Full Length Research Paper

ABSTRACT
The Gram-positive bacterium Enterococcus faecalis is an etiological agent of endodontic infections in oral biofilms. In the present study, we analyzed the effect of N-acyl homoserine lactones (AHLs) quorum sensing signal molecules on E. faecalis biofilm formation ability. Our results provide important effects of N-Hexanoyl-L-homoserine lactone (C6-HSL) and N-octanoyl-L-homoserine lactone (C8-HSL) molecules on E. faecalis biofilm formation. These AHLs molecules promoted the biofilm formation of E. faecalis.
Key words: N-acyl homoserine lactones (AHLs), bacterial communication, biofilm, Enterococcus faecalis, quorum sensing.
INTRODUCTION
Several species of bacteria employ a mechanism of intercellular communication known as quorum sensing (QS). This signaling process allows the cells comprising a bacterial colony to coordinate their gene expression in a cell-density dependent manner (Fuqua et al., 2001; Bassler and Losick, 2006). Thus, quorum sensing enables the cells within a bacterial colony to act co-operatively, facilitating population-dependent adaptive behavior (Atkinson and Williams, 2009). Quorum sensing has been shown to play a critical role in pathogenic bacteria-host interactions (Boyer and Wisniewski-Dyé, 2009).
Bacterial intercellular communication relies on autoinducers (AI), endogenous chemical compounds which are synthesized intracellularly throughout the growth of the bacteria and secreted into the environment. Some AI are bound to the membrane receptors located on the cell surface, while others may penetrate the membrane and bind to the intracellular receptors. After binding with the receptors, AI activate a broad spectrum of effector systems affecting the regulation of expression of a number of genes, including those encoding the synthesis and secretion of antibiotics, virulent factors, and formation of cell aggregates or surface biofilms (Whitehead et al., 2001; Sun et al., 2004; Waters and Bassler, 2005; Bassler and Losick, 2006; Camilli and Bassler, 2006; Steindler and Venturi, 2007).
The most widespread and important autoinducers are lactones of N-acylated homoserine (AI-1, AHL) (Galloway et al., 2011). AHLs are the most common class of autoinducer used by Gram-negative bacteria; indeed quorum sensing mediated by AHLs represents one of the best-understood bacterial systems at the molecular level (Geske et al., 2008).
Cyclic peptides are the major class of cell-cell signaling molecules in Gram-positive bacteria (Duan et al., 2003; Xavier and Bassler, 2003; Vendeville et al., 2005; De Keersmaecker et al., 2006). Furthermore, small-molecule effects can be controlled by altering the concentration of the small molecule, thereby allowing the generation of dose-response data (Geske et al., 2008; Galloway et al., 2011).
E. faecalis is a Gram-positive cocci; facultative anaerobic bacteria that is known to be the most frequently detected species in root canals with failed endodontic treatment (Wang et al., 2011). It has been reported that the quorum sensing system in E. faecalis is closely related to the biofilm formation (Lu et al., 2013). However, the effect of quorum sensing signal molecules N-Acyl homoserine lactones (AHLs) produced only by Gram-negative bacteria on biofilm formation of Gram-positive oral bacteria is not yet investigated and not well understood. The objective of this study was to investigate the effect of a quorum sensing signal molecules AHLs produced by Gram-negative bacteria on biofilm formation of a Gram-positive bacterium which is E. faecalis recovered from root canals of patients with endodontic infections.
MATERIALS AND METHODS
Bacterial strain, growth conditions and identification
E. faecalis strain was isolated from clinical samples of teeth of patients with endodontic infections in the dental surgery service of the University Hospital of Annaba (Algeria). E. faecalis was grown anaerobically (BD Gas Pak Anaerobe Generating Container System) at 37°C on M17 agar plates. M17 broth medium (Sigma-Aldrich) supplemented with 1% glucose was used for broth cultures (Kristich et al., 2004). Isolates, which were suspected as Enterococcus were immediately Gram stained to verify that they were Gram-positive, and analyzed for catalase-negative test (it is generally agreed that the genus Enterococcus comprises Gram-positive cocci that are catalase negative, usually facultative, anaerobic bacteria that grow in 6.5% NaCl, 40% bile salts. They grow at 10 and 45°C and can resist 30 min at 60°C) (Schleifer and Kilpper-Bälz, 1984; Schleifer and Kilpper-Bälz, 1987; Leclerc et al., 1996; Manero and Blanch, 1999). Isolates were identified to species level according to the standard biochemical tests using the API 20Strep system (bioMérieux, USA) for the identification of E. faecalis strain (Winston et al., 2004; Zoletti et al., 2006). The identification of E .faecalis strain was also confirmed using the mass spectrometry (MALDI-TOF) analysis.
AHLs molecules
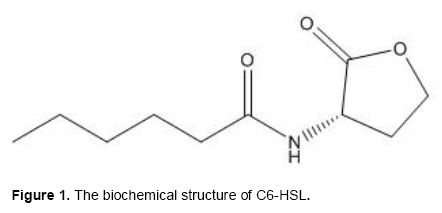
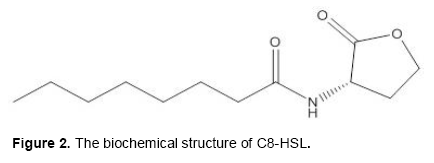
The AHLs quorum sensing signal molecules used in this study were purchased from Sigma-Aldrich (Spain); they are commercially available. In the present study, we will investigate the effect of two AHLs molecules: N-hexanoyl-L-homoserine lactone (C6-HSL) (Figure 1) and N- octanoyl-L-homoserine lactone (C8-HSL) (Figure 2) on biofilm formation of the Gram-positive bacterium E. faecalis. The concentration of each AHL molecule added to the M17 liquid culture of E. faecalis was 1 µL.
Biofilm formation
An automatic microplate reader was used to investigate the biofilm formation ability of E .faecalis strain. The microplates for biofilm cultivation were sterile 96-well polystyrene (Sterilin Limited, U.K).
The inoculum was prepared from E. faecalis grown in M17 broth supplemented with 1% glucose (weight/volume), and 200 µL is poured into the well (Christensen et al., 1985; Kennedy and O’Gara, 2004; Stepanovi? et al., 2007). The negative control wells (NC) (without AHLs) contain only 100 µL of E. faecalis culture in M17 broth supplemented with 1% glucose (initial optical density ODi at 600 nm was OD600 = 0.2 nm) + 100 µL of fresh and sterile M17 broth supplemented with 1% glucose per well. The positive control wells (PC1 and PC2) (with AHLs) contained 100 µL of E. faecalis culture in M17 broth supplemented with 1% glucose (OD600 = 0.2 nm) + 99 µL of fresh and sterile M17 broth supplemented with 1%glucose + 1 µL of the AHL molecule used (C6-HSL and C8-HSL) respectively per well (the positive control PC1 with C6-HSL; and the positive control PC2 with C8-HSL). The inoculated microplate was covered with a lid and incubated for 48 h of growth at 37°C, anaerobically using BD Gas Pak Anaerobe Generating Container System. After incubation, the wells in the microplate were emptied from the liquid culture of E. faecalis, then washed three times with a sterile saline solution of (NaCl 0.9 % weight/volume).
After washing, the remaining attached bacteria should be heat-fixed (drying completely) by exposing them to hot air at 56°C for 30 min; then the adherent biofilm layer formed on the bottom and walls of the well was stained; 200 µL of crystal violet 0.1% per well for 30 min at room temperature (Seneviratne et al., 2013). After staining, the wells were rinsed off by placing the microplate under running water. Washing was continued until the washings were free of the stain. Then, the microplate was air dried at room temperature. Then, the dye bound to the cells were resolubilized, that is eluted from attached cells with 95% ethanol (200 µL per well); ethanol was gently added and thereafter the microplate was covered with the lid (to minimize evaporation) and left at room temperature for at least 30 min without shaking (Branda et al., 2005). Finally, the optical density (OD) of each well stained with crystal violet was measured at 570 nm using the automatic microplatereader. Optical density at 570 nm (OD570) was measured to assess biofilm formation ability by the E. faecalis strain in each well. Then, the average in each lane (8 wells) of the microplate was calculated. This protocol was repeated three times to confirm the effect of AHLs used on E .faecalis biofilm formation.
RESULTS
Bacterial strain, growth conditions and identification
Mass spectrometry (MALDI-TOF) allowed the identi-fication of E. faecalis strain by analysis of its protein content and study of ionic species moving in elec-tromagnetic fields. The ions generated from the bacterial colony of E. faecalis spread on the steel metallic target, were separated according to their time of flight (TOF). It means the measurement of the time that ions took when they were subjected to an accelerating voltage to travel the length of the flight tube. The separation of ions depends mainly on their mass. For each group of ions that have the same ratio m/z (mass/charge), a signal was recorded in the detector as a time/intensity function. Thus, the set of peaks constituted a recorded mass spectrum. The spectra generated from the whole bacteria were then compared to reference spectra present in the data base of an expert system. Comparison of the spectrum of our strain E. faecalis (previously identified by the standard biochemical test using API 20 Strep system) was obtained from the mass spectrometer with thousands of reference spectra. The name of the bacterium displayed on the screen was: E. faecalis. The spectrum of E. faecalis was obtained after placing the plate containing the whole bacterial cells in the spectrometer.
Quantification of E. faecalis biofilm formed on polystyrene
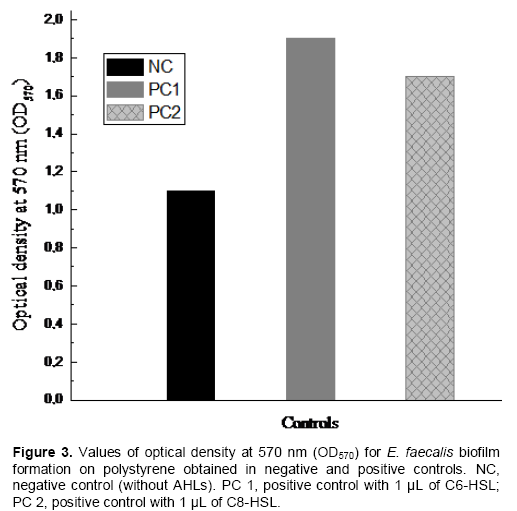
The results of the average of optical density at 570 nm (OD570) obtained were: In the negative control wells NC (without AHLs) OD570 = 1.1 nm; in the positive control wells PC1 (with 1 µL of C6-HSL molecule) OD570 = 1.9 nm; in the positive control wells PC2 (with 1 µL of C8-HSL molecule) OD570 = 1.7 nm.
The results mentioned above have shown that the addition of 1 µL of the AHLs molecules tested in this study (C6-HSL or C8-HSL) to the culture of E. faecalis in M17 broth supplemented with glucose 1% promoted the biofilm formation ability of E. faecalis strain (Figure 3). The values of the optical density at 570 nm (OD570) obtained in the negative and positive controls were the same in the three repetitions.
DISCUSSION
The E. faecalis bacteria have been identified as the most commonly recovered species from teeth with persistent endodontic infections (Martos et al., 2013). E. faecalis has also been implicated in persistent root canal infections (Preethee et al., 2012). Anaerobic bacteria, especially Gram-negative species of Prevotella and Porphyromonas, are frequently associated with signs and symptoms of endodontic origin (Haapasalo, 1989; Sundqvist et al., 1989; Hashioka et al., 1992), and facultative species such as E. faecalis have been isolated in high frequency from refractory cases (Hancock et al., 2001). The cell-to-cell communication of microorganisms is known to be via exertion of certain chemical compounds (signal molecules) and is referred to as quorum sensing. QS phenomenon is widespread in microbial communities (Safari et al., 2014). It is known that bacteria utilize a quorum sensing system to coordinate gene expression by monitoring the concentration of molecules known as AI (Shao et al., 2012). Bacteria use the QS mechanism to regulate a variety of phenotype, such as biofilm formation which is essential for the successful establishment of a symbiotic or pathogenic relationship with eukaryotic host cell (Myszka and Czaczyk, 2012). AHL-QS systems produce and respond to AHLs using two proteins that mediate signal production and response, LuxI and LuxR-like proteins, respectively (Nealson et al., 1970; Ruby, 1996). LuxR-like proteins are response regulators that mediate the expression of genes required for communal behavior in response to intracellular concentrations of cognate AHLs molecules (Fuqua and Winans, 1994; Fuqua et al., 1996; Cude and Buchan, 2013). Some bacterial pathogens are capable of colonizing infection sites and employing N-acyl homoserine lacones (AHLs) based quorum-sensing systems to co-ordinate biofilm formation (Bhargava et al., 2012). In the present study, we confirmed that the AHLs quorum sensing signal molecules (C6-HSL and C8-HSL) promoted the biofilm formation of the Gram-positive bacterium E. faecalis isolated from endodontic infections.
In 2012, a study realized in Beijing (China) at the institute of disease control and prevention, academy of military medical sciences, reported that the quorum sensing system in E. faecalis is based on AI-2 molecule.
Further, the role of AI-2 in biofilm formation by E. faecalis was investigated, showing that the addition of AI-2 molecule to E.f aecalis V583 cultures resulted in increased biofilm formation (Shao et al., 2012). Thus, the quorum sensing system in E. faecalis is closely related to the biofilm formation of this bacterium (Lu et al., 2013). From the results obtained in the present study, we suggest that the N-Acyl Homoserine Lactones AHLs molecules produced by Gram-negative bacteria can interfere with the quorum sensing system of a Gram-positive bacterium which is E. faecalis and consequently regulate the biofilm formation ability in E. faecalis strain. This suggestion confirms the possible interspecies com-munication between Gram-positive and Gram-negative bacteria.
Mature dental biofilms consist of towering microcolonies in which the resident bacterial cells interact with one another and exchange messages in the form of signaling molecules and metabolites. It is evident that mutually beneficial interactions between microbial cells are essential to the development of biofilms in the oral cavity (Jakubovics, 2010). Thus, the presence of Gram-negative bacteria in oral biofilms can influence the biofilm formation and growth of Gram-positive bacteria like E. faecalis by producing quorum sensing signal molecules (AHLs) and consequently promoting the development of endodontic infections.
In 2010, a study realized in Osaka (Japan) at the Department of Restorative Dentistry and Endodontology, Osaka University graduate school of dentistry, reported that three synthetic N-acyl homoserine lactones analogues (N-acyl HSL) inhibited the biofilm formation in the Gram-negative bacterium Porphyromonas gingivalis which is a primary agent of periodontal disease in oral biofilms (Asahi et al., 2010). Thus, the effects of N-acyl homoserine lactones molecules on bacterial biofilm formation are different depending on the Gram-positive or Gram-negative bacteria (Figure 4).
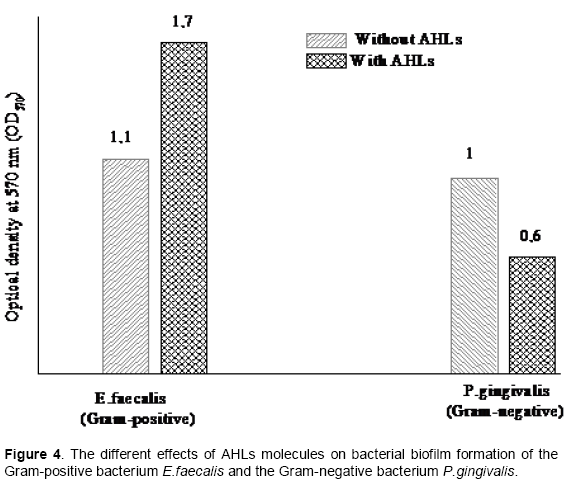
These different effects of the autoinducer molecules named AHLs on bacterial biofilm formation of the Gram-positive strain E. faecalis and the Gram-negative strain P. gingivalis confirms that the difference in biochemical composition of the bacterial cell wall between Gram-positive and Gram-negative bacteria plays a critical role in recognition and binding of the autoinducer molecule and also in the resulting effects of this molecule. The biochemical composition of the cell wall of E. faecalis strain has shown that the cell was endowed with a wall resistant to mechanical disruption, peptidoglycan, penicillin binding proteins (PBPs), muramidase enzyme and accessory wall polymers such as teichoic acid and lipoteichoic acid (Signoretto et al., 2000). The biochemical composition of cell-wall of Gram-positive bacteria has shown a very high proportion of the amino-acids such as: alanine, glutamic acid, lysine, diaminopimelic acid, aspartic acid and glycine. These were associated with varying combinations of sugars and amino sugars (Cummins and Harris, 1956). The Gram-negative bacteria possess a lipid-rich outer membrane (as well as a plasma membrane) and a thin peptidoglycan layer. Gram-positive bacteria are enshrouded in thicker, more resilient cell walls (Beveridge, 1999).
CONCLUSION
The present study has shown that quorum sensing signal molecules named “AHLs” can have a significant effect on oral Gram-positive bacteria by promoting the biofilm formation of E. faecalis strain isolated from teeth of patients with endodontic infections in Algeria. Furthermore, it was evident that these AHLs molecules act differently on biofilm formation of Gram-positive or Gram-negative oral bacteria. AHLs promoted the biofilm formation of the Gram-positive oral bacterium (E. faecalis) but inhibited the biofilm of the Gram-negative oral bacterium (P. gingivalis). Finally, we have to take advantage of these results to well understand the biological activity of these signal molecules in oral biofilms in order to minimize human oral infections.
CONFLICT OF INTEREST
The authors declare that they have no conflict of interest
ACKNOWLEDGEMENTS
We would like to thank Professor Jean Christophe Giard from the Laboratory Of Microbiology, Research Unit of Microbial Risks (U2RM), CHU Côte de Nacre, 14033 Caen Cedex, France for his technical assistance to identify the bacterial strain E. faecalis by MALDI-TOF (mass spectrometry) analysis.
REFERENCES
|
Asahi Y, Noiri Y, Igarashi J, Asai H, Suga H, Ebisu S (2010). Effects of N-acyl homoserine lactone analogues on Porphyromonas gingivalis biofilm formation. J. Periodontal. Res. 45(2):255-261. |
|
|
Atkinson S, Williams P (2009). Quorum sensing and social networking in the microbial world. J. R. Soc. Interface. 6 (40):959-978. |
|
|
Bassler BL, Losick R (2006). Bacterially speaking. Cell. 125(2):237-246. |
|
|
Beveridge TJ (1999). Structures of Gram-negative cell walls and their derived membrane vesicles. J. Bacteriol. 181(16):4725-4733. |
|
|
Bhargava N, Sharma P, Capalash N (2012). N-acyl homoserine lactone mediated interspecies interactions between A. baumannii and P. aeruginosa. Biofouling 28(8): 813-822. |
|
|
Boyer M, Wisniewski-Dyé F (2009). Cell-cell signalling in bacteria: Not simply a matter of quorum. FEMS. Microbiol. Ecol. 70(1):1-19. |
|
|
Branda SS, Vik A, Friedman L, Kolter R (2005). Biofilms: The matrix revisited. Trends Microbiol. 13 (1):20-26. |
|
|
Camilli A, Bassler BL (2006). Bacterial small-molecule signaling pathways. Science. 311(5764):1113-1116. |
|
|
Christensen GD, Simpson WA, Younger JJ, Baddour LM, Barrett FF, Melton DM, Beachey EH (1985). Adherence of coagulase-negative staphylococci to plastic tissue culture plates: A quantitative model for the adherence of staphylococci to medical devices. J. Clin. Microbiol. 22 (6):996-1006. |
|
|
Cude WN, Buchan A (2013). Acyl-homoserine lactone based quorum sensing in the Roseobacter clade: complex cell to cell communication controls multiple physiologies. Front. Microbiol. 4(336):1-12. |
|
|
Cummins CS, Harris H (1956). The chemical composition of the cell wall in some Gram-positive bacteria and its possible value as a taxonomic character. J. Gen. Microbiol. 14(3):583-600. |
|
|
De Keersmaecker SC, Sonck K, Vanderleyden J (2006). Let LuxS speak up in AI-2 signaling. Trends. Microbiol. 14(3):114-119. |
|
|
Duan K, Dammel C, Stein J, Rabin H, Surette MG (2003). Modulation of Pseudomonas aeruginosa gene expression by host microflora through interspecies communication. Mol. Microbiol. 50(5):1477-1491. |
|
|
Fuqua C, Parsek MR, Greenberg EP (2001). Regulation of gene expression by cell-to-cell communication: Acyl-homoserine lactone quorum sensing. Annu. Rev. Genet. 35 (1): 439-468. |
|
|
Fuqua C, Winans S, Greenberg EP (1996). Census and consensus in bacterial ecosystems: the LuxR-LuxI family of quorum-sensing transcriptional regulators. Annu. Rev. Microbiol. 50(1):727-751. |
|
|
Fuqua WC, Winans SC (1994). A LuxR-LuxI type regulatory system activates Agrobacterium Ti plasmid conjugal transferrin the presence of a plant tumor metabolite. J. Bacteriol. 176(10):2796-2806. |
|
|
Galloway WR, Hodgkinson JT, Bowden SD, Welch M, Spring DR (2011). Quorum sensing in Gram-negative bacteria: Small-molecule modulation of AHL and AI-2 quorum sensing pathways. Chem. Rev. 111(1):28-67. |
|
|
Geske GD, O'Neill JC, Blackwell HE (2008). Expanding dialogues: From natural autoinducers to non-natural analogues that modulate quorum sensing in Gram-negative bacteria. Chem. Soc. Rev. 37(7):1432-1447. |
|
|
Haapasalo M (1989). Bacteroides spp. in dental root canal infections. Endod. Dent. Traumatol. 5(1):1-10. |
|
|
Hancock HH, Sigurdsson A, Trope M, Moiseiwitsch J (2001). Bacteria isolated after unsuccessful endodontic treatment in a north american population. Oral. Surg. Oral. Med. Oral. Pathol. Oral. Radiol. Endod. 91(5):579-586. |
|
|
Hashioka K, Yamasaki M, Nakane A, Horiba N, Nakamura H (1992). The relationship between clinical symptoms and anaerobic bacteria from infected root canals. J. Endod. 18(11):558-561. |
|
|
Jakubovics NS (2010). Talk of the town: interspecies communication in oral biofilms. Mol. Oral. Microbiol. 25(1):4-14. |
|
|
Kennedy CA, O'Gara JP (2004). Contribution of culture media and chemical properties of polystyrene tissue culture plates to biofilm development by Staphylococcus aureus. J. Med. Microbiol. 53(11):1171-1173. |
|
|
Kristich CJ, Li YH, Cvitkovitch DG, Dunny GM (2004). Esp-independent biofilm formation by Enterococcus faecalis. J. Bacteriol. 186(1):154-163. |
|
|
Leclerc H, Devriese LA, Mossel DAA (1996). Taxonomical changes in intestinal (faecal) enterococci and streptococci: consequences on their use as indicators of faecal contamination in drinking water. J. Appl. Bacteriol. 81(5):459-466. |
|
|
Lu ZS, Meng L, Liu ZH, Ren GH, Sun AJ, Liu XM (2013). Expression of quorum-sensing related genes during Enterococcus faecalis biofilm formation. Chin. J. Stomatol. 48(8):485-489. |
|
|
Manero A, Blanch AR (1999). Identification of Enterococcus spp. with a biochemical key. Appl. Environ. Microbiol. 65(10):4425-4430. |
|
|
Martos J, FerrerLuque CM, González-Rodríguez MP, Arias-Moliz MT, Baca P (2013). Antimicrobial activity of essential oils and chloroform alone and combinated with cetrimide against Enterococcus faecalis biofilm. Eur. J. Microbiol. Immunol. 3(1):44-48. |
|
|
Myszka K, Czaczyk K (2012). N-acyl homoserine lactones AHLs as phenotype control factors produced by Gram-negative bacteria in natural ecosystems. Pol. J. Environ. Stud. 21(1):15-21. |
|
|
Nealson K H, Platt T, Hastings JW (1970). Cellular control of the synthesis and activity of the bacterial luminescent system. J. Bacteriol. 104(1):313-322. |
|
|
Preethee T, Kandaswamy D, Hannah R (2012). Molecular identification of an Enterococcus faecalis endocarditis antigen efaA in root canals of therapy-resistant endodontic infections. J. Conserv. Dent. 15(4):319-322. |
|
|
Ruby EG (1996). Lessons from a cooperative, bacterial-animal association: the Vibrio fischeri-Euprymnascolopes light organsymbiosis. Annu. Rev. Microbiol. 50(1):591-624. |
|
|
Safari M, Amache R, Esmaeilishirazifard E, Keshavarz T (2014). Microbial metabolism of quorum-sensing molecules acyl-homoserine lactones, γ-heptalactone and other lactones. Appl. Microbiol. Biotechnol. 98(8):3401-3412. |
|
|
Schleifer KH, Kilpper-Bälz R (1984). Transfer of Streptococcus faecalis and Streptococcus faecium to the genus Enterococcusnom.rev. as Enterococcus faecaliscomb.nov. and Enterococcus faeciumcomb.nov. Int. J. Syst. Bacteriol. 34(1): 31-34. |
|
|
Schleifer KH, Kilpper-Bälz R (1987). Molecular and chemotaxonomic approaches to the classification of streptococci, enterococci and lactococci: a review. Syst. Appl. Microbiol. 10(1):1-19. |
|
|
Seneviratne CJ, Yip JWY, Chang JWW, Zhang CF, Samaranayake LP (2013). Effect of culture media and nutrients on biofilm growth kinetics of laboratory and clinical strains of Enterococcus faecalis. Arch. Oral. Biol. 58(10):1327-1334. |
|
|
Shao C, Shang W, Yang Z, Sun Z, Li Y, Guo J, Wang X, Zou D (2012). LuxS-dependent AI-2 regulates versatile functions in Enterococcus faecalis V583. J. Proteome. Res. 11(9):4465-4475. |
|
|
Signoretto C, Del Mar Lleò M, Tafi MC, Canepari P (2000). Cell Wall Chemical Composition of Enterococcus faecalis in the Viable but Nonculturable State. Appl. Environ. Microbiol. 66(5):1953-1959. |
|
|
Steindler L, Venturi V (2007). Detection of quorum-sensing N-acyl homocysteine lactone signal molecules by bacterial biosensors. FEMS. Microbiol. Lett. 266(1):1-9. |
|
|
Stepanović S, Vuković D, Hola V, Di Bonaventura G, Djukić S, Cirković I, Ruzicka F (2007). Quantification of biofilm in microtiter plates: Overview of testing conditions and practical recommendations for assessment of biofilm production by Staphylococci. APMIS. 115(8):891-899. |
|
|
Sundqvist G, Johansson E, Sjögren U (1989). Prevalence of black pigmented Bacteriodes species in root canal infections. J. Endod. 15(1):13-19. |
|
|
Sun J, Daniel R, Wagner-Döbler I, Zeng AP (2004). Is autoinducer-2 a universal signal for interspecies communication: a comparative genomic and phylogenetic analysis of the synthesis and signal transduction pathways. BMC. Evol. Biol. 4(36):1-11. |
|
|
Vendeville A, Winzer K, Heurlier K, Tang CM, Hardie KR (2005). Making sense of metabolism: Autoinducer-2, LuxS and pathogenic bacteria. Nat. Rev. Microbiol. 3(5):383-396. |
|
|
Wang L, Dong M, Zheng J, Song Q, Yin W, Li J, Niu W (2011). Relationship of biofilm formation and gelE gene expression in Enterococcus faecalis recovered from root canals in patients requiring endodontic retreatment. J. Endod. 37(5):631-636. |
|
|
Waters CM, Bassler BL (2005). Quorum sensing: Cell-to-cell communication in bacteria. Annu. Rev. Cell. Dev. Biol. 21(1):319-346. |
|
|
Whitehead NA, Barnard AM, Slater H, Simpson NJ, Salmond GP (2001). Quorum-sensing in Gram-negative bacteria. FEMS. Microbiol. Rev. 25(4):365-404. |
|
|
Winston LG, Pang S, Haller BL, Wong M, Chambers HF, Perdreau-Remington F (2004). API 20 Strep identification system may incorrectly speciate enterococci with low level resistance to vancomycin. Diagn. Microbiol. Infect. Dis. 48(4):287-288. |
|
|
Xavier KB, Bassler BL (2003). LuxS quorum sensing: More than just a numbers game. Curr. Opin. Microbiol. 6(2):191-197. |
|
|
Zoletti GO, Siqueira JF, Santos KRN (2006). Identification of Enterococcus faecalis in root-filled teeth with or without periradicular lesions by culture-dependent and independent approaches. J. Endod. 32(8):722-726. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


