ABSTRACT
The emergence of bacterial resistance to antibiotics is a serious challenge to the global health system. The QS inhibition is one of the approaches to fight against antibiotic resistance in bacteria. Securidaca longepedunculata is a medicinal plant that roots are the only organ used against microbial diseases. This species is threatened with extinction due to the massive use of its roots in phytotherapy. In this study, the anti-QS and anti-biofilm activities of S. longepedunculata leaves methanolic extract at 100 to 400 µg/mL was assessed against the bacterial strains Chromobacterium violaceum CV026 and Pseudomonas aeruginosa PAO1. The results showed anti-QS and anti-biofilm activity of S. longepedunculata leaves which reduced violacein production in C. violaceum CV026 by 12 to 59%. The virulence factor pyocyanin in P. aeruginosa PAO1 was inhibited from 13 to 46%. Biofilm formation was significantly inhibited (41%) at 400 µg/mL.
Key words: Securidaca longepedunculata, anti-quorum, anti-biofilm.
National Institutes of Health (NIH) estimated that more than 80% of human microbial infections are associated with bioï¬lms which is present in about 65% of chronic infections (Jamal et al., 2018). Biofilms are aggregations of microorganisms which live in an extracellular matrix. This matrix is composed by extracellular polymeric substances (EPS), including polysaccharides, nucleic acids, proteins and lipids, at a liquid interface (Li et al., 2020). In biofilms, the bad penetration of antibiotic, reduced nutrition and growth, adaptive stress responses, and persister cells formation would constitute a multi-layered defense system (Stewart, 2002). Cells growing in the biofilm are resistant to both antibiotic therapy and the host immune defense system. This situation is the cause of recurrent and recalcitrant infections (Li et al., 2020). Bacteria attached to a surface and growing in a biofilm are protected against the action of antibiotics, biocides and other control methods (Abebe, 2020). Persistent cells are a subpopulation of bacteria which can transiently survive under the lethal effect of antibiotic treatment and thus contribute to the strong resistance of biofilms (Dincer et al., 2020).
Bacteria control the expression profile of genes according to the size of the microbial population through the mechanism of quorum sensing (QS), which allows to form different forms of biofilm (Abebe, 2020). QS acts on the development of the biofilm and controls the production of virulence factors (enzymes, toxins) necessary for resistance to phagocytosis (Li et al., 2020). One of the strategies proposed for overcoming resistance in biofilm is the use of QS inhibitors (Li et al., 2020).
Pseudomonas aeruginosa is an opportunistic bacterium responsible for chronic infections which occurred in hospitals especially in patients suffering from cystic fibrosis. The biofilm formation and the production of several virulence factors characterize the infection process. At the same time, the sessile growth mode leads to a reduction of bacterial sensitivity to both host defenses and antimicrobial agents. Quorum sensing, which is the inter-bacterial communication system, is correlated with their biofilm production and resistance (Ciofu and Tolker-nielsen, 2019).
Medicinal plants offer a variety of phytochemicals with a new potential control of microbial diseases, due to the spectrum of secondary metabolites present in the extracts, which include phenolic, quinones, flavonoids, alkaloids and terpenoids (Asfour, 2018).
As a result, the commonly used plants in ethnomedicine constitute an alternative to search bioactives compounds against virulence factors or their production.
Securidaca longepedunculata is a highly medicinal plant whose roots and stem bark are the only organs used against bacterial disease and for the treatment of chronic wounds in traditional medicine (Mongalo et al., 2015). This plant is threatened because of the massive use of its roots (Compaoré et al., 2018).
This study aimed to assess anti-QS and anti-biofilm activities of S. longepedunculata leave extract.
Bacterial strains and growth conditions
The Plant Biotechnology Laboratory of Université Libre de Bruxelles (Belgium) has kindly offered P. aeruginosa PAO1 and Chromobacterium violaceum CV026 strains for the biological activities assessment. Luria-Bertani (LB) broth was used as culture medium (37°C for PAO1 and 30°C for CV026).
Plant material collection and extraction
The leaves of S. longepedunculata were harvested in Badara locality (Region of "Haut Bassin"). Voucher specimen (CI : 16713) was identified and deposited in the herbarium of "Université Joseph KI ZERBO", Burkina Faso. The powder from dried samples was extracted with methanol for one day. An evaporator was used to concentrate the extracts under vacuum before drying them.
Assessment of inhibition of violacein production in C. violaceum CV026
The method of Choo et al. (2006)was used to assess the ability of S. longepedunculata extract to affect negatively the QS system through its effect on violacein production in C. violaceum CV026. In the presence of exogenous N-hexanoyl-L-homoserine lactone (C6-HSL; Sigmaâ€Aldrich), C. violaceum CV026 which is a mutant strain is capable of producing violacein. Briefly, to the mixture of S. longepedunculata extract (in DMSO) and C6-HSL, a diluted culture of C. violaceum CV026 (18 h at 30°C) was added. The final concentrations were 100 to 400 µg/mL for the extracts and 10 µM for C6-HSL. After tubes incubation at 30°C (24 h, 175 rpm), bacterial growth was assessed by measuring the bacterial turbidity (OD600 nm). The bacterial culture (1 mL) was first centrifuged (7000 rpm, 10 min) and then the violacein was dissolved in 1 mL of DMSO. Quantification of violacein production was performed by measuring its absorbance at 585 nm.
Inhibition of pyocyanin production in P. aeruginosa PAO1
The QS controls the pyocyanin production. The inhibitory capacity of S. longepedunculata leaves extract on this production was assessed according to the method of Ouedraogo and Kiendrebeogo (2016). Briefly, the extract was used to make a series (dilution by half) of concentrations (in DMSO) which were each added overnight to a culture of P. aeruginosa PAO1. The final concentrations of the extracts were 100 to 400 µg/mL. Bacterial growth was assessed by measuring the bacterial turbidity (OD600nm) after 18 h incubation (37°C, 175 rpm) of tubes. To assess the production of pyocyanin (A380), the supernatant was used.
Biofilm formation and quantification
The ability of S. longepedunculata extract to inhibit biofilm formation was assessed according to the method of Vandeputte et al. (2010). A volume of 200 µL of P. aeruginosa PAO1 culture was supplemented with leaves extract solution for final concentrations ranging from 100 to 400 µg/mL in round-bottomed wells. The supernatant was removed after 24 h incubation at 37°C and the biofilm was fixed with methanol after washing with distilled water. A crystal violet solution (0.1% in water) was added to the wells followed by 30 min incubation at room temperature. The crystal violet stained was dissolved with 200 µL of acetic acid (33% in water) in order to read solution absorbance at 590 nm.
Statistical analysis
One way analysis of variance (ANOVA) followed by Tukey test of Graph Pad Prism software was used to determined statistical significance; p value ≤ 0.05 was considered significant (n=3).
Anti-QS activity of S. longepedunculata leaves
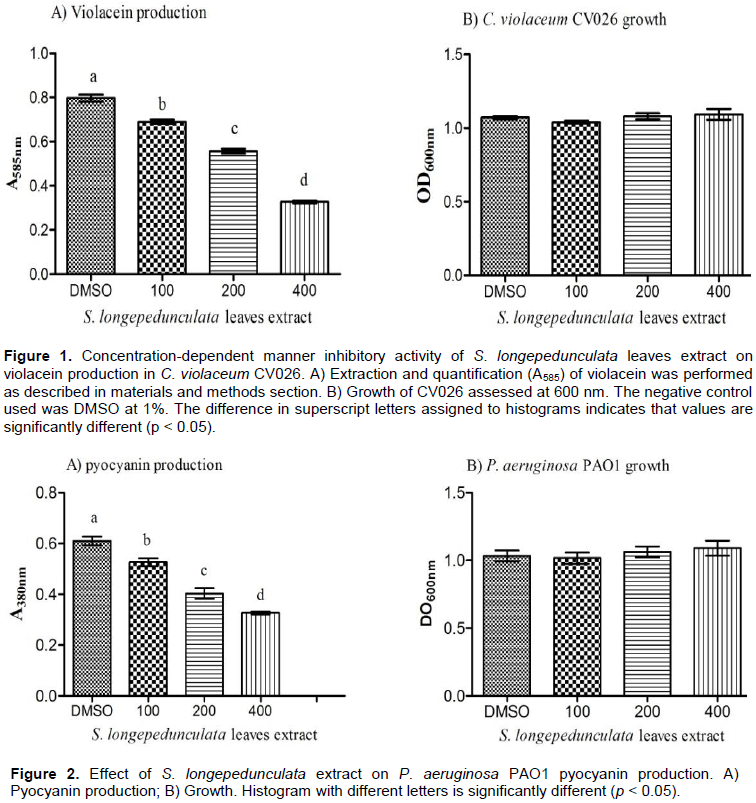
To search anti-QS compounds, the strain of C. violaceum CV026 is indicated. The leaves extract at different concentrations (100-400 µg/mL) was used to assess the inhibitory capacity of S. longepedunculata on violacein production after 24 h of growth. Figure 1A shows that compared to the control (DMSO at 1%), the production of violacein is reduced by 12 to 59% by S. longepedunculata leaves extract in concentration-dependent manner. The growth of C. violaceum CV026 is not affected by this reduction (Figure 1B). These results confirm that S. longepedunculata leaves extract contains anti-QS compounds.
S. longepedunculata leaves extract affects QS-controlled extracellular virulence factor production
The leaves extract reduced significantly the production of violacein, thus showing an inhibitory effect of S. longepedunculata on QS system. The QS system in P. aeruginosa controls the production of pyocyanin which is the virulence factor. The redox cycle of host cells is altered by a blue-green phenazine pigment which is the pyocyanin produced in culture medium (Liu and Nizet, 2009). The assessment of S. longepedunculata ability to inhibit pyocyanin production showed significant reduction effects ranging from 13 to 46% in dose-dependent manner (100 to 400 µg/mL) of leaves extract (Figure 2A). The growth of P. aeruginosa PAO1 was not affected by the extracts (Figure 2B).
S. longepedunculata leaves extract affect biofilm formation
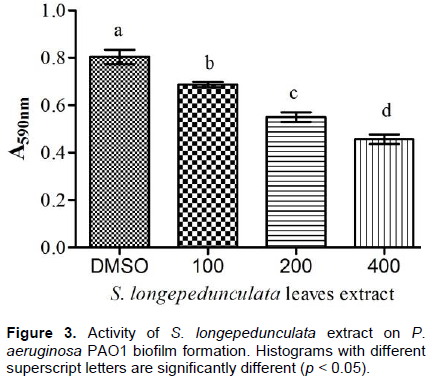
The formation of P. aeruginosa biofilm PAO1 is related to QS system (Jimenez et al., 2012). Based on anti-QS activity observed, the ability of S. longepedunculata leaves extract to inhibit biofilm formation of P. aerugionosa PAO1 was evaluated. Figure 3 shows that biofilm formation is significantly inhibited by S. longepedunculata leaves extract at different concentrations (100-400 µg/mL). An inhibition of 41% was recorded at the concentration of 400 µg/mL.
Bacterial biofilms are difficult to control and show high resistance to antibiotics (Koo et al., 2017). The destruction of a fully formed biofilm necessarily involves the use of molecules capable of penetrating its structure or destructuring it (Paluch et al., 2020).
This study showed that S. longepedunculata leaves methanolic extract have anti-QS activity, inhibit virulence factors and biofilm formation. High concentrations of S. longepedunculata extract did not show bactericidal activity against C. violaceum and P. aerugionosa. This absence of bactericidal activity shows that the leaves of S. longepedunculata act only through their inhibitory effects on QS and the formation of the biofilm. An inhibition of biofilm formation would make bacteria accessible and sensitive to the immune system and to antibacterial. This result would suggest a possible combination of S. longepedunculata leaves and some antimicrobial compounds in external treatment. Previous work has reported that leaves of S. longepedunculata are rich in polyphenol, flavonoids, and alkaloids (Karama et al., 2018). The biological activities of S. longepedunculata observed in this work are obviously due to the nature of bioactive compounds contained in its leaves. Vasavi et al. (2016)reported that plant flavonoids have the ability to interfere with inter-microbial communication and have anti-biofilm activity. Karama et al. (2020)reported the presence of compounds such as quercetin, chrysin, rutin, isorhamnetin, luteolin, gallic acid, ellagic acid, ferulic acid and tannic acid in leaves of S. longepedunculata. The anti-QS activity against C. violaceum of various flavonoids such as quercetin and luteolin have been reported (Bali et al., 2019). Rekha et al. (2016)reported that quercetin showed good inhibition of pyocyanin production.
The anti-biofilm activity of rutin against multidrug-resistant P. aeruginosa has been reported(Deepika et al., 2018). Wang et al. (2017)showed that rutin significantly inhibited the biofilm formation of Streptococcus suis without impairing its growth in vitro.
Likewise, the anti-QS and anti-biofilm effect of luteolin and chrysin have been reported against Gram-positive and Gram-negative bacteria (Cho et al., 2015; Shen et al., 2014).
Another study showed quercetin activity against virulence factors production and biofilm formation of P. aeruginosa PAO1 (Ouyang et al., 2016). Significant inhibitions were recorded with quercetin against pyocyanin, protease and elastase production and biofilm formation (Quecan et al., 2019). Other work had reported that quercetin inhibited the production of violacein in C. violaceum 12472, at 50 and 100 µg/mL, respectively (Vasavi et al., 2014).
In addition, Vikram et al. (2010)have shown that quercetin removes Escherichia coli O157: H7 and Vibrio harveyi biofilm formation. Tannic acid has reduced the QS regulated violacein production up to 47.7% (Sivasankar et al., 2019). According to a past study (Karama et al., 2018), the leaves of S. longepedunculata showed an alkaloid content of 245 µg/g of methanolic extract. Studies reported that alkaloids have shown the ability to reverse biofilm resistance (Othman et al., 2019; Su et al., 2020).
The anti-QS and biofilm formation inhibitory activity of S. longepedunculata leaves extract would be due to different bioactive compounds it contains. The results observed with S. longepedunculata extracts show that this plant could be used against microbial infections of P. aeruginosa. However, subsequent studies could accurately identify anti-QS and biofilm inhibitor molecules through bioguided screening. As S. longepedunculata is threatened with extinction, the leaves could be used instead of the roots in phytotherapy and thus contribute to a sustainable use of this plant in the management of microbial disease. The biological activity observed with the extract of S. longepedunculata would be due to the properties of anti-quorum sensing and anti-biofilm compounds in the phytochemistry of its leaves.
This study showed that methanolic extracts from S. longepedunculata leaves have the ability to inhibit significantly the production of QS-controlled factors including violacein and pyocyanin from C. violaceum and P. aeruginosa, respectively and the formation of biofilm. The extract of S. longepedunculata did not affect negatively the growth of bacteria used.
S. longepedunculata leaves would act only by inhibiting QS and biofilm formation. The anti-QS activity of leaves shows that they could be used in the treatment of P. aeruginosa infections. This work results show that S. longepedunculata leaves would contain anti-QS and anti-biofilm compounds that could be identified by further studies.
The authors have not declared any conflict of interests.
REFERENCES
|
Abebe GM (2020). The Role of Bacterial Biofilm in Antibiotic Resistance and Food Contamination. International Journal of Microbiology pp. 1-10.
Crossref
|
|
|
|
Asfour HZ (2018). Anti-Quorum Sensing Natural Compounds. Journal of Microscopy and Ultrastructure 6(1):1-10.
Crossref
|
|
|
|
|
Bali EB, Türkmen KE, Erdönmez D, Saglam N (2019). Comparative study of inhibitory potential of dietary phytochemicals against quorum sensing activity of and biofilm formation by Chromobacterium violaceum 12472, and swimming and swarming behaviour of Pseudomonas aeruginosa PAO1. Food Technology and Biotechnology 57(2):212-221.
Crossref
|
|
|
|
|
Cho HS, Lee JH, Cho MH, Lee J (2015). Red wines and flavonoids diminish Staphylococcus aureus virulence with anti-biofilm and anti-hemolytic activities. Biofouling 31(1):1-11.
Crossref
|
|
|
|
|
Choo JH, Rukayadi Y, Hwang JK (2006). Inhibition of bacterial quorum sensing by vanilla extract. Letters in Applied Microbiology 42(6):637-641.
|
|
|
|
|
Ciofu O, Tolker-nielsen T (2019). Tolerance and resistance of Pseudomonas aeruginosa biofilms to antimicrobial agents - How P . aeruginosa can escape antibiotics. Frontiers in Microbiology 10:1-15.
Crossref
|
|
|
|
|
Compaoré M, Meda RN, Zerbo P, Karama I, Traoré O, Lamien-meda A, Kiendrebeogo M, Novak J (2018). Availability Evaluation of Twelve Antimalarial Medicinal Plants from Western Regions of Burkina Faso. Journal of Diseases and Medicinal Plants 4(3):80-88.
Crossref
|
|
|
|
|
Dincer S, Uslu FM, Delik A (2020). Antibiotic Resistance in Biofilm.In Bacterial Biofilms, Dincer S, Özdenefe MS, Arkut A, IntechOpen 1-360.
Crossref
|
|
|
|
|
Jamal M, Ahmad W, Andleeb S, Jalil F, Imran M, Asif M, Hussain T, Ali M, Rafiq M, Atif M (2018). Bacterial biofilm and associated infections. Journal of the Chinese Medical Association 81(1):7-11.
Crossref
|
|
|
|
|
Jimenez PN, Koch G, Thompson JA, Xavier KB, Cool RH, Quax WJ (2012). The Multiple Signaling Systems Regulating Virulence in Pseudomonas aeruginosa. Microbiology and Molecular Biology Reviews 76(1):46-65.
Crossref
|
|
|
|
|
Karama I, Compaoré M, Djandé A, Traoré O, Lagnika L, Kiendrebeogo M (2020). Antiplasmodial Activities of Flavonoids from Leaves of Securidaca longepedunculata Fresen (Polygalaceae). American Journal of BioScience 8(1):1-5.
Crossref
|
|
|
|
|
Karama I, Compaoré M, Traore O, Sanon S, Lagnika L, Kiendrebeogo M (2018). Comparative study of leaves and root bark from Securidaca longepedunculata Fresen (Polygalaceae): phytochemistry and antiplasmodial activity. The Pharma Innovation Journal 7(10):178-184.
|
|
|
|
|
Koo H, Allan RN, Howlin RP, Stoodley P, Hall-Stoodley L (2017). Targeting microbial biofilms: Current and prospective therapeutic strategies. Nature Reviews Microbiology 15(12):740-755.
Crossref
|
|
|
|
|
Li Y, Xiao P, Wang Y, Hao Y (2020). Mechanisms and control measures of mature biofilm resistance to antimicrobial agents in the clinical context. ACS Omega 5(36):22684-22690.
Crossref
|
|
|
|
|
Liu GY, Nizet V (2009). Color me bad : microbial pigments as virulence factors. Trends in Microbiology 17(9):406-13.
Crossref
|
|
|
|
|
Mongalo NI, McGaw LJ, Finnie JF, Staden JV (2015). Securidaca longepedunculata Fresen (Polygalaceae): A review of its ethnomedicinal uses, phytochemistry, pharmacological properties and toxicology. Journal of Ethnopharmacology 165:215-226.
Crossref
|
|
|
|
|
Othman L, Sleiman A, Abdel-Massih RM (2019). Antimicrobial activity of polyphenols and alkaloids in Middle Eastern plants. Frontiers in Microbiology 10:1-28
Crossref
|
|
|
|
|
Ouedraogo V, Kiendrebeogo M (2016). Methanol extract from Anogeissus leiocarpus (DC) Guill . et Perr . ( Combretaceae ) stem bark quenches the Quorum Sensing of Pseudomonas aeruginosa PAO1. Medicines 3(4):26.
Crossref
|
|
|
|
|
Ouyang J, Sun F, Feng W, Sun Y, Qiu X, Xiong L, Liu Y, Chen Y (2016). Quercetin is an effective inhibitor of quorum sensing, biofilm formation and virulence factors in Pseudomonas aeruginosa. Journal of Applied Microbiology 120(4):966-974.
Crossref
|
|
|
|
|
Paluch E, Rewak-Soroczyńska J, Jędrusik I, Mazurkiewicz E, Jermakow K (2020). Prevention of biofilm formation by quorum quenching. Applied Microbiology and Biotechnology 104(5):1871-1881.
Crossref
|
|
|
|
|
Quecan BX, Santos JT, Rivera ML, Hassimotto NM, Almeida FA, Pinto UM (2019). Effect of quercetin rich onion extracts on bacterial quorum sensing. Frontiers in Microbiology 10:867.
Crossref
|
|
|
|
|
Rekha PD, Vasavi HS, Vipin C, Saptami K, Arun AB (2016). A medicinal herb Cassia alata attenuates quorum sensing in Chromobacterium violaceum and Pseudomonas aeruginosa. Letters in Applied Microbiology 64(3):231-238.
Crossref
|
|
|
|
|
Deepika MS, Thangam R, Sakthidhasan P, Arun S, Sivasubramanian S, Thirumurugan R (2018). Combined effect of a natural flavonoid rutin from Citrus sinensis and conventional antibiotic gentamicin on Pseudomonas aeruginosa biofilm formation. Food Control 90:282-294.
Crossref
|
|
|
|
|
Shen XF, Ren LB, Teng Y, Zheng S, Yang XL, Guo XJ, Wang XY, Sha KH, Li N, Xu GY, Tian HW, Wang XY, Liu XK, Li J, Huang N (2014). Luteolin decreases the attachment, invasion and cytotoxicity of UPEC in bladder epithelial cells and inhibits UPEC biofilm formation. Food and Chemical Toxicology 72:204-211.
Crossref
|
|
|
|
|
Sivasankar C, Jha NK, Ghosh R, Shetty PH (2019). Anti quorum sensing and anti virulence activity of tannic acid and it 's potential to breach resistance in Salmonella enterica Typhi / Paratyphi A clinical isolates. Microbial Pathogenesis 138:103813.
Crossref
|
|
|
|
|
Stewart PS (2002). Mechanisms of antibiotic resistance in bacterial biofilms. International Journal of Medical Microbiology 292(2):107-13.
Crossref
|
|
|
|
|
Su T, Qiu Y, Hua X, Ye B, Luo H, Liu D, Qu P, Qiu Z (2020). Novel Opportunity to Reverse Antibiotic Resistance: To Explore Traditional Chinese Medicine With Potential Activity Against Antibiotics-Resistance Bacteria. Frontiers in Microbiology 11:1-11.
Crossref
|
|
|
|
|
Vandeputte OM, Kiendrebeogo M, Rajaonson S, Diallo B, Mol A, Jaziri M, Baucher M (2010). Identification of catechin as one of the flavonoids from Combretum albiflorum bark extract that reduces the production of quorum-sensing-controlled virulence factors in Pseudomonas aeruginosa PAO1. Applied and Environmental Microbiology 76:243-253.
Crossref
|
|
|
|
|
Vasavi HS, Arun AB, Rekha PD (2014). Anti-quorum sensing activity of Psidium guajava L. flavonoids against Chromobacterium violaceum and Pseudomonas aeruginosa PAO1. Microbiology and Immunology 58(5):286-293.
Crossref
|
|
|
|
|
Vasavi HS, Arun AB, Rekha PD (2016). Anti-quorum sensing activity of flavonoid-rich fraction from Centella asiatica L. against Pseudomonas aeruginosa PAO1. Journal of Microbiology Immunology and Infection 49(1):8-15.
Crossref
|
|
|
|
|
Vikram A, Jayaprakasha GK, Jesudhasan PR, Pillai SD, Patil BS (2010). Suppression of bacterial cell-cell signalling, biofilm formation and type III secretion system by citrus flavonoids. Journal of Applied Microbiology 109(2):515-527.
Crossref
|
|
|
|
|
Wang S, Wang C, Gao L, Cai H, Zhou Y, Yang Y, Xu C, Ding W, Chen J, Muhammad I, Chen X, He X, Liu D, Li Y (2017). Rutin inhibits Streptococcus suis biofilm formation by affecting CPS biosynthesis. Frontiers in Pharmacology 8:379.
Crossref
|
|