Full Length Research Paper

ABSTRACT
This work highlighted a putative link between the physiological activity and genetic diversity of Methylobacterium species and the association with sugarcane roots and rhizoplane. In total, 40 isolates previously described as pink-pigmented facultative methylotrophic bacteria (PPFMs), were evaluated for their ability to fix nitrogen and solubilize inorganic phosphate, amylase and pectinase activity. This in vitro potential was positively correlated with the community isolated from the root tissues than those from the rhizoplane. Regarding the genomic fingerprinting, the (BOX-PCR) approach revealed a low similarity among the isolates, occurring sole 7 haplotypes harboring more than 70% of similarity among band patterns. These results revealed that the genomic fingerprinting of the isolates recovery from roots is different from the rhizoplane. Besides that, these haplotypes occurred on both sugarcane varieties. Using a phylogenetic sequencing approach based on the 16S rRNA gene, we observed a high abundance of sequences similar to Methylobacterium radiotolerans colonizing both plant tissue and sugarcane varieties were observed. Hence, it was suggested that the plant should select those Methylobacterium spp. with a high biotechnological potential to promote plant growth. Therefore, the bioprospection of specific endophytic bacterial groups comprise an important source of biotechnological potential to improve sugarcane growth and production.
Key words: Pink-pigmented facultative methylotrophic bacteria, plant growth promotion, BOX-PCR, 16S rDNA, Methylobacterium radiotolerans.
INTRODUCTION
MATERIALS AND METHODS
RESULTS AND DISCUSSION
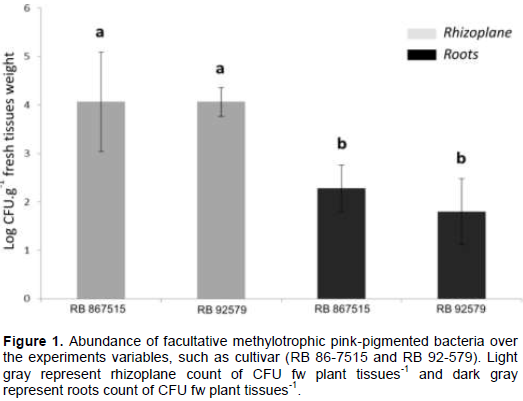
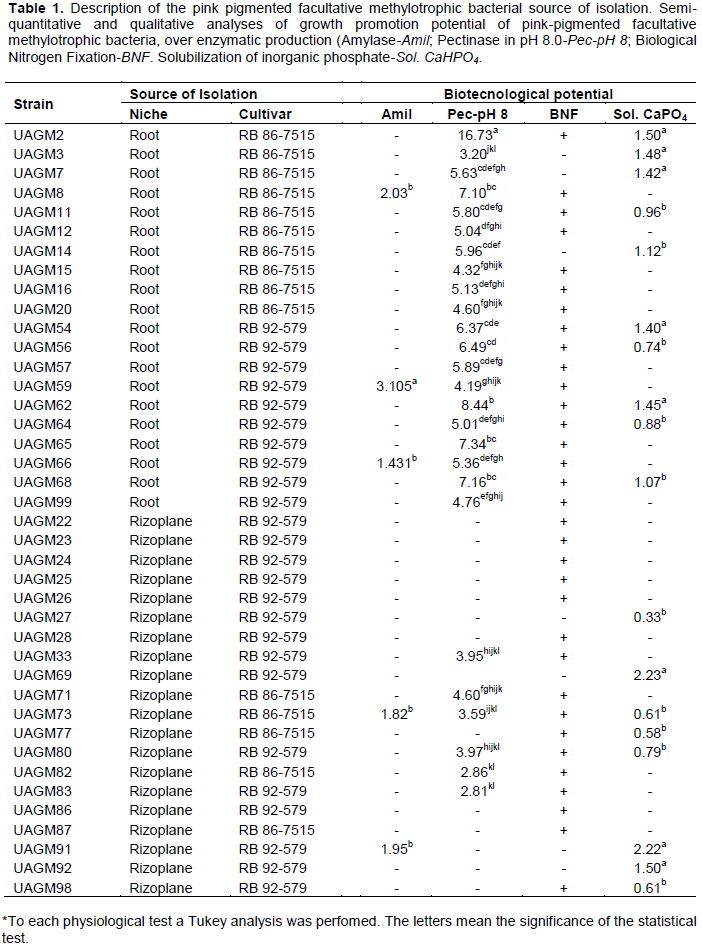
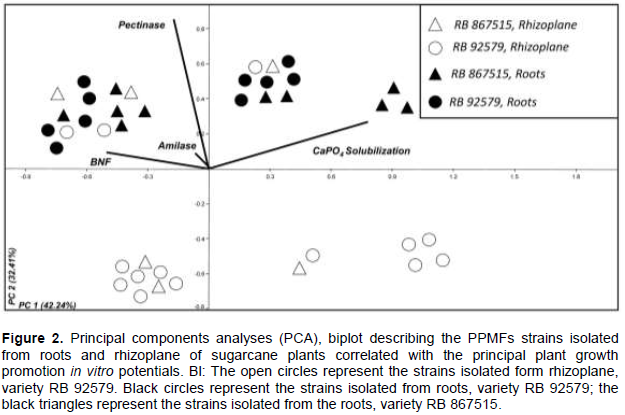
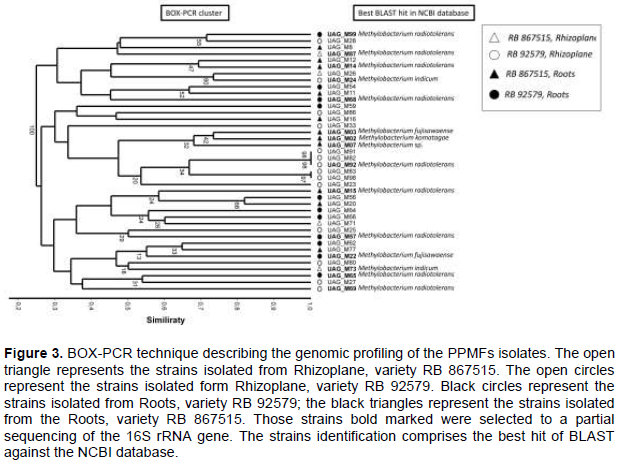

CONCLUSION
The main goal of this study was to show that the association between sugarcane and Methylobacterium spp. are based on many specific traits; the in vitro potentials to promote plant development such as atmospheric nitrogen fixation, phosphate solubilization and extracellular enzyme activity, and specific genomic fingerprints, and not in relation to its taxonomical identity or phylogenetic distance. Hence, those are only some steps in the complete understanding of the interaction between Methylobacterium genus and sugarcane. The complete mechanisms that might determine this relationship are yet to be elucidated. For this reason, further genomic studies must be conducted in order to understand these interaction mechanisms between M. radiotolerans and sugarcane, and even more, their interaction under greenhouse conditions.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors thank CNPQ (180203/2008-8) and (142344/2013-3) for awarding the research grant to Pedro Avelino Maia de Andrade as well as the Rural Federal University of Pernambuco and the Soil Sciences Department at the College of Agriculture “Luiz de Queiroz” University of São Paulo (Esalq-USP). They also acknowledge the critical review of Msc. Priscila Alves Giovani.
REFERENCES
|
Alves MH, Campos-Takaki GM, Porto ALF, Milaniz AI (2002). Screening of Mucor spp. for the production of amylase, lipase, and protease, Brazilian Journal of Microbiology 33:225-230. |
|
|
Ambrosini AS, Passaglia LM (2015). Ecological role of bacterial inoculants and their potential impact on soil microbial diversity. Plant and Soil 400(1-2):193-207. |
|
|
Ardanov P, Lyastchenko S, Karppinen K, Häggman H, Kozyrovska N, Pirttilä AM (2015). Effects of Methylobacterium sp. on emergence, yield, and disease prevalence in three cultivars of potato (Solanum tuberosum L.) were associated with the shift in the endophytic microbial community. Plant and Soil 405(1-2): 299-310. |
|
|
Ardanov P, Sessitsch A, Aggman HH, Kozyrovska N, Pirttila AM (2012). Methylobacterium-induced endophyte community changes correspond with protection of plants against pathogen attack. PLoS ONE 7:e46802. |
|
|
Assumpção LC, Lacava PT, Dias ACF, Azevedo JL, Menten JOM (2009). Diversity and biotechnological potential of endophytic bacterial community of soybean seeds. Brazilian Journal of Agricultural Research 44:503-510. |
|
|
Azevedo JL, Maccheroni Jr W, Pereira JO, Araujo WL (2000). Endophytic microorganism: A review on insect control and recent advances on tropical plants. Electronic Journal of Biotechnology 3(1). doi: 10.2225/vol3-issue1-fulltext-4. |
|
|
Bashan Y, Gonzalez LE (1999). Long-term survival of the plant-growth-promoting bacteria Azospirillum brasilense and Pseudomonas fluorescens in dry alginate inoculant. Applied Microbiology and Biotechnology 51(2):262-266. |
|
|
Batista BD, Lacava PT, Ferrari A, Teixeira-Silva NS, Bonatelli ML, Tsui S, et al (2018). Screening of tropically derived, multi-trait plant growth- promoting rhizobacteria and evaluation of corn and soybean colonization ability. Microbiological Research 206:33-42. |
|
|
Batista BD, Taniguti LM, Almeida JR, Azevedo JL, Quecine MC (2016). Draft Genome Sequence of Multi-trait Plant Growth-Promoting Bacillus sp. Strain RZ2MS9. Genome Announcements 4: e01402-16. |
|
|
Berraquero FR, Baya AM, Cormenzana AR (1976). Index determination for the study of phosphate solubilization by soil bacteria. Ars Pharmaceutica 17:399-406. |
|
|
Bordonal RO, Carvalho JLN, Lal R, de Figueiredo EB, de Oliveira BG, La Scala N (2018). Sustainability of sugarcane production in Brazil. A review. Agronomy for Sustainable Development 38:13. |
|
|
Carrim AJJI, Barbosa EC, Vieira JDG (2006). Enzymatic activity of endophytic bacterial isolates of Jacaranda decurrens Cham. (Carobinha-do-campo). Brazilian Archives of Biology and Technology 49:353-359. |
|
|
Cerqueira VS, Hollenbach EB, Maboni F, Camargo FAO, Peralba MCR, Bento FM (2012). Bioprospection and selection of bacteria isolated from environments contaminated with petrochemical residues for application in bioremediation. World Journal of Microbiology and Biotechnology 28:1203-1222. |
|
|
Ceska M (1971). Enzyme catalysis of solidified media. European Journal of Biochemistry 22:186-192. |
|
|
Chistoserdova SL, Chen W, Lapidus A, Lidstron ME (2003). Methylotrophy in Methylobacterium extorquens AM1 from a genomic point of view. Journal of Bacteriology 185(10):2980-2987. |
|
|
Costa DP, Dias ACF, Durrer A, Andrade PAM, Gumiere T, Andreote FD (2014). Differential composition of bacterial communities in the rhizosphere of sugarcane varieties. Brazilian Journal of Soil Science 38(6):1694-1702. |
|
|
Dourado MN, Andreote FD, Dini-Andreote F, Conti R, Araujo JM, Araujo WL (2012). Analysis of 16s rRNA and mxaF genes revealing insights into Methylobacterium niche-specific plant association. Genetics and Molecular Biology 1:142-148. |
|
|
Dourado MN, Neves AAC, Santos DS, Araujo WL (2015). Biotechnological and agronomic potential of endophytic pink-pigmented methylotrophic Methylobacterium spp. BioMed Research International. 1-19: article ID909016. |
|
|
Galdos, M.V. Cerri, C.C. Cerri, C.E.P (2009). Soil carbon stocks under burned and unburned sugarcane in Brazil. Geoderma 153:347-352. |
|
|
Hammer O, Harper DAT, Ryan PD (2001). PAST: paleontological statistics software package for education and data analysis. Palaeontological Electronica 4:1–9 |
|
|
Hardoim PR, Van Overbreek LS, Van Elsas JD (2008). Properties of bacterial endophytes and their proposed role in plant growth. Trends in Microbiology 16:463–471. |
|
|
Heuer H, Krsek M, Baker P, Smalla K, Wellington EM (1997). Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-eletrophoretic separation in denaturing gradients. Applied and Environmental Microbiology 63:3233–3241. |
|
|
Ibort P, Imai H, Uemura M, Aroca R (2018). Proteomic analysis reveals that tomato interaction with plant growth promoting bacteria is highly determined by ethylene perception. Journal of Plant Physiology 220:43-59. |
|
|
Jayashree S, Vadivukkarasi P, Anand K, Kato Y, Sundaram Seshadri (2011). Evaluation of pink-pigmented facultative methylotrophic bacteria for phosphate solubilization. Archives in Microbiology 193:543-552. |
|
|
Jourand P, Renier A, Rapior S, DE Faria SM, Prin Y, Galiana A, Giraud E, Dreyfus B (2005). Role of methylotrophy during symbiosis between Methylobacterium nodulans and Crotalaria podocarpa. Molecular Plant-Microbe Interaction 18:1061–1068. |
|
|
Kuklinsky-Sobral J, Araujo WL, Mendes R, Geraldi IO. Pizzirani-Kleiner AA, Azevedo JL (2004). Isolation and characterization of soybean-associated bacteria and their potential for plant growth promotion. Environmental Microbiology 6:1244-1251. |
|
|
Leite M, Pereira A, Souza A, Andrade P, Barbosa M, Andreote F, et al (2018). Potentially diazotrophic endophytic bacteria associated to sugarcane are effective in plant growth-promotion. Journal of Experimental Agriculture International 21:1–15. |
|
|
Madhaiyan M, Alex THH, Ngoh S Te, Prithiviraj B, Ji L (2015). Leaf-residing Methylobacterium species fix nitrogen and promote biomass and seed production in Jatropha curcas. Biotechnology and Biofuels BioMed Central 8:1-14. |
|
|
Madhaiyan M, Poonguzhali S, Lee HS, Suandaram SP (2005). Pink-pigmented facultative methylotrophic bacteria accelerate germination, growth and yield of sugarcane clone Co86032 (Saccharum officinarum L.). Biology and Fertility of Soils 41:350-358. |
|
|
Majumder D, Kangjam B, Devi KJ, Lyngdoh D, Tariang J, Thakuria D, Goyal A, Gupta VK, Sharma GD, Tuohy MG, Gaur R (2016). Endophytes: an emerging microbial tool for plant disease management. In: Gupta, V. K., Sharma, G. D., Tuohy, M. G., Gaur, R, ed. The handbook microbiological bioresources. 1st ed. India, Meghalaya: Cabi pp. 179-192. |
|
|
Oliveira JTC, Figueredo EF, Diniz WPS, de Oliveira LFP, de Andrade PAM, Andreote FD, Kuklinsky-Sobral J, de Lima DR, Freire FJ (2017). Diazotrophic bacterial community of degraded pastures. Applied and Environmental Soil Science. ID 2561428, 10 pages. |
|
|
Omer ZS, Tombolin R, Broberg A, Gerhardson B (2004). Indole-3-acetic acid production by pink-pigmented facultative methylotrophic bacteria. Plant Growth and Regulation 43:93-96. |
|
|
Rademaker JLW, de Bruijn FJ (1997). Characterization and classification of microbes by rep-PCR genomic fingerprinting and computer assisted pattern analysis. In DNA Markers: Protocols, Applications and Overviews. Edited by G. Caetano-Anollés P. M. Gresshoff. New York, NY: John Wiley pp. 151-157. |
|
|
Raja P, Uma S, Sundaram S (2006). Non-nodulating pink-pigmented facultative Methylobacterium sp. with a functional nifH gene. World Journal Microbiology and Biotechnology 22:1381-1384. |
|
|
Rodrigues AA, Araújo MVF, Soares RDES, Francesco B, Oliveira RDE, Sibov ST, Vieira JDG (2018). Isolation and screening for multi-trait plant growth promotion actinobacteria from organic sugarcane rhizosphere. International Journal of Microbiology Research 10(5):1193-1198. |
|
|
Rosenblueth M, Martinez-Romero E (2006). Bacterial endophytes and their interactions with hosts. Molecular Plant-Microbe Interactions 19:827-837. |
|
|
Rosenblueth, M, Martinez-Romero E (2004). Rhizobium etli maize populations and their competitiveness for root colonization. Archives in Microbiology 181:337-344. |
|
|
Rosseto PB, Dourado MN, Quecine MC, Andreote FD, Araújo WL, Azevedo JL, Pizzirani-Kleiner (2011). Specific plant induced biofilm formation in Methylobacterium species. Brazilian Journal of Microbiology 42: 878-883. |
|
|
Saitou N, Nei M (1987). The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular Biology and Evolution 4:406-425. |
|
|
Sánchez-López AS, Pintelon I, Stevens V, Imperato V, Timmermans JP, González-Chávez C, Carrillo-González R, Van Hamme J, Vangronsveld J, Thijs S (2018). Seed endophyte microbiome of Crotalaria pumila unpeeled: Identification of plant-beneficial methylobacteria. International Journal of Molecular Sciences 19:1-20. |
|
|
Senthilkumar M, Krishnamoorthy R (2017). Isolation and characterization of tomato leaf phyllosphere methylobacterium and their effect on plant growth. International Journal Current Microbiology and Applied Science 6:2121-2136. |
|
|
Shameer S, Prasad TNVKV (2018). Plant growth promoting rhizobacteria for sustainable agricultural practices with special reference to biotic and abiotic stresses. Plant Growth Regulation 84:603-615. |
|
|
Soedarjo M, Hemscheidt TK, Borthaku D (1994). Mimosine, a toxin present in leguminous trees (Leucaena spp.), induces a mimosine- degrading enzyme activity in some Rhizobium strains. Applied and Environmental Microbiology 60:4268-4272. |
|
|
Sruthilaxmi CB, Babu S (2017). Microbial bio-inoculants in Indian agriculture Ecological perspectives for a more optimized use. Agriculture Ecosystem and Environment 24(2):23-25. |
|
|
Stamford NP, Coelho LC, Araujo JM (2001). Production and characterization of a thermostable alpha-amylase from Nocardiopsis sp. endophyte of yam bean. Bioresources Technology 76:137-141. |
|
|
Stirling G, Helen H, Pattison T, Stirling M (2016). Introduction: Soil health, soil biology and sustainable agriculture and evidence based information Soil health. In: Peter Storer (ed), Soil Biology, Soil borne diseases and Sustainable Agriculture, 1st ed. Queensland Aus. Csiro publishing pp. 1-5. |
|
|
Sy A, Giraud E, Jourand P, Garcia N, Willems A, de Lajudie P, Prin Y, Neyra M, Gillis M, Boivin-Masson C, Dreyfus B (2001). Methylotrophic Methylobaterium bacteria nodulate and fix nitrogen in symbiosis with legumes. Journal of Bacteriology 183(1):214-220. |
|
|
Toyama H, Anthony C, Lidstrom ME (1998). Construction of insertion and deletion mxa mutants of Methylobacterium extorquens AM1 by electroporation. FEMS Microbiology Letters 166(1):1-7. |
|
|
Trotsenko IuA, Ivanova EG, Doronina NV (2001). Aerobic methylotrophic bacteria as phytosymbionts. Microbiology 70:623-632. |
|
|
Unica (2017). Brazilian sugarcane industry association. |
|
|
Verma SC, Ladha JK, Tripathi AK (2001). Evaluation of plant growth promotion and colonization ability of endophytic diazotrophs from deep-water rice. Journal of Biotechnology 91:127-141. |
|
|
Walitang DI, Kim K, Madhaiyan M, Kim YK, Kang Y, Sa T (2017). Characterizing endophytic competence and plant growth promotion of bacterial endophytes inhabiting the seed endosphere of Rice. BMC Microbiology 17:1-13. |
|
|
Wink M, Mohamed GIA (2003). Evolution of chemical defense traits in the Leguminosae: Mapping of distribution patterns of secondary metabolites on a molecular phylogeny inferred from nucleotide sequences of rbcL gene. Biochemical and Systematic Ecology 31:897-917. |
|
|
Zhang M, Lidstrom ME (2003). Promoters and transcripts for genes involved in methanol oxidation in Methylobacterium extorquens AM1. Microbiology 149(4):1033-1040. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


