Full Length Research Paper

ABSTRACT
Enterobactericeae producing extended spectrum b-lactamases (ESBL) are likely to express carbapenemases OXA-48 which have hydrolytic activity on carbapenems. The aim of this work was to evaluate prevalence of blaOXA-48 for Klebsiella pneumoniae isolates producing ESBL. This work was conducted at the French Reference Center for Antibiotic Resistance with strains from Senegalese Hospital. Standard antibiogram was performed in accordance to CA-SFM/EUCAST 2016 and presence of ESBL was confirmed by synergistic image. Polymerase chain reaction (PCR) was performed to detect systematically blaOXA-1, blaTEM-1, blaCTX-M-1, blaCTX-M-9 and blaOXA-48 gene in case of decrease sensitivity to carbapenem. PCR products were extracted, purified, sequenced and whole-genome sequence (WGS) were used for the analyses. Plasmids extraction was performed by Kado and Liu method. Five isolates harbored a decreased susceptibility to carbapenems. They were positive for blaOXA-48 gene and also expressed blaCTX-M-15. Analysis of the five plasmids by WGS identified a single IncL/M type plasmid of 63 kb and other genes for aminoglycosides and quinolones resistance. Carbapenemase-producing Enterobacteriaceae represent new threat to public health. Decrease in carbapenem susceptibility should be an alert for rapid detection of carbapenemases and to prevent their spread. Phenotypic or molecular methods should be available in many laboratories to take appropriate preventive and therapeutic measures.
Key words: Klebsiella pneumoniae, extended spectrum b-lactamases (ESBL), carbapenemase, blaOXA-48, Senegal.
INTRODUCTION
Carbapenems, antibiotics of β-lactam family, are active on most Gram negative bacilli, including Enterobacteriaceae and non-fermenting bacteria such as Pseudomonas aeruginosa and Acinetobacter baumannii (Nordmann et al., 2010). Exclusively used in hospitals, they are reserved for treatment of multi-resistant bacterial infections. However, these last two decades was marked by emergence of bacteria producing carbapenemases able to hydrolyze them. Carbapenemases were increasingly described to constitute threat and universal public health problem. These enzymes have been described in many African countries; however, their prevalence is poorly defined and has not been systematically studied (Manenzhe et al., 2015). Carbapenemases belong to three classes according to Ambler classification (Walther-Rasmussen et al., 2006). Class A and D are referred to as serine because they have serine at their active sites, whereas class B have zinc (zinc dependent) at their active site and are referred to as metallo-β-lactamases. Class A enzymes group together Klebsiella pneumoniae carbapenemase (KPC), imipenem (IMI) and Guyana extended-spectrum (GES) and hydrolyze all β-lactams. Class B metallo-β-lactamases are plasmid mediated, or in some cases chromosomal, and the most common enzymes among clinical isolates in this group include Verona imipenemase (VIM), imipenemase (IMP) and New Dehli Metallo-β-lactamase (NDM). These enzymes are able to hydrolyse all β-lactams except for aztreonam, monobactam and their hydrolytic activity is reduced or inhibited in vitro by ethylene diamine tetra-acetic acid (EDTA) but not by clavulanic acid. Finally, class D or oxacillinase (OXA-48, OXA-163 and OXA-181), attacks poorly or non third generation cephalosporins and are not inhibited by clavulanic acid or EDTA (Queenan et al., 2007). OXA-48 enzyme was first described in Turkey in 2004 and then rapidly disseminated in several European countries (Carrër et al., 2010). This later was also described in North Africa: Tunisia (Cuzon et al., 2010), Algeria (Agabou et al., 2014), Morocco (Benouda et al., 2010); in West Africa: Senegal (Moquet et al., 2011), Côte d'Ivoire (Jeannot et al., 2013); and in East Africa: Kenya (Pitout et al., 2008). In Senegal, the first description of OXA-48 was made in 2011, but probably dates back further since imipenem introduction in 2008. BlaOXA-48 gene is often associated with presence of ESBL, thus conferred multi-resistance of bacteria. Simultaneous presence of several mechanisms of resistance in a same bacterium associated with absence of new classes of antibiotics makes complex therapeutic monitoring of patients. Phenotypic or molecular detection of carbapenemases thus becomes indispensable to reduce their spread. Several methods are available, such as phenotype techniques (modified Hodge test), hydrolytic method (carbaNP), mass spectrometry or molecular methods (Burckhardt et al., 2011; Hrabak et al., 2011; Poirel et al., 2015). This research is not easy in countries with limited resource and can better explain underestimated prevalence.
MATERIALS AND METHODS
Bacterial isolates were collected from two senegaleses hospitals and included forty-nine Klebsiella pneumoniae from urinary samples. These isolates were transported according to recommendations of biological products for characterization in the National Reference Center (CNR) for resistance of enterobacteria in Clermont-Ferrand, France. After reculture on Trypticase-Soja-Agar (TSA), bacterial identification was confirmed by mass spectrometry VITEK® MS (Biomérieux, La Balme, France). Antibiogram was performed by diffusion method according to the recommendations of CA-SFM-EUCAST 2016 (CASFM, 2016). Presence of ESBL was confirmed by the synergistic image between combination with third generation of cephalosporins (C3G) and amoxicillin-clavulanic acid. The DNA of each isolate was extracted from colonies by thermal shock and then used as a template to search the following genes: blaCTX-M1 (CTT CCA GAA TAA GGA ATC; CCG TTT CCG CTA TTA CAA), blaCTX-M9 (CTG ATG TAA CAC GGA TTG AC; TTA CAG CCC TTC GGC GAT), blaOXA-1 (ATATTATCTACAGCAGCG; TTGGCTTTTATGCTTGATG) blaTEM-1 (TAA AAT TCT TGA AGA CG; TCT GAC AGT TAC CAA TGC). For five cases with decrease susceptibility to carbapenems (ertapenem, imipenem), PCR of blaOXA-48 (GGG GAC GTT ATG CGT GTA TT; OXA-48B (GAG CAC TTC TTT TGT GAT GGC) and others genes (repA, traU, parA) were performed as previously described (Beyrouthy et al., 2014). PCR products were extracted, purified (UltraClean® Microbial DNA Isolation kit, Quiagen) and sequenced by Sanger method. Transferability of the blaOXA-48 gene was studied by a mating-out assay using Escherichia coli rifampicin resistant as recipient bacteria. Selection was performed on agar plates supplemented with ticarcillin (32 mg/L) and rifampicin (300 mg/L). The plasmid content of the bacteria and the size of plasmids were determined using plasmid DNA extracted by Kado and Liu method. Whole genome sequencing (WGS) and genome assembly: Bacterial DNA was extracted from overnight cultures with DNeasyUltraClean Microbial Kit (Qiagen, Hilden, Germany) as recommended by the manufacturer. The whole-genome sequences of isolates were determined by de novo assembly of 2x150-bp paired-end reads generated using Illumina sequencing technology (San Diego, CA, USA). The assemblies were performed using SPAdes (Nurk et al., 2013), the reads were mapped with the Burrows-Wheeler aligner (BWA) (Li et al., 2009), the final polishing of the assembly used Pilon (Walker et al., 2014) and the genome was annotated with RAST (Aziz et al., 2008).
Whole-genome sequence analysis
Isolates were typed in silico using multi-locus sequence typing (MLST) with the K. pneumoniae scheme (http://bigsdb.pasteur.fr/klebsiella/klebsiella.html). The antibiotic resistance genes were characterized using the Comprehensive Antibiotic Resistance Database (CARD) and Resfinder (Jia et al., 2017; Zankari et al., 2012). Replicon typing was performed with PlasmidFinder (http://www.genomicepidemiology.org/) using the Enterobacteriaceae database with the detection thresholds set to 95% sequence identity.
RESULTS
Forty-nine isolates of K. pneumonia producing ESBL were tested to research the presence of blaCTX-M-1, blaCTX-M-9, blaTEM-1 and blaOXA-1 genes. Forty-six of the isolates (93%) carried a blaCTX-M1 type gene. The blaTEM-1 and blaOXA-1 genes were present in n = 40/49 and n = 38/49 isolates respectively. Five isolates showed a decreased sensitivity to imipenem and ertapenem (Figure 1). Polymerase chain reaction (PCR) multiplex of carbapenemase was positive for the blaoxa-48 gene in all five isolates, and this was confirmed by the simplex PCR blaoxa-48. The successful plasmid extraction showed two types, one with a PM between 62 and 70 Kb probably associated with the blaOXA-48 gene as shown in the Figure 2. Transconjugants were obtained from all the clinical OXA-48-encoding strains by a mating-out assay. All of them expressed a β-lactam resistance phenotype compatible with OXA-48 production, and the presence of the blaOXA-48 gene was confirmed by PCR. Plasmids from clinical isolates and their transconjugants were extracted and hybridized with a probe specific for the blaOXA-48 gene. The results showed that the blaOXA-48 gene was located in all isolates on ~ 63 kb conjugative plasmids. Further analysis of these plasmids from the transconjugants revealed similar restriction profiles (data not shown). These results suggested that the plasmids in isolates from Senegal shared the IncL/M pOXA-48a-like backbone. A similar 63,434 bp-long IncL/M plasmid, carried blaOXA-48 was identified by WGS in all isolates. The blaOXA-48 encoding plasmid from Senegal covered 99.98% of the reference plasmid pOXA-48a (JN626286.1). Difference between Senegalese plasmids and pOXA-48a were the presence of the blaOXA-48 gene embedded within a Tn1999.2-type composite transposon, which differs from Tn1999 by the insertion of the IS1R insertion sequence in the IS1999 insertion sequence located upstream of blaOXA-48, and the truncation of the hypothetical gene orf25 with an additional copy of the IS1R. Other genes were detected as repA, traU and parA involved in the replication, transport and partition of plasmid pOXA-48a, which further confirms its presence (Figure 3). All of these five isolates expressed simultaneously the blaCTX-M-15 gene but also other β-lactamases and markers of resistance to aminoglycosides and quinolones (Table 1).
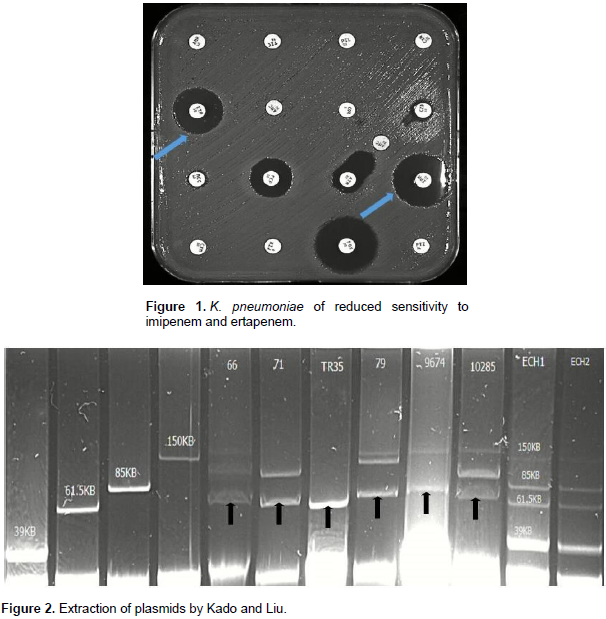
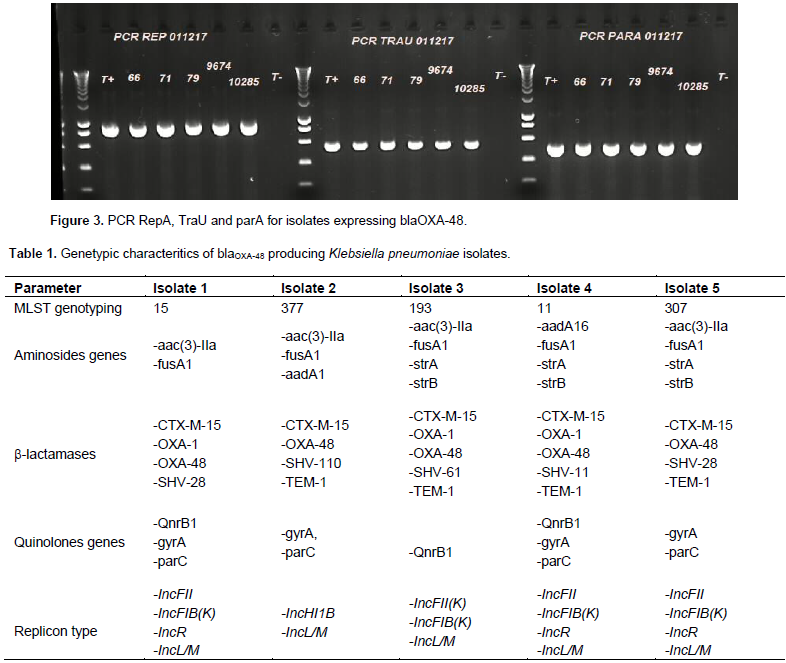
DISCUSSION
A prevalence of 10% for the blaOXA-48 gene was found in K. pneumoniae isolates. All of them hydrolyzed third generation of cephalosporins and aztreonam, and simultaneously expressed blaCTX-M-15. The carbapenemase OXA-48 confer resistance to β-lactamase inhibitors and are known to hydrolyze carbapenems and third generation cephalosporins more weakly (Bonnin et al., 2013; Tzouvelekis et al., 2014). However, in some Enterobacteriaceae (K. Pneumoniae and E. coli), the association of OXA-48 with ESBL (CTX-M or SHV) or lack of membrane permeability increase level resistance to carbapenems (Bonnet et al., 2011; Cuzon et al., 2011). OXA-48 is the main carbapenemase produced by Enterobacteriaceae and most widespread enzyme that has emerged in all countries around Mediterranean and in Africa (Carrër et al., 2008; Nordmann, 2014). It is mainly found in K. pneumoniae and E. coli, although other species of Enterobacteriaceae can produce this enzyme. OXA-48 has distinction of spreading more easily between species of this family compared to other enzymes such as KPC, NDM (Nordmann et al., 2014). In France, among Enterobacteriaceae producing carbapenemases, K. pneumoniae remained majority with 59% and OXA-48 enzyme was observed up to 52% (Poirel et al., 2004). Several risk factors are associated with the acquisition of carbapenemase producing bacteria in healthcare settings, such as recent antibiotic therapy, prolonged hospital stay, use of invasive devices and immunosuppression (Jeon et al., 2008). This study was unable to establish a link between infection with these multidrug resistant bacteria and level of mortality. However, in literature it has been shown that mortality is high with infections caused by bacteria expressing blaOXA-48 gene (Doi et al., 2015). All these five isolates also expressed ESBL CTX-M-15; this combination OXA-48/CTX-M-15 leads to a higher level of carbapenem resistance. This association has often been found in other studies (Carrër et al., 2010; Cuzon et al., 2011). In Tunisia, isolates of K. pneumoniae that simultaneously expressed carbapenemaseVIM-4, CTX-M-15 and CMY-4 were observed (Ktari et al., 2006). The blaTEM-1 and blaOXA-1 genes were expressed in four and three strains, respectively.
The same association of blaOXA-48, blaTEM and blaOXA-1 has been already described in Senegal by Moquet et al. (2011) with nosocomial strains of K. pneumoniae. In the same study, a community strain of K. pneumoniae expressed only blaOXA-48. All these OXA-48 isolates expressed other determinants of aminoglycoside and / or quinolone resistance. Coexistence between CTX-M and aminoglycoside and / or quinolone resistance genes has been described in Africa (Alibi et al., 2015; Breurec et al., 2013). In Senegal, Moquet et al. (2011) observed a co-presence of CTX-M15 / AAC'6 at 78.8% and CTX-M15 / QNR at 65%. These different mechanisms of resistance in the same bacteria have a direct consequence on the management which becomes more complex with a reduction of the therapeutic arsenal. The blaOXA-48 gene is usually carried by a 62 kb plasmid; analysis of that plasmids by WGS sequencing shows a single plasmid belonged to the incompatibility group IncL of 63 kb. IncL type plasmids are currently detected worldwide in Enterobacteriaceae isolates of different origin and are considered to be epidemic resistance plasmids and contribute to the diffusion of the carbapenemase-encoding genes blaNDM-1 and blaOXA-48 (Beyrouthy et al., 2014; Bonnin et al., 2013). The presence of the repA, traU and parA genes further confirmed the plasmid support of the blaOXA-48 gene found in these isolates. Therapeutic management of carbapenem-producing enterobacteria makes use of other antibiotics such as aminoglycosides, tigecycline, colistin, fosfomycin or quinolones (Nordmann et al., 2011). Although efficacy of colistin and tigecycline are not sure, activity can be increasing when they are combined (Tzouvelekis et al., 2014). Mortality rate from infections with Enterobacteriaceae producing carbapenemase shows that current therapeutic approaches are insufficient and need to be revised. Nevertheless, mortality may be reduced if patients are treated with a combination containing carbapenem (Tzouvelekis et al., 2014). Mechanisms of resistance are usually combined in the same strain and often only colistin or tigecycline sometimes remain effective (Chassagne, 2012). However, combination of these two antibiotics can reduce mortality (Carmeli et al., 2010). Detection of carbapenemase then remains a necessity in case of decreased carbapenem sensitivity in enterobacteria producing ESBL. However, this task seems difficult in our countries where the majority of laboratories have a limited number of phenotypic methods or molecular techniques.
CONCLUSION
Carbapenemase OXA-48 expressed by enterobacteria producing ESBL represents a new threat to public health. Surveillance of antimicrobial resistance remains a necessity in both hospital and community. This one must integrate the phenotypic detection and the molecular characterization of the genes involved. This will reduce the spread of the bacteria that harbor them and take appropriate preventive and therapeutic measures.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors are grateful to CNR of Enterobacteriaceae in Clermont Ferrand (France) and the French cooperation which support the experiment of this study. Their sincere thanks go to the practitioners in the Laboratory of Bacteriology of the Saint Louis Hospital, Aristide Le Dantec hospital and CNR of Enterobacteriaceae for their technical assistance.
REFERENCES
|
Agabou A, Pantel A, Ouchenane Z, Lezzar N, Khemissi S, Satta D, Sotto A, Lavigne JP (2014). First description of OXA-48-producing Escherichia coli and the pandemic clone ST131 from patients hospitalised at a military hospital in Algeria. European journal of clinical microbiology and infectious diseases. 33(9):1641-1646. |
|
|
Alibi S, Ferjani A, Boukadida J (2015). Molecular characterization of extended spectrum beta-lactamases produced by Klebsiella pneumoniae clinical strains from a Tunisian Hospital. Medicine and Infectious Diseases, 45(4):139-143. |
|
|
Aziz RK, Bartels D, Best AA, DeJongh M, Disz T, Edwards RA, Formsma K, Gerdes S, Glass EM, Kubal M, Meyer F (2008). The RAST Server: rapid annotations using subsystems technology. BMC Genomics. 9:75. |
|
|
Benouda A, Touzani O, Khairallah MT, Araj GF, Matar GM (2010). First detection of oxacillinase-mediated resistance to carbapenems in Klebsiella pneumoniae from Morocco. Annals of tropical medicine and parasitology, 104(4):327-330. |
|
|
Beyrouthy R, Robin F, Delmas J, Gibold L, Dalmasso G, Dabboussi F, Hamzé M, Bonnet R (2014). IS1R-mediated plasticity of IncL/M plasmids leads to the insertion of bla OXA-48 into the Escherichia coli Chromosome. Antimicrobial Agents and Chemotherapy, 58(7):3785-3790. |
|
|
Bonnet R (2011). β-lactamines et entérobactéries. In : Antibiogramme ; 3ème Edition, Chapitre 16:165-188. View. |
|
|
Bonnin RA, Nordmann P, Poirel L (2013). Screening and deciphering antibiotic resistance in Acinetobacteria baumannii: a state of the art. review of anti-infective therapy, 11(6):571-583. |
|
|
Breurec S, Guessennd N, Timinouni M, Le TA, Cao V, Ngandjio A, Randrianirina F, Thiberge JM, Kinana A, Dufougeray A, Perrier-Gros-Claude JD, Boisier P (2013). Klebsiella pneumoniae resistant to third-generation cephalosporins in ï¬ve African and two Vietnamese major towns: multiclonal population structure with two major international clonal groups, CG15 and CG258. Clinical Microbiology Infection, 19(4):349-355. |
|
|
Burckhardt I, Zimmermann S (2011). Using matrix-assisted laser desorption ionization-time of flight mass spectrometry to detect carbapenem resistance within 1 to 2.5 hours. Journal Clinical Microbiology, 49(9):3321-3324. |
|
|
Carmeli Y, Akova M, Cornaglia G, Daikos GL, Garau J, Harbarth S, Rossolini GM, Souli M, Giamarellou H (2010). Controlling the spread of carbapenemase-producing Gram-negatives: therapeutic approach and infection control. Clinical Microbiology Infection, 16(2):102-111. |
|
|
Carrër A, Poirel L, Eraksoy H, Cagatay AA, Badur S, Nordmann P (2008). Spread of OXA-48-Positive Carbapenem-Resistant Klebsiella pneumoniae isolates in Istanbul, Turkey. Antimicrobial Agents and Chemotherapy, 52(8):2950-2954. |
|
|
Carrër A, Poirel L, Yilmaz M, Akan OA, Feriha C, Cuzon G et al (2010). Emerging spread of OXA-48-encoding plasmid from Turkey and beyond. Antimicrobial Agents and Chemotherapy, 54(3):1369-1373. |
|
|
Chassagne C (2012). Caractérisation phénotypique de souches d'entérobactéries produisant une oxacillinase-48 isolées lors d'une épidémie survenue au CHU de Nancy en 2009-2011. Thèse Pharmacie, Université de Lorraine. View. |
|
|
Comité de l'Antibiogramme de la Société Française de Microbiologie (CASFM) (2016). Recommandations. View. |
|
|
Cuzon G, Naas T, Lesenne A, Benhamou M, Nordmann P (2010). Plasmid-mediated carbapenem-hydrolysing OXA-48 b-lactamase in Klebsiella pneumoniae from Tunisia. International Journal of antimicrobial Agents, 36(1):91-93. |
|
|
Cuzon G, Ouanich J, Gondret R, Naas T, Nordmann P (2011). Outbreak of OXA-48-positive carbapenem-resistant Klebsiella pneumoniae isolates in France. Antimicrobial Agents and Chemotherapy, 55(5):2420-2423. |
|
|
Doi Y, Paterson DL (2015). Carbapenemase-Producing Enterobacteriaceae. Seminars in Respiratory and Critical Care Medicine, 36(1):74-84. |
|
|
Hrabak J, Walkova R, Studentova V, Chudackova E, Bergerova T (2011). Carbapenemase activity detection by matrixassisted laser desorption ionization-time of flight mass spectrometry. Journal of clinical microbiology, 49(9):3222-3227. |
|
|
Jeannot K, Guessennd N, Fournier D, Meuller E, Gbonon V, Plesiat P (2013). Outbreak of metallo-b-lactamase VIM-2-positive strains of Pseudomonas aeruginosa in the Ivory Coast. Journal of Antimicrobial Chemotherapy, 68(12):2952-2954. |
|
|
Jeon MH, Choi SH, Kwak YG Chung JW, Lee SO, Jeong JY, Woo JH, Kim YS (2008). Risk factors for the acquisition of carbapenem-resistant Escherichia coli among hospitalized patients. Diagnostic Microbiology and Infectious Disease, 62(4):402-406. |
|
|
Jia B, Raphenya AR, Alcock B, Waglechner N, Guo P, Tsang KK et al. (2017). CARD 2017: expansion and model-centric curation of the comprehensive antibiotic resistance database. Nucleic Acids Res.45(D1): D566-D573. doi: 10.1093/nar/gkw1004. |
|
|
Ktari S, Arlet G, Mnif B, Gautier V, Mahjoubi F, Ben Jmeaa M, Bouaziz M, Hammami A (2006). Emergence of multidrug-resistant Klebsiella pneumoniae isolates producing VIM-4 metallo-β-lactamase, CTX-M-15 extended-spectrum β-lactamase, and CMY-4 AmpCβ-lactamase in a Tunisian university hospital. Antimicrobial Agents and Chemotherapy,50(12):4198-4201. |
|
|
Li H, Durbin R (2009). Fast and accurate short read alignment with Burrows-Wheeler transform. Bio informal Oxford England, 15;25(14):1754-1760. |
|
|
Manenzhe RI, Zar HJ, Nicol MP, Kaba M (2015). The spread of carbapenemase-producing bacteria in Africa: a systematic review. Journal of antimicrobial chemotherapy, 70(1):23-40. |
|
|
Moquet O, Bouchiat C, Kinana A, Seck A, Arouna O et al. (2011). Class D OXA-48 carbapenemase in multidrug-resistant enterobacteria, Senegal. Journal of emerging infectious diseases, (Centers for Disease Control and Prevention). Letters. 17 (1):143-144. |
|
|
Nordmann P, Carrer A (2010). Carbapenemases in Enterobacteriaceae. Archives of pediatric, 17:S154-62. |
|
|
Nordmann P, Poirel L, Toleman MA, Walsh TR (2011). Does broad- spectrum β-lactam resistance due to NDM-1 herald the end of the antibiotic era for treatment of infections caused by Gram-negative bacteria? Journal of antimicrobial chemotherapy, 66(4):689-692. |
|
|
Nordmann P (2014). Carbapenemase producing Enterobacteriaceae overview of a major public health challenge. Medicine and Malaria Infection, 44(2):51- 56. |
|
|
Nordmann P, Poirel L (2014). The difficult to control spread of carbapenemase producers among Enterobacteriaceae worldwide. Clinical Microbiology and Infectious Diseases, 20(9):821-830. |
|
|
Nurk S, Bankevich A, Antipov D, Gurevich AA, Korobeynikov A, Lapidus A, Prjibelski AD, Pyshkin A, Sirotkin A, Sirotkin Y, Stepanauskas R (2013). Assembling single-cell genomes and mini-metagenomes from chimeric MDA products. Journal of Computational Biology. 20(10):714-37. |
|
|
Pitout JD, Revathi G, Chow BL, Kabera B, Kariuki S, Nordmann P, Poirel L (2008). Metalloâ€βâ€lactamaseâ€producing Pseudomonas aeruginosa isolated from a large tertiary centre in Kenya. Clinical Microbiology and Infection, 14(8):755-759. |
|
|
Poirel L, Héritier C, Tolün V and Nordmann P (2004). Emergence of Oxacillinase Mediated Resistance to Imipenem in K. pneumoniae. Antimicrobial Agents and Chemotherapy, 48(1):15-22. |
|
|
Poirel L, Nordmann P (2015). Rapidec Carba NP: Test for Rapid Detection of Carbapenemase Producers. Journal of Clinical Microbiology, 53(9):3003-3008. |
|
|
Queenan AM, Bush K (2007). Carbapenemases: the versatile β-lactamases. Clinical Microbiology Reviews, 20(3):440-458. |
|
|
Tzouvelekis LS, Markogiannakis A, Piperaki E, Soul M, Daikos GL (2014). Treating infections caused by carbapenemase-producing Enterobacteriaceae. Clinical Microbiology and Infectious Diseases, 20(9):862-872. |
|
|
Walker BJ, Abeel T, Shea T, Priest M, Abouelliel A, Sakthikumar S et al. (2014). Pilon: an integrated tool for comprehensive microbial variant detection and genome assembly improvement. PloS One. 9(11):e112963. |
|
|
Walther-Rasmussen J, Hoiby N (2006). OXA-type carbapenemases. Journal of antimicrobial chemotherapy, 57(3):373-383. |
|
|
Zankari E, Hasman H, Cosentino S, Vestergaard M, Rasmussen S, Lund O et al. (2012). Identification of acquired antimicrobial resistance genes. Journal of antimicrobial chemotherapy, 67(11):2640-2644. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


