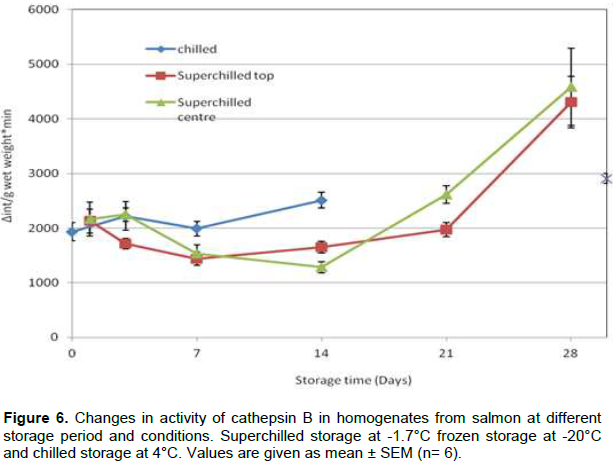
Cathepsins are also lysosomal enzymes, and the cathepsin B activity in CTF (Figure 5) was stable for the first one week of storage in all samples. There was a significant increase in activity of cathepsin B in cell tissue fluid between day 21 and 28 for chilled and superchilled samples which might be due to cell denaturation during superchilled storage.
For the homogenates (Figure 6), there is a slight increase in the activity for the chilled samples while the activity in the superchilled samples decrease during the first 1 to 2 weeks and then increase from day 21 to 28.
No signiï¬cant differences were found in cathepsin B activity in CTF between top and centre parts of the superchilled samples.
CONCLUSION
No signiï¬cant differences were found in biochemical changes between top and centre parts of the superchilled samples. The amount of CTF increased significantly from day 1 to 3 in the top and centre parts of the superchilled samples. The amount of CTF was stable between day 3 and 14 but increased significantly from day 21 to 28. A significant increase in the activity of β-N-acetyl-glucosaminidase in the CTF at the top was observed between day 3 and 7, while at the centre at day 7 of storage. There was also a significant increase in β-N-acetyl-glucosaminidase in both the centre and the surface of the superchilled samples between day 21 and 28 of storage whereby the activity in these samples is on the same level as in the frozen samples. In the superchilled samples, the cathepsin activity in CTF and homogenates were stable for the first one week of storage. There was significant increase in activity of cathepsin B in cell tissue fluid and homogenates between day 21 and 28 for both the chilled and superchilled samples. These ï¬ndings provides valuable information on the quality of food product for the food industry in relation to ice crystallisation/recrystallisation during superchilled storage.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors are grateful to the Research Council of Norway (RCN project number 178280) for its ï¬nancial support.
REFERENCES
|
Bahuaud D, Mørkøre T, Langsrud Ø, Sinnes K, Veiseth E, Ofstad R, Thomasse MS (2008). Effects of −1.5°C superchilling on quality of Atlantic salmon (Salmo salar) pre-rigor fillets: Cathepsin activity, muscle histology, texture and liquid leakage. Food Chemistry 111:329-339. |
|
|
Banerjee R, Maheswarappa NB (2017). Superchilling of muscle foods: Potential alternative for chilling and freezing. Critical Reviews in Food Science and Nutrition pp.1-8. |
|
|
Barrett AJ, Kirschke H (1981). Cathepsin B, cathepsin H, Cathepsin L (eds.), Methods in Enzymology pp. 535-561. |
|
|
Benjakul S, Visessanguan W (2010). Impacts of freezing and frozen storage on quality changes of seafoods. In: Devahastin S (eds.), Physicochemical aspects of food engineering and processing. CRC press, New York pp. 283-306. |
|
|
Blond G, Meste ML (2004). Principles of frozen storage. In Handbook of frozen foods. CRC Press pp. 33-61. |
|
|
Chevalier D, Sequeira-Munoz A, Bail AL, Simpson BK, Ghoul M (2001). Effect of freezing conditions and storage on ice crystal and drip volume in turbot (Scophthalmus maximus) Evaluation of pressure shift freezing vs. air-blast freezing. Innovative Food Science Emerging Technology 1:193-201. |
|
|
Duun AS, Hemmingsen AKT, Haugland A, Rustad T (2008), Quality changes during superchilled storage of pork roast. LWT - Food Science and Technology 41(10):2136-2143. |
|
|
Duun AS, Rustad T (2008). Quality of superchilled vacuum packed Atlantic salmon (Salmo salar) fillets stored at −1.4 and −3.6 °C. Food Chemistry 106(1):122–131. |
|
|
Gao HY (2007). Methods of pre-cooling for fresh cod (Gadus morhua) and influences on quality during chilled storage at 1.5 oC. The United Nations University, Fisheries Training Programme, Iceland. Available at: |
|
|
Hagiwara T, Wang H, Suzuki T, Takai R (2002). Fractal analysis of ice crystals in frozen food. Journal of Agricultural Food Chemistry 50:3085-3089. |
|
|
Jiang ST, Lee TC (1985). Changes in free amino acids and protein denaturation of fish muscle during frozen storage. Journal of Agricultural Food Chemistry 33:839-844. |
|
|
Johansen T (2013). Superchilling of Salmon (Salmo salar). MSc. thesis, Norwegian University of Science and Technology (NTNU), Norway. Available at: https://brage.bibsys.no/xmlui/handle/11250/245920 |
|
|
Kaale LD, Eikevik TM, Bardal T, Kjorsvik E (2013a). A study of the ice crystals in vacuum-packed salmon fillets (Salmon salar) during superchilling process and following storage. Journal of Food Engineering 115:20-25. |
|
|
Kaale LD, Eikevik TM, Bardal T, Kjorsvik E, Nordtvedt TS (2013b). The effect of cooling rates on the ice crystal growth in air-packed salmon fillets during superchilling and superchilled storage. International Journal Refrigerating 36:110-119. |
|
|
Kaale LD, Eikevik TM, Kolsaker K, Stevik AM (2013c). Modelling and simulation of food products in superchilling technology. Journal of Aquatic and Food Production Technology 23:409-420. |
|
|
Kaale LD, Eikevik TM, Rustad T, Nordtvedt TS (2014). Changes in water holding capacity and drip loss of Atlantic salmon (Salmo salar) muscle during superchilled storage. LWT - Food Science and Technology 55(2):528-535. |
|
|
Kiani H, Sun D-W (2011). Water crystallization and its importance to freezing of foods: A review. Trends in Food Science Technology 22:407-426. |
|
|
Martino MN, Otero L, Sanz PD, Zaritzky NE (1998). Size and location of ice crystals in pork frozen by high-pressure-assisted freezing as compared to classical methods. Meat Science 50:303–313. |
|
|
Milanesi AA, Bird JWC (1972). Lysosomal enzymes in aquatic species - II. Distribution and particle properties of thermally acclimated muscle lysosomes of rainbow trout, Salmo gairdneri. Comparative Biochemistry Physiology Part B: Comparative Biochemistry 41(3):573-591. |
|
|
Mittal GS, Griffiths MW (2005). Pulsed electric field processing of liquid foods and beverages. In Emerging technologies for food processing. Food science and technology, international series, Elsevier Ltd. pp. 99-139. |
|
|
Nilsson K, Ekstrand Bo (1993). The effect of storage on ice and various freezing treatments on enzyme leakage in muscle tissue of rainbow trout (Oncorhynchus mykiss). Zeitschrift für Lebensmittel-Untersuchung und Forschung 197(1):3-7. |
|
|
Nilsson K, Ekstrand Bo (1994). Enzyme leakage in muscle tissue of rainbow trout (Oncorhynchus mykiss) related to various thawing treatments. Zeitschrift für Lebensmittel-Untersuchung und Forschung 198(3):253-257. |
|
|
Ocano-Higuera VM, Marquez-Rios E, Canizales-Davila M, Castillo-Yanez FJ, Pacheco-Aguilar R, Lugo-Sanchez ME, Garcia-Orozco KD, Graciano-Verdugo AZ (2009). Postmortem changes in cazon fish muscle stored on ice. Food Chemistry 116:933-938. |
|
|
Petzold G, Aguilera JM (2009). Ice morphology: Fundamentals and technological applications in foods. Food Biophysics 4:378-396. |
|
|
Smith PG (2011). An Introduction to Food Process Engineering. In Introduction to Food Process Engineering. Springer, Boston, MA. pp. 1-3. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


