Full Length Research Paper

ABSTRACT
The crystal (Cry) proteins produced by Bacillus thuringiensis determine a particular strain's toxicity profile. This study was focused on cloning a cry1Ia gene based on amplicon restriction fragment length polymorphism (ARFLP) profile which would be helpful in developing new biopesticides with broader and higher spectrum of toxicity against Lepidoptera and Coleoptera insect pests. The present paper describes cry1Ia gene from a local isolate of Bacillus thuringiensis (B.t) CFE20(3). A PCR-restriction fragment length polymorphism method for identification of cry1I-type genes from Bacillus thuringiensis was established by designing a pair of primers based on the conserved regions of the genes to amplify 2,169 bp cry1I-type gene fragments. Amplification products were digested with the KpnI and XbaI, and new kind of cry1I-type genes was successfully identified. Escherichia coli DH5α was transformed with recombinant DNA comprising pTZ57R/T and Bt cry1I (2169 bp) amplified from a native isolate CFE20(3) for cloning. The cloned 2169 bp was sequenced and then ligated in the expression vector pQE30 for transformation of E. coli M15 and SG13009 for expression analysis. The sequence obtained shows 99% homology with known cry1Ia from B. thuringiensis subsp. Kurstaki. An expected band size of 81 KDa was observed after sodium dodecyl sulphate - polyacrylamide gel electrophoresis (SDS-PAGE) analysis indicating the expression of cry1I. The toxicity of crude recombinant Cry1I proteins was determined against third instar larvae of Diamond back moth, Plutella xylostella and Spodoptera litura. Cry1I protein was found to be effective against Plutella xylostella.
Key words: Bacillus thuringiensis, cry1I, SDS-PAGE, Plutella xylostella, Spodoptera litura.
INTRODUCTION
Crystal proteins from the Gram-positive spore-forming bacterium, Bacillus thuringiensis are toxic to a wide variety of insects that are economically important as pests. Many different genes encoding the B. thuringiensis endotoxin have been isolated and characterized. The genes have been classified as cry1 to cry72, cyt1, cyt2 and cyt3 and are ranked according to their homology. The 72 groups of cry genes are divided into class and subclasses according to their amino acid similarities (Crickmore et al., 1998; Song et al., 2003; Pooja et al., 2013; B. thuringiensis toxin nomenclature website at http://www.biols.susx.ac.uk/home/Neil_Crickmore /Bt/). Cry1 proteins that are active against lepidopteran insects are produced as crystalline parasporal inclusions during sporulation. Generally, the crystals are composed of pro-toxins of approximately 130 kDa, but cry1I-type genes are usually silent genes capable of encoding a protein of about 81 kDa in B. thuringiensis strains (Choi et al., 2000; Gleave et al., 1993). We decided to screen B. thuringiensis isolates for cry1I genes with the aim of finding novel cry1I genes, which could encode insecticidal proteins toxic to insensitive or resistant insect pests (Song et al., 2003).
The sporulating cells of Bt release crystal proteins into the surrounding medium and after exposure to alkaline environment the crystal protein is activated. The C terminal end of the protein recognizes the receptor site in the gut membrane and then its N terminal end undergoes conformational changes and loop like structure is formed which ultimately insert in the membrane and make pores. The lysis of the cells due to the formation of non specific pores causes the paralysis of the gut and the larvae stop feeding. This brings about larval death (Schwartz et al., 1993; Lorence et al., 1995; Pietrantonio and Gill, 1996).
The Cry proteins are classified based on similarities among amino acid sequences, and are grouped into 72 classes. There are more than 500 different cry genes that encode these proteins that may be done using the polymerase chain reaction technique. This technique has been applied in several studies, including to find new isolates with entomopathogenic potential. The type of cry gene present in an isolate correlates with the specificity of the produced Cry protein (Lamptey et al., 1991).
Some of these proteins have toxicities to more than one insect order; for example, Cry1I is toxic to the Lepidoptera and Coleoptera orders (Tailor et al., 1992). Other cry1I genes have been characterized and published (Tounsi et al., 2003; Selvapandiyan et al., 1998; Gleave et al., 1993; Choi et al., 2000; Bergamasco et al., 2011; Pooja et al., 2013). The cry1I genes encode proteins of about 70 to 80 kDa that are segregated during the vegetative phase of B. thuringiensis and do not accumulate as crystals (Grossi-de-Sa et al., 2007).
With advances in biotechnology, genes that encode proteins with insecticidal characteristics may be isolated from B. thuringiensis, cloned into Escherichia coli or B. thuringiensis mutants for cry genes (cry-B) and even modified to express adequately in plants. Bt cotton plants have already been adopted by some farmers, which has decreased insecticidal application by 50 to 65% (Bergamasco et al., 2011; Santos et al., 2003). The objec-tives of the present work were to clone the cry1Ia gene of a native isolate into an expression vector, express the Cry1Ia protein in E. coli and evaluate its insecticidal activity against Plutella xylostella and Spodoptera litura.
MATERIALS AND METHODS
Bacillus thuringiensis strains, plasmids
Native Bacillus thuringiensis isolate CFE20(3) and CFE25(2) which are available in Institute of Agri Biotechnology, University of Agricultural Sciences, Dharwad was used to amplify cry1Ia gene. HD1 was used as a reference strain. The T/A cloning vector pTZ57R/T was obtained from InsTA clone PCR cloning kit #K1213, Fermentas and pQE30 from Qiagen (Cat. No. 32915), the host E. coli JM109 for maintenance and E. coli M15 and SG13009 for expression analysis from Qiagen.
Amplification of cry1I gene
Gene specific primer which was synthesized at Sigma Aldrich Pvt. Ltd., Bangalore, was used for amplification of cry1I gene. The forward and reverse primers used were 5’GGATCCATGAAACTAAAGAATCAAGATAAGC3’ and 3’CTGCAGCATGTTACGCTCAATATGGAGT 5’, respectively. PCR was performed with 3U Taq DNA polymerase, 1 mM dNTP, 5 pM primer each, 25 mM MgCl2 in a final volume of 100 μl. Amplification was done in an Eppendorf thermal cycler under the following conditions: 5 min of denaturation at 94°C followed by 35 cycles of amplification with a 1 min denaturation at 94°C, 1 min of annealing at 50.4°C, 2 min of extension at 72°C, final extension step of 45 min at 72°C.
Amplicon restriction fragment length polymorphism (ARFLP)
The amplified product of cry genes from the potent isolates was used for checking the variants by restriction analysis on the basis of ARFLP of the PCR amplified DNA as outlined by Kuo and Chak (1996). The purified
Molecular cloning and nucleotide sequencing
PCR amplified products were ligated to the T/A cloning vector pTZ57R/T (Sambrook and Russell, 2001) using the Fermentas DNA ligation kit. The transformed cells were spread on LB agar plates containing X-gal (20 mg/ml), Isopropyl-β-D-thiogalactopyranoside (IPTG) (24 mg/ml) and ampicillin (100 μg/ml). The plates were then incubated at 37°C for 12 to 16 h and the transformed colonies were further streaked on Luria agar with ampicillin (100 μg/ml). The confirmation for the presence of desired DNA fragment in cloning vector was done by PCR using gene specific primers and by restriction analysis, and subsequently, gel electrophoresis carried was out. Nucleotide sequencing was done by using M13 forward and reverse primer at Chromous Biotech Pvt. Ltd., Bangalore. In order to express the cry1I gene, the construct containing cry1I was inserted into the multiple cloning site of an expression vector pQE30 to generate the recombinant expression construct. The complete amplified gene was gel purified using the Mini Elute PCR purification kit (Qiagen) according to the manufacturer’s instruct-tions. The insert sequence and its reading frame were confirmed by BamHI and PstI digestion. The ligated product was first transferred into E. coli JM109 cells for maintenance and then into E. coli M15 (pREP4) (Qiagen) and E. coli SG13009 (pREP) (Qiagen) for expression analysis. For confirmation of the clones, the plasmid was isolated by using alkaline lysis protocol of Birmboim and Doly (1979) and restriction analysis was done for the plasmids of selected clones by using BamHI and PstI restriction endo-nucleases.
Protein analysis and expression studies
For protein analysis, about 5 ml of Luria broth with kanamycin (50 mg/ml) and ampicillin (50 mg/ml), the protein was extracted and analyzed by sodium dodecyl sulphate polyacrylamide gel electrophoresis inoculated with a colony of E. coli containing the recombinant construct and incubated at 37°C overnight under shaking conditions. Overnight grown culture was diluted in fresh Luria broth in 1:100 ratio without selection pressure and incubated at 37°C until the culture reached the log phase of growth (A550-0.5 to 1.0) under shaking conditions which took approximately 3 h. The expression of target protein was induced based on the optimal values of IPTG (1 mM) concentration and it was again incubated for 5 h at 37°C in a shaker. For extraction of proteins, the cell culture was centrifuged at 13,000 rpm for 1 min at room temperature. The pellet was resuspended in 100 μl of T10E1 and 100 μl of 1X SDS gel loading buffer was added to it. The mixture was heated at 90°C on a thermo mixer and centrifuged for 10 min at 4°C. The supernatant was collected in micro centrifuge tubes and protein was quantified by using NanoDrop. The protein preparations were analyzed by SDS-PAGE as described by Sambrook and Russell (2001).
Mass multiplication of Plutella xylostella and Spodoptera litura
P. xylostella
Diamondback moth was mass multiplied in the laboratory by following the method described by Liu and Sun (1984) with minor modifications. The larvae were collected from the infested cabbage field and were reared separately on cabbage leaves which were raised in the green house under insecticide free conditions. Pupae obtained were kept in Petri plate and placed in a cage of 25 cm3 for adult emergence. When the moths started to emerge, mustard seedlings were provided for oviposition. Plastic cups of 6 cm height and 4.5 cm diameter were filled with sterilized vermiculite to a depth of 4 cm and pre-soaked mustard seeds (24 h) treated with Bavistin (2 g/kg) were sown in cups and allowed to germinate under natural conditions. Four to five days after germination, the cups were placed in the oviposition cage and replenished at 24 h interval. The moths laid eggs on both sides of cotyledons. The cups with eggs were transferred to plastic tubs (45 x 30 x 15 cm) for mass rearing. Ten percent honey solution containing the multivitamin/multimineral capsule, Becadexamin from GlaxoSmithkline Pharmaceuticals Limited was provided for the adults as food through cotton swab kept in a sterilized petriplate. For raising the mustard seedlings and rearing of DBM larvae, approximately 12:12 (L:D) photoperiod and 27±2°C temperature was maintained under laboratory conditions.
Eggs hatched in 2-3 days and neonates mined the mustard cotyledons and fed on them. When the seedlings were completely consumed, larvae were transferred to fully expanded cabbage leaves with petiole covered in wet cotton swab to maintain leaf turgidity. The third instar larvae (0.5 ± 0.15 cm length; 1.65 ± 0.20 mg weight) were used for the bioassay.
S. litura
The larvae collected from the infested fields of cabbage were reared separately on cabbage leaves raised in green house under insecticidal free condition. Pupae thus obtained were kept in a sterilized Petri plate and placed in the cage of 25 cm3 for adult emergence. When the moth started emerging, 25-30 days old small cabbage heads were provided for oviposition. The moth laid eggs both on ventral and dorsal surface of leaves, leaves with eggs were transferred to plastic tubs (45 x 30 x 15 cm) for mass rearing. Ten percent of honey solution containing the multivitamin/multimineral capsule, Becadexamin from GlaxoSmithkline Pharmaceuticals Limited was provide as food for adults in sterilized vial with cotton plug. The 3rd instar F1 generation larvae were used for bioassay.
Bioassay of cry1I clones
10 ml of Luria Bertani broth was inoculated with E. coli containing cry1I construct and kept in shaker overnight at 37°C. 1:100 dilution was made for cry1I construct , M15 host and plain pQE30 vector and keep in shaker till it reaches log phase. IPTG was induced (1 mM) of cry1I construct, M15 host and plain pQE30 vector. Cells were pelleted at 5000 rpm for 15 min at 4°C and supernatant was discarded. Cells were resuspended in binding buffer (20 mM sodium phosphate + 500 mM NaCl). Lysozyme was added at the final concentration of 1 mg/ml and incubated on ice for 30 min. Triton X100 was added to the above suspension (1%) and incubated on 4°C for 10 min on a rocking platform. Cell suspension was centrifuged at 5000 rpm for 30 min at 4°C and supernatant was collected in fresh tube. A constant concentration of protein was used for bioassay of P. xylostella and S. litura.
Insecticidal activity against third instar larvae of P. xylostella and S. litura was measured which was conducted on fresh leaf disks by leaf dip bioassays. Disks cut from leaves of cabbages incorporate a suspension of purified inclusions of cry1I clones. Toxicity studies on larvae of the P. xylostella and S. grown in the greenhouse were used for P. xylostella and S. litura. Ten larvae were placed on a leaf disk, and their fates were monitored after 2 days for P. xylostella and S. litura. Bioassays were repeated at least twice.
RESULTS
Amplicon restriction fragment length polymorphism was done in order to identify the presence of cry gene variants among the native isolates. The annealing temperature of cry1I full length primer was standardized by using reference strain B. thuringiensis subsp. kurstaki (HD1). CFE20(3) and CFE25(2) were selected for amplicons restriction length polymorphism analysis in order to find out the variants. When the PCR amplicon of CFE20(3) and HD1 were restricted with KpnI and XmaI, there were differences observed in restriction fragments in relation to the reference strain HD1 (Figure 1).
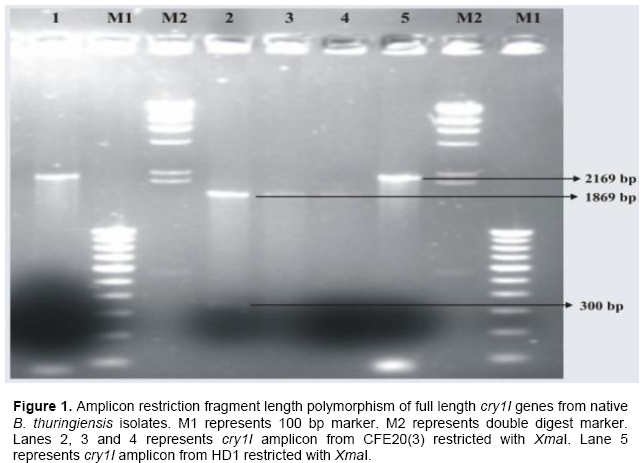
There was one restriction site of XmaI in CFE20(3) giving rise to 1869 and 300 bp as compared to the reference strain HD1 which give an unrestricted band of 2169 bp. CFE20(3) showing varied restriction pattern were selected for further cloning and expression studies.
Based on the ARFLP pattern of the cry1I, variant from the native B. thuringiensis isolates viz., CFE20(3) was cloned into T/A cloning vector pTZ57R/T. The large scale amplification of full length gene encoding cry1I(2169 bp) from CFE20(3) was obtained using cry1I specific primers. A sharp amplicon of the expected size (2169 bp) was eluted from the preparative gel, which was then cloned into a cloning vector, pTZ57R/T. These constructs was transformed into E. coli DH5α and transformants were confirmed by PCR and restriction analysis using BamHI and PstI endonucleases separately giving rise to linear fragment of 4.9 kb including both vector and insert.
The construct containing cry1I was sequenced through primer walking employing M13 primers. The available sequence information from cloned fragments was analyzed using BLAST algorithm available at http:/www.ncbi.nlm.nih.gov. Multiple alignment of amino acid sequences showed 99 percent homology to that of the published cry1IAa sequence.
Following restriction, the 2169bp cry1I fragment from the plasmid DNA were individually cloned into BamHI and PstI sites of the expression vector pQE30 and transformed into E. coli JM109 cells which is a maintenance host. The transformants with cry1I were picked and streaked on Luria agar plates containing Amp100 and Nal10 selection pressure. The recombinants obtained from clones were confirmed by digestion with BamHI and PstI containing both insert (2169bp) and vector (3400 bp) following electrophoresis (Figure 2).
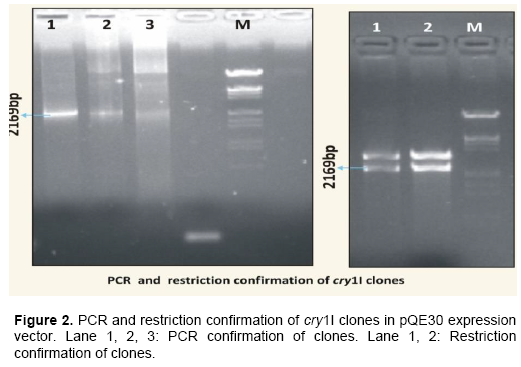
Later, the plasmid from the confirmed recombinant clones in E. coli JM109 cells were transformed into two expression hosts viz. E. coli M15(pREP4) and E. coli SG13009(pREP4). The recombinants were again confir-med by digestion with BamHI and PstI containing both insert (2169 bp) and vector (3400 bp) fragments of the plasmid pREP4 which is present in the host strains E. coli M15 (pREP4) and E. coli SG13009(pREP4) following electrophoresis.
To check the expression of cloned cry1I gene in pQE30, the total protein from IPTG induced cry1I clones in E. coli M15 (pREP4) and E. coli SG13009 (pREP4), plain pQE30 (negative control) and the plain host strain viz. E. coli M15 (pREP4) and E. coli SG13009(pREP4) (negative control) were subjected to SDS PAGE. The protein band of approximately 81kDa in clones and absence of such bands in negative control when loaded at equal amount of protein (500 µg) were observed (Figure 3).
For further confirmation of cry1I clones which expressed 81kDa proteins, bioassay was done against third instar larvae of P. xylostella and S. litura by feeding the total protein of 500 mg/ml. Plain vector pQE30 in both E. coli M15 (pREP4) and the host strains viz. E. coli M15(pREP4) without insert and leaf discs dipped in distilled water served as negative controls. The percent mortality at 24, 48 and 72 h interval was recorded. At 72 h interval, the percent mortality of clones ranged from 60 to 90%. The clone containing cry1I recorded 86.67% mortality (Figure 3). In negative controls, which included the plain E. coli, E. coli with vector and plain water, no mortality was recorded.
When compared with an E. coli extract containing the empty vector, SDS-PAGE analysis demonstrated the presence of a band with a molecular weight of approximately 81 kDa, corresponding to the Cry1Ia protein and several other E. coli proteins (Figure 3).
In the P. xylostella bioassay, protein lysate of cry1Ia construct under induced conditions shows the mortality of 90% under uninduced conditions 40%, where as protein lysate of PQE30 plain vector shows mortality of 30% (Abbott’s formulae, 1925) (Figure 4). Protein lysate of cry1Ia construct was found to be ineffective against S. litura.

DISCUSSION
The use of E. coli as a power house to express genes of various microorganisms has been broadly used successfully. With this technique, a cry1Ia gene from a local isolate of B. thuringiensis CFE20(3) was cloned, expressed in E. coli and the toxicity of the recombinant protein subsequently tested against third instar larvae of Diamond back moth, P. xylostella and In a similar study conducted by Bergamasco et al. (2011), the expression of Cry1Ia protein was confirmed with molecular weight of approximately 81KDa. In a study conducted by Grossi-de-Sa et al. (2007), a new cry1I gene, designed as cry1Ia7, was isolated and protein expressed in E. coli. This protein was demonstrated to be soluble in water and toxic to various insect species. Song et al. (2003) reported that Cry1I protein was shown to be toxic to the diamond back moth (Plutella xylostella), Asian corn borer (Ostrinia furnacalis) and soyabean pod borer (Leguminivora glycinivorella). It was not toxic to the cotton boll worm (Helicoverpa armigera), beet armyworm (Spodoptera exigua), or elm leaf beetle (Pyrrhalto aenescens) in bioassays. Similarly, in the present study, it was observed that expressed product was toxic to the P. xylostella and showed 86.67% mortality but it was not effective against S. litura. Cry1Aa, Cry1Ab, Cry1C and Cry1Da were found to cause more than 93% mortality against third instar larvae of P. xylostella (Cheng et al., 2005). Bergamasco et al. (2011) observed that Cry1I was found to be toxic against Spodoptera frugiperda and Anthonomous grandis. Although many toxins have been found in B. thuringiensis strains, only a few of them have been used to effectively control some determined insect pests. Moreover, some insect pests have developed resistance against some B. thuringiensis toxins. In order to solve these problems, isolation of new strains and toxins is crucial. This study provided a identification of cry1I type gene with new insecticidal properties. However, its insecticidal characteristics may be explored if the Cry1I proteins were successfully expressed in transgenic plants. The present work demonstrated the efficiency of the bacterial system for the expression of the B. thuringiensis Cry1Ia protein, and the high toxicity of the protein to P xylostella.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
We wish to thank Department of Science and Technology, Government of India for providing Inspire Fellowship. This work was supported by a grant from Department of Biotechnology, Government of India, New Delhi.
REFERENCES
|
Abbott WS (1925). A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 18:265-267. |
|
|
Bergamasco VB, Goncalves JF, Polanczk RA, Desiderio JA, Lemos MV (2011) . Expression of a new Bacillus thuringiensis cry1Ia gene in Escherichia coli with strong activity against cotton pests. Aust. J. Basic Appl. Sci. 5(12): 526-533. |
|
|
Birmboim HC, Doly J (1979). A rapid alkaline lysis procedure for screening recombinant plasmid DNA. Nucleic Acids Res. 7(6):1515-1523. |
|
|
Cheng JC, Hsieh FC, Liu BL, Kao SS (2005) . Cloning and expression of Cry1Ab, Cry1C and Cry1Da genes from Bacillus thuringiensis var. aizawai., 6th Pacific conference on the Biotechnology of Bacillus thuringiensis and its Environment impact, Victoria BC. |
|
|
Choi SK, Shin BS, Kong EM, Rho HM, Park SH (2000). Cloning of a new Bacillus thuringiensis Cry1I-type crystal protein. Curr. Microbiol. 41: 6569. |
|
|
Crickmore N, Zeigler DR, Schnepf E, Van Rie J, Lereclus D, Baum J,Bravo A, Dean DH (1998). Bacillus thuringiensis toxin nomenclature[online]http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/index.html |
|
|
Gleave AP, Williams R, Hedges RJ (1993). Screening by polymerase chain reaction of Bacillus thuringiensis serotypes for the presence of CryV-like insecticidal protein genes and characterization of a cryV gene cloned from Bacillus thuringiensis subsp kurstaki. Appl. Environ. Microbiol. 59:1683- 1687. |
|
|
Grossi-de-Sa MF, Magalhaes MQ, Silva MS, Silva SMB, Dias SC, Nakasu YET, Brunetta PSF, Oliveira GR, Oliveira OB, Neto RS, Oliveira LHB, Soares MAZ, Ayub HA., Siqueira A, Figueira ELZ (2007) . Susceptibility of Anthonomus grandis (Cotton Boll Weevil) and Spodoptera frugiperda (Fall Armyworm) to a Cry1Ia-type Toxin from a Brazilian Bacillus thuringiensis Strain. J. Biochem. Mol. Biol. 40(5):773-782. |
|
|
Kuo WS, Chak KF (1996). Identification of novel cry-type genes from Bacillus thuringiensis strains on the basis of restriction fragment length polymorphism of the PCR-amplified DNA. Appl. Environ. Microbiol. 62(4):1369-1377. |
|
|
Lamptey J, Hendrick CV, Tomes NJ, Brown S, Dean DH (1991).Bacillus. In: Levin MA, Seider RJ, Rogul M (eds) Microbial Ecology :Principles, Methods and Applications, McGraw-Hill, New York. pp.20-22. |
|
|
Lorence A, Darszon A, Quintero R, Diaz C, Lievano A, Bravo A (1995) . Delta- endotoxins induce cation channels in Spodoptera frugiperda brush border membranes in suspension and in planar lipid bilayers. FEBS Lett. 360:217-222. |
|
|
Pietrantonio PV, Gill SS (1996). Bacillus thuringiensis delta -endotoxins : action on the insect midgut (eds. M.J. Lehane and P.F. Billingsley) . Chapman and Hall, London. pp. 345-372 . |
|
|
Pooja AS, Krishnaraj PU, Prashanthi SK (2013). Profile of cry from native Bacillus thuringiensis isolates and expression of Cry1I. Afr. J. Biotechnol.12(22):3545-3562. |
|
|
Santos RC, Marcellino LH, Monnerat RG, Gander EG (2003). Mechanical damage in cotton buds caused by the boll weevil. Pesquisa Agropecuária Brasileira. 38:1351-1355 |
|
|
Sambrook J, Russell DW (2001) . Molecular Cloning: A laboratory Manual. Cold Spring Harbor Laboratory Press. |
|
|
Schwartz JL, Masson L, Brousseau R, Rousseaeu E (1993). Lepidopteran specific crystal toxins from., Garneau Bacillus thuringiensis form cation and anion selective channels in planar lipid bilayers . J. Membr. Biol. 132: 53-62. |
|
|
Selvapandiyan A, Reddy VS, Kumar PA, Tewari KK, Bhatnagar RK (1998) . Transformation of Nicotiana tabacum with a native cry1Ia5 gene confers complete protection against Heliothis armigera. Mol. Breed. 4:473-478. |
|
|
Song F, Zhang J, Gu A, Wu Y,Han L, He K, Chen Z, Yao J, Hu Y, Li G, Huang D (2003). Identification of cry1I type genes from Bacillus thuringiensis strains and characterization of a novel cry1I type gene. Appl. Environ. Microbiol. 69(9):5207-5211. |
|
|
Tounsi S, Zouari N, Jaoua S (2003). Cloning and study of the expression of a novel cry1Ia-type gene from Bacillus thuringiensis subsp kurstaki. J. Appl. Microbiol. 95:23-28. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


