Full Length Research Paper

ABSTRACT
Lactic acid bacteria (LAB) strains isolated from six main Ivorian cocoa producer regions were investigated based on their biochemical properties in order to select the best one as potential starter. Three main technological and useful properties for good cocoa beans fermentation were monitored among the 568 isolated LAB strains. Thus, between the 408 cocoa-specific acidifying LAB strains identified, 05.88% (24 isolates) of them were able to maintain this activity in pH and temperature conditions as same as fermentation heap with an acidifying rate ranged within 0.03±0.00 to 1.19±0.070%. All these 24 cocoa-specific acidifying LAB strains displayed the ability to degrade the citrate while six (25%) of them were able to produce acetoin. These six LAB strains namely T1Gb8, T6C5, T6C12, T7C8, T9C9 and T11C5 were identified as Lactobacillus plantarum. They were the best able to continuously produce lactic acid in the required standards. Firstly under varying condition of temperature (30 à 45°C) and pH (3 à 6) with sharp acid (acetic, lactic and citric acids) influence. Secondly beyond 8% of ethanol content. The highlighted properties in this study and the performance of these six strains may make them best candidate as starters for cocoa fermentation control.
Key words: Lactic acid bacteria, cocoa-specific acidifying, technological properties, starter, Ivorian cocoa, fermentation.
INTRODUCTION
Cocoa beans constitute a basic export product for many tropical countries in America (Central and South), Asia (South and Southeast) and mainly West Africa. Export of raw cocoa beans is of great economic importance for West African countries which supply two thirds of the world's cocoa crop. Côte d'Ivoire is the world's leading producer of cocoa beans since 1978 (Deheuvels, 2003). Today, this country is highly dependent on this crop which accounts for 50% of national export income and 22% of the Gross Domestic Product (ICCO, 2016).Cocoa beans are an important source of cocoa powder and the principal raw material of chocolate. The sensory and physical properties of West African cocoa make it a much sought-after product, although overall production quality seems to be declining (Afoakwa et al., 2013). Unfortunately, ensuring the marketable quality of cocoa beans remains difficult due to variability in the organoleptic quality of this product. This inconsistency in cocoa quality is mainly linked to post-harvest processing including fermentation, drying and roasting of the crude and fresh cocoa beans. Fermentation is considered as the most important processing step influencing the quality of cocoa beans. Indeed, during this process, there is initiation of flavor precursors, color development and a significant reduction in bitterness and astringency of the beans (Afoakwa et al., 2013). These changes involve the action of diverse microrganisms naturally found in the cocoa pulp. The succession of these microbial groups has been clearly established and lactic acid bacteria (LAB) population is considered as the second microbial group growing in mucilaginous pulp of cocoa beans after yeast population (Crafack et al., 2013). This micro-biota group is mainly dominating until 24 to 48 h of fermentation (Kouame et al., 2015). The most important roles of LAB are the degradation of the citric acid in the pulp, leading to increase in pH, the production of ethanol and both volatile and non-volatile organic acids such as acetic and lactic acids (Penia et al. 2013). Organic acid will diffuse into the beans cotelydon and induce the enzymatic reactions to form precursor of flavor, aroma and color (Afoakwa et al., 2013). The diffusion of certain amount of acid into the bean during fermentation is essential for the development of flavor precursors. Indeed, the presence of excessive acidity usually correlates with poor development of chocolate flavor (Serra et al., 1997). Thus, acidity is a critical factor in deï¬ning the quality of cocoa used in chocolate manufacturing. The control of cocoa fermentation process is very challenging for farmers in cocoa producing countries because cocoa fermentation has always been a natural process which sometimes results in poorly fermented cocoa. Therefore, studies suggest the use of starter microbial culture as the best approach to improve the fermentation process. In this perspective, LAB strains as monoculture or co-culture would be essential component of starter culture aimed at the control of cocoa beans fermentation process. Studies conducted by differents authors, such as Papalexendratou et al. (2013), Penia et al. (2013) etc., on cocoa fermentation showed that the maximum rate of lactic acid produced ranged between 0.5 and 1.2%. As excessive acidity, mainly lactic acid, produces unsuitable
raw cocoa for chocolate making, it appears more judicious to isolate, screen and characterize LAB strains with cocoa-specific acidifying potential included in this limit of 1.2%. These strains may positively and durably influence the quality of a controlled cocoa fermentation especially if they provide useful additional properties such as citrate degradation and acetoin production (Rodriguez-Campos et al., 2012). Information on cocoa-specific acidifying LAB strains with adequate lactic acidifying potential, citrate degradation and acetoin production may be a starting point for the development of starter culture. Thus, the aim of the research was to select strains of LAB that best meet these different criteria in order to optimize and standardize the cocoa fermentation process.
MATERIALS AND METHODS
Fermentation condition
Cocoa pods were harvested on farms from six cocoa producing regions of Ivory Coast: Cavally (6° 25' 0 North -7° 30' 0 West), Gkôklè (4° 57′ 04″ north 6° 05′ 19″ West), Gôh (6° 15’ 0’’ North 5° 55’ 0’’ West), Haut-Sassandra (7° 0' 0 North -6° 34' 59 West), San-Pedro (4° 44' 54 North -6° 38' 10 West) and Tonkpi (7° 24’ 45’’ North 7° 33’ 14’’ West). Spontaneous cocoa bean fermentation was performed using traditional conditions, with temperature and humidity ranging respectively, from 28 to 30°C, and 60 to 65% for 6 days. The fermenting mass (50 kg) set on banana leaves and covered with banana leaves were constituted of mixed genotypes (Forastero, Trinitario and Criollo cultivars).
Sampling
The fermenting heap was mixed and 100 g beans were collected, at the beginning and each 12 h of fermentation, in sterile Stomacher bag. A total, 13 samples of each fermenting cocoa according to the locality were collected for physico-chemical and microbial analysis.
Isolation of bacterial strains
Isolation of LAB was performed according to the standard method described by Pereira et al. (2012). An amount of 25 g of fermenting cocoa beans was homogenized in 225 mL sterile peptone water in a Stomacher bag (Seward, Worthington, United Kingdom) for 5 min at room temperature. After appropriate dilution of samples in sterile saline solution, 0.1 mL from each dilution was inoculated onto duplicate plates of MRS agar (Oxoid) supplemented with 50 μg/mL of nystatin to inhibit the fungal growth. Plates were incubated at 30°C for 48 to 72 h, under anaerobic conditions.
Preliminary biochemical test of lactic acid bacteria
LAB isolates, identified using conventional colonial morphology, gram staining and biochemical reactions (oxidase and catalase tests), were stored in cryotubes at -20°C in MRS broth supplemented with 20% glycerol for further investigations.
Screening of cocoa-specific acidifying LAB strains
Cocoa-specific acidifying LAB strains were screening based on their acidification capacity at 30°C according to the protocol of Guha et al. (2013). 100 μL of a preculture recording an optical density (OD) of 1 at 600 nm against a control (MRS broth) were used to inoculate 5 mL of MRS broth. The seeded broths were incubated at 30°C in a water bath (Julabo TWB 12) for 48 h. Then, fermented broths were centrifuged at 4500 trs/min for 10 min to pellet out the bacterial growth. Supernatant was used to estimate lactic acid content by titrimetric method with 0.1 NaOH. The percentage of lactic acid was calculated using the formula in equation 1. Each treatment was tested in triplicate.
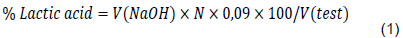
V(NaOH) : Volume of NaOH solution used for titration
N : Normality of the NaOH solution (0.1)
V (test): Sample Volume
Analysis of additional properties of cocoa-specific acidifying lactic acid bacteria isolates
Screening of LAB strains for citrate degradation
The ability of lactic acid bacteria to degrade citrate was investigated using the method described by Kempler and McKay (1980). A basal medium containing 1% milk powder, 0.25% casein peptone, 0.5% glucose and 1.5% agar was first prepared and then 1L of the medium, was supplemented with 10 mL of potassium ferricyanide (10%) solution and 10mL of mixed iron citrate and sodium citrate (2.5%) solution. The medium was inoculated with pure 24 h preculture of LAB strains and incubated in an anaerobic jar at 30 °C for 48 to 72 h. The citrate metabolism was assessed by the formation of a blue complex surrounding the colonies.
Screening of LAB strains for acetoin production
The production of acetoin was investigated using the method described by King (1948). Acetoin production from citrate was checked in the basal liquid medium. Clark and Lubs medium was seeded with pure 24 h preculture of LAB strains and incubated at 30°C for 48 h. After incubation time, acetoin production was revealed in 1mL of cell free supernatant by adding two drops of an alcoholic solution of alpha-naphthol (6%) and two drops of sodium hydroxide (16%) solution. The presence of acetoin was assessed by appearance of red color.
Effect of environmental stress on lactic acid production by cocoa-specific acidifying LAB strains
Influence of temperature and pH
The influence of temperature and pH on the acidification capacity of cocoa-specific acidifying lactic acid bacteria strains was evaluated by Guha et al. (2013) method. Seeded broths were incubated at 30, 35, 40, 45 and 50 °C to assess the influence of temperature and at pH 3; 4; 5 and 6 for pH influence, in a water bath (Julabo TWB 12) for 48 h.
Influence of alcohol and organic acids on lactic acid production by cocoa-specific acidifying LAB strains
To analyze the acidification potential of cocoa-specific LAB strains in alcoholic and acidic conditions, a MRS liquid medium was prepared. After autoclaving, this medium was cooled and maintained in liquid state at 45 °C and supplemented with alcohol (2, 4, 6 and 8%), acetic and lactic acid (0.2; 0.4; 0.6; 0.8; 1; 1.2%), citric acid (0.5; 1; 1.5; 2; 2.5 and 3%). The standard inoculum was prepared as follow: a 18 h preculture was suspended in a saline solution to obtain an absorbance OD 1 at 600 nm. 100 μL were used to inoculate 5 mL of MRS broth. The seeded broths were incubated at 30°C in a water bath for 48 h. After incubation time, the amount of total acid produced by each strain was determined by titration as previously described.
Molecular characterization of lactic acid bacteria starters
16S rRNA gene PCR and sequence analysis
To perform PCR reactions, The isolates of LAB strains were grown for 24 h at 30°C on agar plates. A loopful of pure culture was suspended in 100 µL of sterile distilled water for colony PCR.
PCR amplification of ribosomal 16S gene
The hypervariable regions (V1, V2 and V6) of the different LAB strains were amplified by using universal primers F27 (5’-AGAGTTTGATCCTGGCTCAG-3’) and R520 (5’-ACCGCGGCTGCTGGC-3’) (Ouattara et al., 2011) for colony PCR.
Each mixture (final volume 50 µL) contained about 1 µL of sample 1.25 U of Taq DNA polymerase (Biolabs, Lyon France), 5 µL of 10x Taq Buffer (10 mM TrisHCl, 50 mM KCl, 1,5 mM MgCl2), 1 µL of 10mM dNTP, 2 µL of each primer (10 μM) (Eurofins, Genomics, Germany) supplemented by sterile water Milli-Q. PCR amplification was carried out in a thermocycler (Sensoquest Labcycler) as described previously (Ouattara et al., 2011). After an initial denaturation at 95 °C for 4 min, reactions were run for 35 cycles. Each cycle comprising denaturation at 95 °C for 1 min, annealing at 56 °C for 1 min, extension at 72 °C for 1min; and final extension at 72 °C for 10 min.
Gel electrophoresis
After migration of the products realised at 70 V for 2 h in a tank, amplicons were revealed by incubating on 0.8% agarose gel electrophoresis in a 1x phosphate TAE buffer containing bromide of ethidium. The molecular weight of the products was estimated in reference to a molecular weight marker (Biolabs, France) of 500 bp.
Sequencing and analysis of hypervariable sequence of 16S RNA gene amplified
The PCR products were purified using the nucleospinR Gel and PCR Clean-up kit (MachereyeNagel, Germany). Then, they were sequenced by MWG Eurofins using the primer F27. The basic local alignment search tool (BLAST, blast N) from the NCBI database site (blast.ncbi.nlm.nih.gov/ BLAST/) was used to ï¬nd the closest sequences relative to the ampliï¬ed 16S RNA genes in order to identify our LAB strains
RESULTS AND DISCUSSION
Acidification capacity of LAB strains isolates
A total of 568 strains characterized as gram positive, oxydase negative and catalase negative identified as LAB were isolated from six Ivorian regions. Thus, these LAB strains were anlysed for their capacity to acidify the fermentative medium. The results showed that the number of isolated LAB strains have unequal distribution depending on the region. San Pédro recording the highest (21.47%) number of isolates (Table 1) unlike Gôh, which has the lowest number (08.45%) of strains. This variability indicated the influence of the local geographical area on the composition of the microflora involved in cocoa fermentation (Schwan and Wheals, 2004). Therefore, this factor could impact on the distribution of their different activities. One of the important activities attributed to LAB strains is to acidify the fermentative medium. As this property is susceptible to play a key role in the fermentative process (Afoakwa et al., 2013), it appears important to screen them based on this ability. Most importantly, this type of metabolic pathway should be determined to identify potential starters among these isolates.

All the 568 isolated LAB microflora which were screened for their acidification capacity showed naturally acidification capacity with different levels of acid production (Table 1). Indeed, this natural capacity for acidification of the fermentative medium by LAB is related to the fact that they are able to exhibit an enormous capacity to degrade different carbohydrates and related compounds. Generally, the predominant end product is lactic acid which is the main metabolite of two broad metabolic categories of LAB. The first metabolic category is represented by the homofermentative LAB. They convert glucose into lactic acid more than 90% by the Embden-Meyerhof (E-M) pathway. The second metabolic group is represented by the heterofermentative LAB producing about 50% of lactic acid as the principle by-product of sugar fermentations (Raimbiault 1995).
It should be noted that a acidification scale has been established based on the acidification capacity of LAB from different fermentation medium such as milk, cheese, wine, fish, meat, sausages, butter and cream (Behannis and Kayanush, 2012; Joko et al., 2014). The recorded lactic acid values (around twenty) ranged between 0.05% and 1.6% and allowed to identify three types of LAB strains according to their capacity of acidification: LAB strains with low acidity capacity (acidification rate (AR) < 0.45%); LAB strains with medium acidity capacity (0.45 < AR ≤1.35%) and those with high acidity capacity (AR ≥ 1.35%).
The results obtained are depicted in Table 1. Based on this classification, three types of LAB strains were observed in the different regions except in Cavally and Tonkpi with respect to the low lactic acid producing strains and the regions of Haut Sassandra and San Pédro with respect to the high producers. In addition, medium acidifying LAB strains dominated with 57.57% compared to 24.12% for high acidifying LAB strains and only 18.31% for low acidifying LAB strains. This diversity in acidifying capacity of LAB strains suggesting a difference in acidification potential and therefore efficiency in acidification by these strains. This could be the main determining factor in the variability of cocoa bean quality as cocoa beans acidification often has negative impact on the final quality of the market cocoa.
Therefore, the cocoa market could ultimately be related to cocoa bean quality which is dependant on selective LAB strains with satisfactory level of acidification. This approach could make it possible to ensure the control of the acid level of marketable beans and thus the control of their quality. With regard to lactic acid, the satisfactory level of acidification can only be obtained with LAB strains that do not have high acidification capacity. Indeed, the low volatility of this acid promotes its accumulation in beans. If this acid is produced in large quantities, it can hardly be eliminated. This will irremediably lead to acidic beans and therefore of poor quality. Thereby, LAB strains with low and medium acidification capacity (431) retained our attention, approximately two-thirds of LAB strains isolated.
Then, a second level of selection has been established to better refine our selection. This selection takes into account the maximum rate of lactic acid produced during the fermentation of cocoa. Studies conducted by Afokawa (2013), Kouame et al. (2015) and Papalexendratou et al. (2013; 2015) on cocoa fermentations, around the world, lead to the identification of a maximum rate of lactic acid produced of 1.2%. Thus, in order to mimic the natural conditions of fermentation, it is important that the selected LAB strains do not produce lactic acid beyond this limit (1.2%). The results of this selection are shown in Table 1. Among the 568 strains of LAB previously isolated, approximately two-thirds (71.83%) produced an amount of lactic acid that not exceed 1.2%. This total of 408 LAB strains applies to all the strains with low acidifying capacity (104) and part of the medium acidifying strains (304 of 327). These strains are considered to be "Cocoa-specific acidifying LAB strains". As high acidity, mainly lactic acid, in cocoa beans produces unsuitable raw cocoa, these 408 Cocoa-specific acidifying LAB strains appear to be the best candidates as potential starter culture to control cocoa bean fermentation.
Influence of temperature and pH on lactic acid production by Cocoa-specific acidifying LAB strains
The pH of cocoa beans during fermentation is critical with regard to the biochemical reactions that take place in the beans (Afokawa et al., 2013). The ability of the 408 Cocoa-specific acidifying LAB strains to maintain their acidifying potential ≤1.2% at temperatures up to 45°C is depicted in Table 2. Only 104 strains maintained their acidifying potential under 45°C of temperature among the 408 strains previously selected.
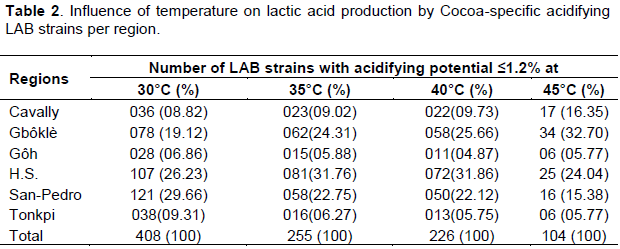
Cocoa fermentation is generally characterized by high temperature up to 45°C occurring between 48 and 72 h of fermentation (Ho et al., 2014). In view of these results, we can assess that the thermo-tolerance exhibited by these 104 heat-resistant strains should allow them to maintain a satisfactory level of lactic acid production under thermic stress conditions during cocoa fermentation.
On the other hand, the effect of pH variations on these 104 heat-resistant strains, showed that only 24 of them were able to continuously maintain their acid production for pH variations ranging from 3 to 6 (data not showed). These pH values are those prevailing in cocoa beans fermentation within 24 to 72 h of fermentation (Kouame et al., 2015). Table 3 highlights the amounts of lactic acid produced by these 24 cocoa-specific acidifying LAB strains under pH fluctuation.

Regarding the rate of lactic acid produced by these Cocoa-specific acidifying LAB strains, values are ranged from a minimum of 0.03 to a maximum of 1.19 for pH values between 3 and 6 (Table 3). These findings pointed out the fact that these Cocoa-specific acidifying LAB strains did not have the same capacity of adaptation to temperature and pH fluctuations of the fermentative heaps even if they are able to maintain their lactic acid production in the required standards.
Additional activity of isolated Cocoa-specific acidifying LAB strains: citrate degradation and acetoin production Citrate degradation and acetoin production are important and relevant properties desirable for a good cocoa fermentation (Schwan and Wheals, 2004). Thus, biochemical characterization of the 24 Cocoa-specific acidifying LAB strains showed that they exhibited the property to break down citric acid. Additionally, only six (25%) of them displayed both the ability to degrade citrate and to produce acetoin.
The screening of citrate consumers among these cocoa-specific acidifying LAB strains indicates that all of them are able to breakdown citrate. Citrate metabolism constitutes, an important and particular property, since LAB strains are not usually able to utilize citric acid as carbon source (Kouame et al., 2015). As citrate metabolism is an essential factor for modulating the pH and promoting the growth of desirable bacterial flora in fermenting cocoa (Schwan and Wheals, 2004) all these 24 LAB strains should be responsible for the raising of the pH recorded during cocoa beans fermentation.
In addition, acetoin production shows that these six cocoa-specific acidifying LAB strains (T1Gb8, T6C5, T6C12, T7C8, T9C9 and T11C5) were able to metabolized citric acid via pathways pII producing acetoin from citrate. This ketone carbonylic compound (such as acetoin) is crucial for the development of good cocoa flavor. Indeed, Rodriguez-Campos et al. (2012) indicated that acetoin appears to be a precursor of tetramethylpyrazine, an important odor-active component of cocoa flavor. Therefore, these six LAB strains are of interest for the fermentation process as potential starter.
16S rRNA gene PCR and identification of isolates
Amplification of hypervariable regions (V1, V2 and V6) of the 16S gene of six potential starters LAB by colony PCR yielded a DNA fragment with respect to the 500bp (Figure 1). Analysis of these DNA fragments in the NCBI data bank led to the identification of these six LAB (T1Gb8, T6C5, T6C12, T7C8, T9C9 and T11C5) isolates as belonging to Lactobacillus plantarum with 99% of similarity. L. plantarum has been isolated from many cocoa fermentations around the world (Camu et al., 2007; Pereira et al., 2012).This observation indicates that, these strains play key roles in cocoa bean fermentation. L. plantarum species has also been used as starter for cocoa fermentation (Joko et al., 2014; Penia et al., 2013) and are widely used in food industies to make other fermented foods (Joko et al., 2014).

Influence of additional stress on lactic acid production by isolated Cocoa-specific acidifying LAB strains.
In addition to pH and temperature, other factors such as acids (acetic, lactic and citric) and alcohols (ethanol) are key parameters that may strongly influence the performance of the LAB strains in lactic acid production. Thus, the six selected LAB strains (T1Gb8, T6C5, T6C12, T7C8, T9C9 and T11C5) and identified as Lactobacillus plantarum were tested to assess the influence of these key parameters on Their ability to maintain lactic acid production.
Effect of acids (acetic and lactic acids) on lactic acid production
In acid conditions induced by lactic, acetic and citric acids, the six selected LAB strains proved to be able to maintain their potential of lactic acid production despite the observed decline. Lactic acid production occurred in medium containing lactic and acetic acid in the same range 0-1.4% for the six strains belonging to Lactobacillus plantarum (Figure 2 (A and B)) while this acid production continues up to 1.6% of acetic acid in the medium. The influence of citric acid on lactic acid production was lesser than the other tested acids (Figure 2C). Lactic acid production was slowly decreased with increasing of citric acid concentrations in a range of 0-3%. All Lactobacillus plantarum tested strains maintained their ability to produce lactic acid in proportions ranging from 35 to 66,96% of the initial level with a maximum of lactic acid (between 1.12 and 1.35%) produced under 0.5% of citric acid in the medium. The effect of acids results in a reduction of cytoplasmic pH that is likely to impair lactic acid production (Lopez and Dimick, 1995). This indicates that, compounds present at variable concentrations in the cocoa pulp such acids may be limiting factors for lactic acid production in LAB strains during cocoa fermentation. However, with their ability to maintain lactic acid production despite pH fluctuations between 3 and 6 in cocoa beans fermentation, all the six selected LAB strains should be particularly interesting as starters.
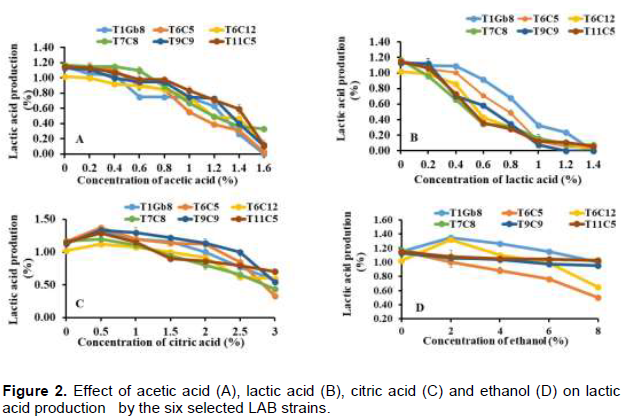
Effect of ethanol on lactic acid production
The influence of ethanol concentration on lactic acid production is shown in Figure 2D. The results suggested that, all these six studied strains maintained a good level of lactic acid production until 8% of ethanol concentration. This lactic acid poduction ranged between 43.50 and 91.67% of their initial potential production. These results are similar to those described by Behannis et al. (2012) with a decrease in acid produced when ethanol concentration increased from 2.5 to 7.5%. These results are interesting because ethanol is a major metabolite of cocoa pulp fermentation and the maximum rate of ethanol produced by yeasts is around 8% during this process (Lefeber et al., 2012). High concentrations of ethanol reduce cellular vitality and increase cell death resulting in a gradual decline in lactic acid production (Birch et al., 2000). Thus, these six ethanol tolerant isolates (T1Gb8, T6C5, T6C12, T7C8, T9C9 and T11C5) at high ethanol concentration, could both maintain their citrate and acetoin metabolism and their lactic acid production in the required standards to improve the final fermenting cocoa bean quality.these six studied strains maintained a good level of lactic acid production until 8% of ethanol concentration. This lactic acid poduction ranged between 43.50 and 91.67% of their initial potential production. These results are similar to those described by Behannis et al. (2012) with a decrease in acid produced when ethanol concentration increased from 2.5 to 7.5%. These results are interesting because ethanol is a major metabolite of cocoa pulp fermentation and the maximum rate of ethanol produced by yeasts is around 8% during this process (Lefeber et al., 2012). High concentrations of ethanol reduce cellular vitality and increase cell death resulting in a gradual decline in lactic acid production (Birch et al., 2000). Thus, these six ethanol tolerant isolates (T1Gb8, T6C5, T6C12, T7C8, T9C9 and T11C5) at high ethanol concentration, could both maintain their citrate and acetoin metabolism and their lactic acid production in the required standards to improve the final fermenting cocoa bean quality.
all these six studied strains maintained a good level of lactic acid production until 8% of ethanol concentration. This lactic acid poduction ranged between 43.50 and 91.67% of their initial potential production. These results are similar to those described by Behannis et al. (2012) with a decrease in acid produced when ethanol concentration increased from 2.5 to 7.5%. These results are interesting because ethanol is a major metabolite of cocoa pulp fermentation and the maximum rate of ethanol produced by yeasts is around 8% during this process (Lefeber et al., 2012). High concentrations of ethanol reduce cellular vitality and increase cell death resulting in a gradual decline in lactic acid production (Birch et al., 2000). Thus, these six ethanol tolerant isolates (T1Gb8, T6C5, T6C12, T7C8, T9C9 and T11C5) at high ethanol concentration, could both maintain their citrate and acetoin metabolism and their lactic acid production in the required standards to improve the final fermenting cocoa bean quality.
CONCLUSION
In this study, six LAB strains (T1Gb8, T6C5, T6C12, T7C8, T9C9 and T11C5) involved in Ivorian cocoa fermentation were identified as Lactobacillus plantarum. These particular cocoa-specific acidifiying bacteria presented both the required acidifying capacity and in a lesser extent, citrate degradation and acetoin production. All of them were able to maintain their lactic acid production under fermentative stress conditions in the required standards. That make them suit candidate as starter for cocoa fermentation control.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
This work was supported by a Ph.D. grant to the first author. The authors are gratful to the National Flowers Center of Felix Houphouët-Boigny University for their availability and support during the fermentations.
REFERENCES
|
Afoakwa OE, Kongor JE, Takrama JF, Budu AS (2013). Changes in acidification, sugars and mineral composition of cocoa pulp during fermentation of pulp preconditioned cocoa (Theobroma cacao) beans. International Food Research Journal, 20:1215-1222. |
|
|
Behannis M, Kayanush AJ (2012). Influence of Ethanol on Probiotic and Culture Bacteria Lactobacillus bulgaricus and Streptococcus thermophilus within a Therapeutic Product. Journal of Medical Microbiology, 2:70-76. |
|
|
Birch RM, Walker GM (2000). Influence of magnesium ions on heat shock and ethanol stress responses of saccharomyces cerevisiae. Enzyme Biochemical Technology, 26:678-687. |
|
|
Crafack M, Mikkelsen MB, Saerens S, Krudsen M, Blennow A, Lowor S, Nielsen DS (2013). Influencing cocoa flavour using Pichia kluyveri and kluyveromyces marxianus in a defined mixed starter culture for cocoa fermentation. International Journal of Food Microbiology, 167:103-116. |
|
|
Deheuvels O, Assiri AA, Petithuguenin P, Kebe BI, Flori A (2003). Production cacaoyere en cote d'ivoire : etat actuel du verger et pratiques paysannes –in: 14th International Cocoa Research Conference, Working paper, Accra (Ghana). |
|
|
Guha A, Soumitra B, Debabrata B (2013). Production of lactic acid from sweet meat industry waste by lactobacillus delbruki. International Journal of Research in Engineering and Technology, 2:630-634. |
|
|
ICCO 2016. International Cocoa Organization. |
|
|
Joko S, Ho E, Cahyaningsih, Betty SL Jenny (2014). Application of lactic acid bacteria to control microbial contaminants during fermentation of cocoa beans. International Journal of Research and Agricultural in Food Science, 2:16-24. |
|
|
Kempler GM, Kay Mc LL (1980). Improved medium for detection of citrate-fermenting streptococcus lactis subsp. Diacetylactis. Journal of Applied and Environmental Microbiology, 39: 927-956. |
|
|
King N (1948). Modification of Voges-Proskauer test for rapid colorimetric determination of acetyl methyl carbimol plus diacetyl in butter. Dairy Inductries, 13:860-866. |
|
|
Kouame LM, Doue GG, Adom NJ, Ouattara HG, Niamke SL (2015). Biochemical characterization of microbial populations involved in Loh-Djiboua cocoa's fermentation in Cote d'Ivoire. Food Environmental Safety, 14:196-205. |
|
|
Lefeber T, Papalexandratou Z, Gobert W, Camu N, De Vuyst L (2012). On-farm implementation of starter culture for improved cocoa bean fermentation and its influence on the flavour of chocolate produced thereof. Food of Microbiology, 30:379-392. |
|
|
Lopez AS, Dimick PS (1995). Cocoa fermentation. In: Reed, G., Nagodawithana, T. W. (Eds.), Enzymes, Biomass, Food and Feed. VCH, Weinheim. pp 561-577. |
|
|
Ouattara HG, Reverchon S, Niamke SL, Nasser W (2011). Molecular identification and pectate lyase production by Bacillus strains involved in cocoa fermentation. Food Microbiology, 28:1-8. |
|
|
Papalexendratou Z, Lefeber T, Bahrin Lee SO, Daniel HM, De Vuyst L (2013). Hanseniaspora Opuntiae, Saccharomyces Cerevisiae, Lactbacillus fermentum, and Acetobacter pasteurianus predominate during well-performed Malaysian cocoa bean box fermentation, underlining the importance of these microbial species for a successful cocoa bean fermentation process. Food Microbiology, 35:73-85. |
|
|
Pereira GVD, Miguel M, Ramos CL, Schwan RF (2012). Microbiological and physicochemical characterization of small scale cocoa fermentations and screening of yeast and bacterial strains to develop a defined starter culture. Applied and Environmental Microbiology, 78:5395-5405. |
|
|
Raimbiault M (1995). Importance des bactéries lactiques dans les fermentations du manioc. In Transformation Alimentaire du Manioc. T. Agbor Egbe, A. Brauman, D. Griffon, S. Trèche (éd ORSTOM). |
|
|
Rodriguez-Campos J, Escalona-Buendia HB, Contreras-Ramos SM, Orozco-Avila I, Jaramillo-Flores ME, Lugo-Cervantes (2012). Effects of fermentation time and drying temperature on volatile compounds in cocoa. Food Chemistry, 132:277-288. |
|
|
Serra BJ, Ventura CF (1997). Evaluation of bitterness and astringency of polyphenolic compounds in cocoa powder. Food Chemistry, 60:365-370. |
|
|
Schwan RF, Wheals AE (2004). The microbiology of cocoa fermentation and its role in chocolate quality. Critical Reviews in Food Science and Nutrition, 44:205-221. |
|
|
Yao A (2009). La fermentation du manioc en gari dans l'Afrique de l'Ouest : production d'un starter de bactéries lactiques lyophilisées. Thèse de Doctorat Unique, Université de Liège, 213 p. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


