Full Length Research Paper

ABSTRACT
This study investigated the co-carriage of plasmid mediated quinolone resistance (PMQR) and extended spectrum beta-lactamase (ESBL) producing lactose non-fermenting (LNF) Enterobacteriaceae isolated from poultry birds. This was a descriptive cross-sectional study carried out between September, 2016 and March, 2017. The Kirby-Bauer disk diffusion method was used to determine the antimicrobial susceptibility patterns. ESBL screening disc kit was used to detect ESBL activities. Detection of ESBL and PMQR genes was carried out by means of polymerase chain reaction. In total, 207 LNF Enterobacteriaeae isolates were recovered from the cloacal swabs of poultry birds within the Calabar Metropolis. ESBL-producing isolates were 162 (78.3%) while fluroquinolone resistant isolates were 194 (93.7%). Among the ESBL-producing isolates, resistance to Ciprofloxacin, Norfloxacin, Levofloxacin, Ofloxacin and Nalidixic acid was 55 (34.2%), 26 (16.1%), 35 (21.7%), 50 (31.1%), and 162 (100%), respectively. About 65% of the quinolone resistant isolates were positive for at least one of the PMQR and ESBL genes in this study. Strict antimicrobial screening, surveillance of resistant isolates as well as the judicious practice of antimicrobial administration in the poultry setting with special emphasis on fluoroquinolones is advised given the high prevalence of co-existent ESBL and PMQR genes.
Key words: LNF enterobacteriaeae, Extended spectrum beta-lactamases, quinolone resistance
INTRODUCTION
MATERIALS AND METHODS
RESULTS
DISCUSSION
PMQR genes have been reported to be carried on mobile gene elements and can be easily transferred among different bacterial strains and species (Osińska et al., 2016; Redgrave et al., 2014). This potential exacerbates the development of multi drug resistance because PMQR reportedly reduces microbial susceptibility to antibiotics and supports the occurrence of resistance-associated mutations on bacterial chromosomes, thus making Enterebacteriaceae infections much more difficult to treat. In this study, we look into the co-carriage of PMQR and ESBL genes in LNF Enterebacteriaceae isolates in Calabar, Nigeria.
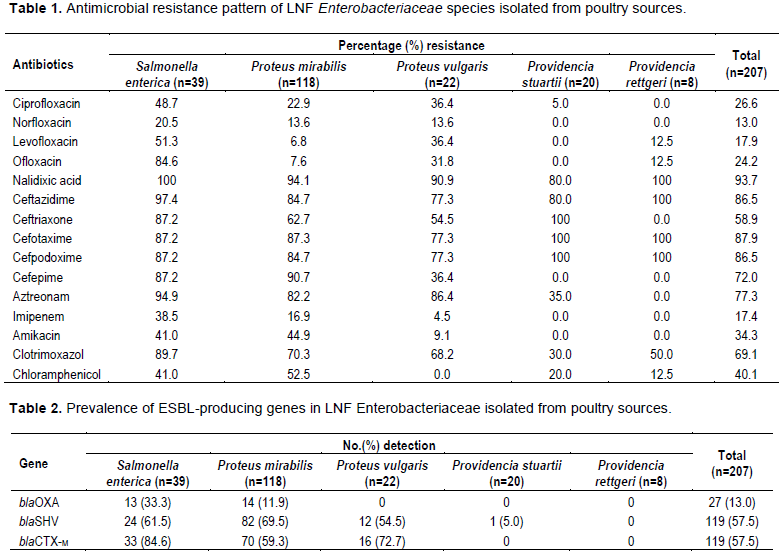
All 207 LNF Enterobacteriaceae demonstrated varied degrees of resistance to the 11 antibiotics used in this study. However, the isolates were most resistant to Nalixidic acid and were least resistant to Imipenem, Levofloxacin and Norfloxacin. This partially agrees with a similar study carried out in Azerbaijan and Iran on ESBL-PMQR co-carriage where resistance to Nalixidic acid was highest (68.5%) closely followed by resistance to Levofloxacin (55%) and Norfloxacin (65%) (Azargun et al., 2018). The injudicious and common use of Nalixidic acid in comparison to other fluroquinolones, Levofloxacin and Norfloxacin could be the reason for such high resistance to Nalixidic acid in this region. Hence we recommend strict selection and rotation of antimicrobial agents coupled with the continuous monitoring of susceptibility profiles of antimicrobial agents to determine best treatment options
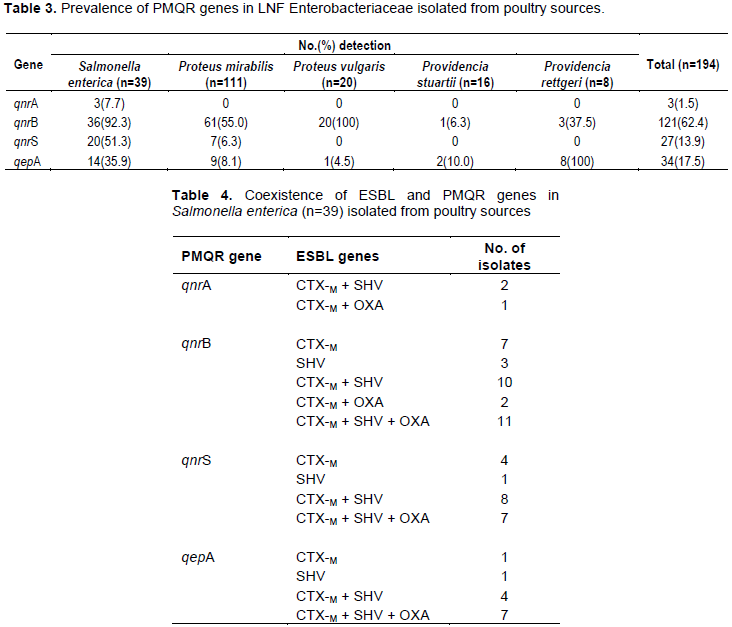
Our results further revealed that 78% (162) of the isolated produced ESBL and the most prominent genes of the 3 ESBL-producing genes tested were the blaSHV and the blaCTX-M. This correlates with several studies that have reported both genes as the most prevalent ESBL-producers (Giske et al. 2008)There was also high prevalence of PMQR genes (65%) among the 194 isolates that were resistant to fluroquinolones with qnrB being the most prevalent. This agrees with several studies that have suggested of the added advantage given by PMQR genes to fluoroquinolone resistance.
Upon further analysis, we discovered that resistance to fluoroquinolones was significantly higher among ESBL-producing isolates than non-ESBL-producing isolates. This implies that co-carriage of ESBL and PMQR genes is associated with ESBL-producing isolates. This result agrees with several studies in Asia and Africa but disagrees with a few other studies, as specific effects of PMQR co-carriage on treatment outcomes has been difficult to document (Jacoby et al., 2014; Jiang et al., 2012; Shakya et al., 2013)Nonetheless, such high prevalence of multidrug resistance among ESBL-producing isolates poses a serious challenge to antimicrobial therapy.
About 65.5% (127) of the quinolone resistant isolates were positive for at least one of the PMQR genes and ESBL genes used in this study. To further buttress on the issue of co-carriage, our study using S. enterica isolates demonstrated that among the 34 ESBL-producing isolates, 41% (14) carried at least one PMQR gene, that is, at least one PMQR gene was co-carried with ESBL gene in its plasmid. This was in consonance with a previous study in which a high proportion of PMQR genes was observed among isolates possessing the ESBL genes (17).
The association of ESBL genes and PMQR genes are of importance for public health concerns. However, this study did not include all the known ESBL genes and PMQR genes and molecular epidemiology was not performed, which could have further buttressed the evidences of the study. This study demonstrates high prevalence of LNF Enterobacteriaceae in a poultry setting. This agrees with several other studies done in USA and China (Projahn et al., 2018; Schwaiger et al., 2012). Due to the high prevalence of co-resistance to beta-lactam and fluoroquinolone antibiotics; we recommend the judicious practice of antimicrobial administration in the poultry setting with special emphasis on fluoroquinolones. We also recommend the continuous surveillance and monitoring of multidrug resistant isolates which should aid in proper antimicrobial administration in both humans and animals.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interest.
REFERENCES
|
Abrar S, Ain NU, Liaqat H, Hussain S, Rasheed F, Riaz S (2019). Distribution of bla CTX - M, bla TEM , bla SHV and bla OXA genes in Extended-spectrum-β-lactamase-producing Clinical isolates: A three-year multi-center study from Lahore, Pakistan. Antimicrobial resistance and infection control 8:80. |
|
|
Azargun R, Sadeghi MR, Soroush Barhaghi MH, Samadi Kafil H, Yeganeh F, Ahangar Oskouee M, Ghotaslou R (2018). The prevalence of plasmid-mediated quinolone resistance and ESBL-production in Enterobacteriaceae isolated from urinary tract infections. Infection and drug resistance 11:1007-1014. |
|
|
Bajaj P, Kanaujia PK, Singh NS, Sharma S, Kumar S, Virdi JS (2016). Quinolone co-resistance in ESBL- or AmpC-producing Escherichia coli from an Indian urban aquatic environment and their public health implications. Environmental Science and Pollution Research 23(2):1954-1959. |
|
|
Cheesbrough M (n.d.). District Laboratory Practice in Tropical Countries, Part 2 Second Edition, viewed 6 November 2018 View. |
|
|
Chen X, Zhang W, Pan W, Yin J, Pan Z, Gao S, Jiao X (2012). Prevalence of qnr, aac(6')-Ib-cr, qepA, and oqxAB in Escherichia coli isolates from humans, animals, and the environment. Antimicrobial agents and chemotherapy 56(6):3423-3427. |
|
|
Ferreira CM, Ferreira WA, Almeida NCO, da S, Naveca FG, Barbosa M, das GV (2011). Extended-spectrum beta-lactamase-producing bacteria isolated from hematologic patients in Manaus, State of Amazonas, Brazil. Brazilian journal of microbiology: [publication of the Brazilian Society for Microbiology] 42(3):1076-1084. |
|
|
Giske CG, Monnet DL, Cars O, Carmeli Y (2008). ReAct-Action on Antibiotic Resistance 2008, Clinical and Economic Impact of Common Multidrug-Resistant Gram-Negative Bacilli. Antimicrobial Agents and Chemotherapy 52(3):813-821. |
|
|
Jacoby GA, Strahilevitz J, Hooper DC (2014). Plasmid-mediated quinolone resistance. Microbiology spectrum 2(5). |
|
|
Jiang H-X, Tang D, Liu Y-H, Zhang X-H, Zeng Z-L, Xu L, Hawkey PM (2012). Prevalence and characteristics of -lactamase and plasmid-mediated quinolone resistance genes in Escherichia coli isolated from farmed fish in China. Journal of Antimicrobial Chemotherapy 67(10):2350-2353. |
|
|
Kao C-Y, Udval U, Huang Y-T, Wu H-M, Huang A-H, Bolormaa E, Yan J-J, Urangoo Z, Batbaatar G, Khosbayar T, Wu J-J (2016). Molecular characterization of extended-spectrum β-lactamase-producing Escherichia coli and Klebsiella spp. isolates in Mongolia. Journal of Microbiology, Immunology and Infection 49(5):692-700. |
|
|
Mathai D, Rhomberg PR, Biedenbach DJ, Jones RN (2002). India Antimicrobial Resistance Study Group 2002, Evaluation of the in vitro activity of six broad-spectrum beta-lactam antimicrobial agents tested against recent clinical isolates from India: a survey of ten medical center laboratories. Diagnostic microbiology and infectious disease 44(4):367-77. |
|
|
Ni Q, Tian Y, Zhang L, Jiang C, Dong D, Li Z, Mao E, Peng Y (2016). Prevalence and quinolone resistance of fecal carriage of extended-spectrum β-lactamase-producing Escherichia coli in 6 communities and 2 physical examination center populations in Shanghai, China. Diagnostic Microbiology and Infectious Disease 86(4):428-433. |
|
|
Oghenevo OJ, Bassey BE, Yhiler NY, Francis UM, Angela OE (2016). Antibiotic Resistance in Extended Spectrum Beta-Lactamases (Esbls) Salmonella Species Isolated from Patients with Diarrhoea in Calabar, Nigeria. Journal of Clinical Infectious Diseases and Practice 01(01):1-5. |
|
|
Orji MU, Onuigbo HC, Mbata TI (2005). Isolation of Salmonella from poultry droppings and other environmental sources in Awka, Nigeria. International Journal of Infectious Diseases 9(2):86-89. |
|
|
Osińska A, Harnisz M, Korzeniewska E (2016). Prevalence of plasmid-mediated multidrug resistance determinants in fluoroquinolone-resistant bacteria isolated from sewage and surface water. Environmental Science and Pollution Research 23(1):10818-10831. |
|
|
Pallecchi L, Bartoloni A, Riccobono E, Fernandez C, Mantella A, Magnelli D, Mannini D, Strohmeyer M, Bartalesi F, Rodriguez H, Gotuzzo E, Rossolini GM (2012). 'Quinolone Resistance in Absence of Selective Pressure: The Experience of a Very Remote Community in the Amazon Forest'. PLoS Neglected Tropical Diseases 6(8). |
|
|
Pourahmad Jaktaji R, Mohiti E (2010). Study of Mutations in the DNA gyrase gyrA Gene of Escherichia coli. Iranian Journal of Pharmaceutical Research 9(1):43-48. |
|
|
Projahn M, Pacholewicz E, Becker E, Correia-Carreira G, Bandick N, Kaesbohrer A (2018). Reviewing Interventions against Enterobacteriaceae in Broiler Processing: Using Old Techniques for Meeting the New Challenges of ESBL E. coli?. BioMed research international P 7309346. |
|
|
Redgrave LS, Sutton SB, Webber MA, Piddock LJV (2014). Fluoroquinolone resistance: mechanisms, impact on bacteria, and role in evolutionary success', Trends in Microbiology 22:438-445. |
|
|
Schwaiger K, Huther S, Hölzel C, Kämpf P, Bauer J (2012). Prevalence of antibiotic-resistant enterobacteriaceae isolated from chicken and pork meat purchased at the slaughterhouse and at retail in Bavaria, Germany. International journal of food microbiology 154(3):206-211. |
|
|
Schwarz S, Kehrenberg C, Walsh TR (2001). Use of antimicrobial agents in veterinary medicine and food animal production. International journal of antimicrobial agents 17(6):431-437. |
|
|
Shakya P, Barrett P, Diwan VM, Yogyata SH, Chhari N, Tamhankar AJ, Pathak A, Lundborg CS, Gootz T, Salyers A, Gupta A, Wang Y, Blake D, Hillman K, Fenlon D, Low J, Lester S, Pilar PD, Wang F, Schael IP, Jiang H, O'Brien T, Nys S, Okeke I, Kariuki S, Dinant G, Driessen C, Stobberingh E, Seidman J, Anitha K, Kanungo R, Bourgeois A, Coles C, Sahoo K, Tamhankar A, Sahoo S, Sahu P, Klintz S, Lundborg C, Pathak D, Pathak A, Marrone G, Diwan V, Lundborg C, Pathak A, Mahadik K, Dhaneria S, Sharma A, Eriksson B, Lundborg C, Fochsen G, Deshpande K, Diwan V, Mishra A, Diwan VK, Thorson A, Costa A, De Diwan V, Sabde Y, Diwan V, Saraf V, Mahadik V, Diwan VK, Costa A, De Turgeon M, Mathai E, Chandy K, Thomas K, Antoniswamy B, Joseph I, Mathai M, Sorensen T, Holloway K, Magiorakos A, Srinivasan A, Carey R, Carmeli Y, Falagas M, Giske C, Harbarth S, Hindler J, Kahlmeter G, Olsson-Liljequist B, Perez-Trallero E, Garcia-de-la-Fuente C, Garcia-Rey C, Baquero F, Aguilar L, Dal-Re R, Garcia-de-Lomas J, Sahuquillo-Arce J, Selva M, Perpinan H, Gobernado M, Armero C, Lopez-Quilez A, Gonzalez F, Vanaclocha H, Pathak A, Chandran S, Mahadik K, Macaden R, Stålsby L, Bartoloni A, Pallecchi L, Benedetti M, Fernandez C, Vallejos Y, Guzman E, Villagran A, Mantella A, Lucchetti C, Bartalesi F, Bartoloni A, Cutts F, Leoni S, Austin C, Mantella A, Guglielmetti P, Roselli M, Salazar E, Paradisi F, Bartoloni A, Bartalesi F, Mantella A, Dell'Amico E, Roselli M, Strohmeyer M, Barahona H, Barron V, Paradisi F, Rossolini G, Vatopoulos A, Varvaresou E, Petridou E, Moustaki M, Kyriakopoulos M, Kapogiannis D, Sarafoglou S, Fretzagias A, Kalapothaki V, Zaoutis T, Goyal M, Chu J, Coffin S, Bell L, Nachamkin I, McGowan K, Bilker W, Lautenbach E, Allin S, Stabile M, Kumar R, Indira K, Rizvi A, Rizvi T, Jeyaseelan L, Bloom S, Wypij D, Gupta M, Das Gaur A, Ramteke P, Pathak S, Bhattacherjee J, Diwan V, Tamhankar A, Khandal R, Sen S, Aggarwal M, Marothi Y, Iyer R, Sundblad-Tonderski K, Stalsby-Lundborg C, Diwan V, Tamhankar A, Aggarwal M, Sen S, Khandal R, Sen S, Stalsby-Lundborg C, Wright J, Gundry S, Conroy R (2013). Antibiotic resistance among Escherichia coli isolates from stool samples of children aged 3 to 14 years from Ujjain, India. BMC Infectious Diseases 13(1):477. |
|
|
Strahilevitz J, Jacoby GA, Hooper DC, Robicsek A (2009). Plasmid-Mediated Quinolone Resistance: a Multifaceted Threat. Clinical Microbiology Reviews 22(4):664-689. |
|
|
Sun J, Ke B, Huang Y, He D, Li X, Liang Z, Ke C (2014). The Molecular Epidemiological Characteristics and Genetic Diversity of Salmonella Typhimurium in Guangdong, China, 2007-2011', M.A. Webber (ed.) PLoS ONE 9(11):e113145. |
|
|
Tran JH, Jacoby GA (2002). Mechanism of plasmid-mediated quinolone resistance. Proceedings of the National Academy of Sciences of the United States of America 99(8):5638-5642. |
|
|
Xiong D, Song L, Pan Z, Jiao X (2018). Identification and Discrimination of Salmonella enterica Serovar Gallinarum Biovars Pullorum and Gallinarum Based on a One-Step Multiplex PCR Assay. Frontiers in Microbiology 9:1718. |
|
|
Yangkam YN, Bassey BE (2015). Antimicrobial Susceptibility Patterns of Salmonella Species from Sources in Poultry Production Settings in Calabar, Cross River State, Nigeria. American Journal of Health Research 3(2):76. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


