Full Length Research Paper

ABSTRACT
Fusarium oxysporum is an ubiquitous soil-borne fungus, having a high genetic and ecological diversity with the potential to cause diseases of many crop species of economic interest. Indeed, some strains of F. oxysporum known as pathogens generate common diseases such as wilting, root and crown rot on host plants. Two formae speciales are confined to the tomato: F. oxysporum f.sp. lycopersici (FOL) causing Fusarium wilt, while F. oxysporum f.sp. radicis-lycopersici (FORL) causes Fusarium crown and root rot. The study include 27 strains isolated from the stems, crown and roots of infected tomato plants; to confirm the identity of the fungus, the isolates were identified using analysis based on morphological criteria and sequencing of the translation elongation factor 1-alpha (TEF) gene using ef1 and ef2 primers. Twenty three strains belonged to F. oxysporum, three strains to Fusarium solani, and one strain to Fusarium redolens. Tomato seedlings were tested to confirm the pathogenicity of the isolates tested. Pathogenicity test confirmed that twenty two F. oxysporum isolates were pathogenic on tomato and produced crown and root rot typical of F. oxysporum f.sp. radicis-lycopersici forma specialis, while one strain initially identified as F. oxysporum did not induce disease symptoms and is considered as non-pathogenic. Additionally, no symptoms of Fusarium wilt were observed at all; therefore no strains can be affiliated to F. oxysporum f.sp. lycopersici forma specialis.
Key words: Fusarium oxysporum, tomato, molecular identification, pathogenicity.
INTRODUCTION
In Algeria, the cultivation of tomato (Solanum lycopersicum L.) occupies a privileged place in the socio-economic sector. It is in fact, regarded as a priority crop with a total area of 22497 ha (FAO, 2013). Although estimated at 975075 tonnes and 43.34 t/ha (FAO, 2013), the production remains low compared to other Mediterranean countries, this is partly due to some fungal diseases, such as Fusarium wilt caused by Fusarium oxysporum f.sp. lycopersici (Fol) (Snyder and Hansen, 1940). This pathogen invades the root epidermis and extends into the vascular tissue. It colonizes the xylem vessels producing mycelium and conidia. The characteristic wilt symptoms appear as a result of severe water stress due to vessel clogging (Beckman, 1987) or Fusarium crown and root rot caused by one of the most destructive pathogens; F. oxysporum f.sp. radicis lycopersici (Forl), the latter persists in soil (Benhamou et al., 1997) and produce a brown discoloration of roots, limited up to 10 to 30 cm above the ground.
The tap root of infected plants often rots partially or completely, and brown cankers which appear in the soil line. Infected plants can be stunted and withered. The infection can be spread by wounds and natural holes, contaminated seeds, microconidia from the air and infested soil or compost (Jarvis, 1988; Di Primo et al., 2001; Steinkellner et al., 2005; Koike et al., 2006). Actually, Fusarium crown and root rot was first discovered in Japan on 1969 (Menzies and Jarvis, 1994) and has spread worldwide since 1970 (Yamamoto et al., 1974) and recently appeared in Algeria.
This disease still causes massive damages in greenhouses and open fields, which incite us to characterize it in order to better understand its development and be able to control it. The aim of this study was to isolate F. oxysporum, to identify it using both morphological characteristics and DNA sequencing of the translation elongation factor 1-alpha gene (TEF), which appears to be consistently a single copy in Fusarium and shows a high level of sequence polymorphism among closely related species. For these reasons, TEF has become the marker of choice as a single-locus identification tool in Fusarium (Geiser et al., 2004; Kristensen et al., 2005). Additionally, the pathogenicity was also accessed.
MATERIALS AND METHODS
Isolation of fungi
Strains were isolated from tomato plants showing symptoms of wilt, crown and root rot. Samples were collected from the most productive regions of Algeria; from the West (Aïn Temouchent, Oran, Mostaganem, Mascara and Chlef), the East (Skikda, Guelma, Annaba and El Taref) and the South (El Oued). The roots, crowns and stems were first cut into small pieces, rinsed with distilled water, disinfected with sodium hypochlorite (5%) for 5 min and rinsed again with sterile distilled water to remove traces of bleach water and then dried using sterile filter papers.
The fragments were then cut lengthwise and placed in Petri dishes containing Potato Dextrose Agar (PDA) prepared as follows: potato infusion (made by boiling 200 g of sliced potatoes in 1 L of distilled water for 30 min) was thoroughly mixed with 20 g of Dextrose and 20 g of Agar powder. The pH was adjusted to 5.6 and the volume was completed to 1 L before autoclaving for 15 min at 121°C. The medium was mixed with 200 mg/L of Streptomycin to prevent bacterial proliferation (Davet and Rouxel, 1997; Rapilly, 1968). The plates were incubated at 25°C for 4 to 5 days. Strain purification was carried out by single spores culture according to Booth (1971). A drop of sterile distilled water was placed on a sterile slide under the dissecting microscope. An accumulation of spores is obtained on the wet tip of a needle and the point of the needle introduced into the drop of water on slide. The spores can be observed to flow from the tip of the needle into the water. When the suspension is adequate the needle can be withdrawn. Experience of the correct dilution can easily be obtained and is approximately the point when the spores are clearly distinguishable in the water and are not obscured by overlapping. The spore suspension on the slide is then picked up by a sterile loop and streaked across a 2% water agar plate. After 24 h of incubation at 25°C, the obtained germinating spores were then cultured on PDA.
Morphological identification
The morphological identification of strains was performed using Carnation Leaf Agar (CLA) medium based on the characteristics of microconidia, macroconidia, phialides and chlamydospores. Although this initial identification using morphological characters is important to sort the species into smaller groups. This is highly recommended to perform more advanced methods such as molecular identification (Leslie and Summerell, 2006).
Molecular identification
Differences in DNA sequences of genes have been used to support morphological identification of Fusarium sp. species (Harrow et al., 2010; Yli-Matilla et al., 2002). Therefore, strains were identified at the species level based on sequencing of the translation elongation factor 1-alpha gene using ef1 and ef2 primers (Geiser et al., 2004).
Mycelium from our pure isolates and from the pathogenic strain: F. oxysporum f.sp. radicis lycopersici NH48 (Genbank accession: JN222908) (Edel-Hermann et al., 2011) were harvested in order to extract DNA using E.Z.N.A. Fungal DNA Mini Kit (Omega Bio-Tek, Norcross, USA), according to the manufacturer protocol. The amplification of the translation elongation factor 1-alpha gene was performed using the following primers: ef1 (5’-ATG GGT AAG GA(A/G) GAC AAG AC-3’) and ef2 (5’-GGA (G/A)GT ACC AGT (G/C)AT CAT GTT-3’) (O’Donnell et al., 1998). The polymerase chain reaction (PCR) reaction mixture (25 μL) contained: 17.2 μL of H2O, 1.5 μL of 150 µM dNTPs mix, 2.5 μL Taq polymerase buffer + MgCl2 (10X), 0.2 μL (3 U) Taq DNA polymerase (Omega Bio-Tek, Norcross, USA), 1.3 μL (0.5 mM) each of the primers, and 1 μL of genomic DNA. A GeneAmp 9700 thermocycler (Applied Biosystems, Waltham, USA) was used for PCR amplifications with the following amplification cycles: initial denaturation at 94°C for 85 s followed by 35 cycles of denaturation at 95°C for 35 s, annealing at 59°C for 55 s and extension at 72°C for 90 s, followed by final extension for 10 min at 72°C. PCR products were detected by electrophoresis in a 1.5% agarose gel in TBE buffer (Edel-Hermann et al., 2011).
The PCR products were purified with the PureLink Quick PCR Purification Kit (Invitrogen, Carlsbad, USA) and sequenced using an Applied Biosystems 3500 Genetic Analyzer (Applied Biosystems, Waltham, USA). Sequence editing was performed with Molecular Evolutionary Genetics Analysis MEGA5 software (Kumar et al., 2008; Tamura et al., 2013). Sequences were identified via comparison with the GenBank (NCBI) database using the Blastn algorithm (Altschul et al., 1997; Mc Ginnis and Madden, 2004) and blast search against Fusarium-ID database (Geiser et al., 2004).
Pathogenicity test
To highlight the pathogenicity and to determine the forma specialis of strains, a pathogenicity test was performed on tomato seedlings, the variety Monfavet H63-5 F1 hybrid sensitive to the three races of F. oxysporum f.sp. lycopersici was used for this test. Isolates were grown in labeled tubes containing an inclined PDA medium. After 8 days of incubation, 2 mL of liquid minimal medium (LMM) were added to each tube. Thereafter, 1 mL of the suspension was transferred into a new tube containing 9 mL of LMM after agitation then, spore suspensions were filtered in labeled hemolysis tubes. The concentration of the conidial suspension of each isolate was adjusted to approximately 106 spores/mL.
The roots from two weeks old tomato seedlings (stage of emergence of the first true leaf) were cut to the 7 mm inferior edge and then soaked for 30 min in the conidial suspensions (03 seedlings per conidial suspension prepared from each isolate).
Controls consisted of seedling soaked in conidial suspension, prepared from the pathogenic strain NH48 as the positive control was soaked in sterile distilled water instead of conidial suspension as the negative control. The seedlings were then transplanted into pots containing sand previously autoclaved and grown in an experimental greenhouse at 25°C at a photoperiod of 12 h (Edel-Hermann et al., 2011). After 3 weeks, plants were uprooted. The lower stem and taproot were longitudinally sectioned for examination of internal tissues (Kim et al., 2001). Symptom evaluation was made based on a rating scale of symptoms proposed by Vakalounakis and Fragkiadakis (1999). 0 = No symptoms; 1 = Light yellowing of leaves, light or moderate rot on taproot and secondary roots and crown rot; 2 = Moderate or severe yellowing of leaves with or without wilting, stunting, severe rot on taproot and secondary roots, crown rot with or without hypocotyls rot and vascular discoloration in the stem; 3 = Dead of seedlings. Disease incidence percentage was determined using the following formula (Song et al., 2004):
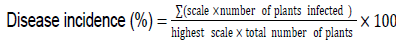
RESULTS AND DISCUSSION
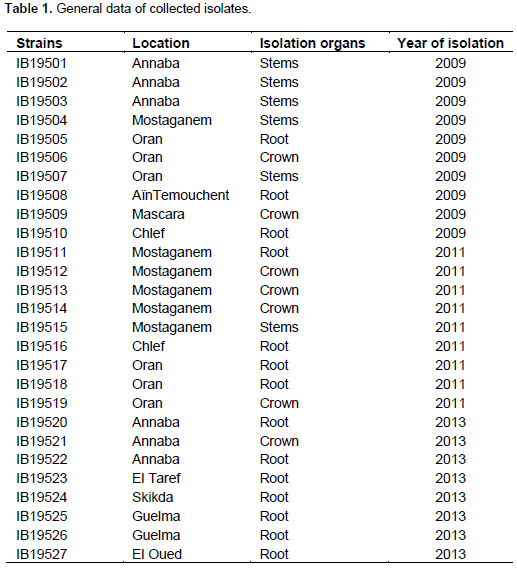
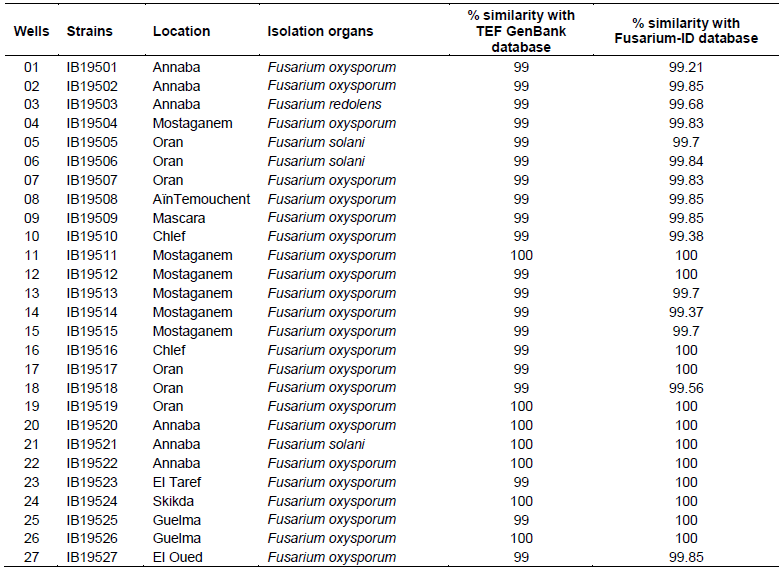
A total of 27 strains were obtained from the Western, Eastern and Southern parts of Algeria. Their origins, the year and the part of the plant from which isolation was performed are shown in Table 1.
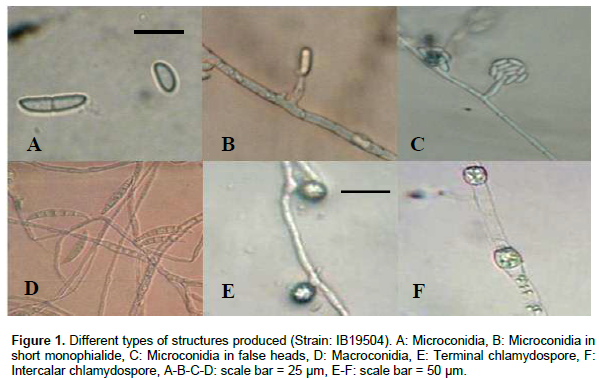
Gerlach and Nirenberg (1982) and Nelson et al. (1983) described the colony appearance of F. oxysporum on PDA as highly variable. Indeed, peach and violet colony colors, observed in the majority of isolates were consistent with the description of Booth (1971). Morphological studies showed Microconidia with zero to one septate; oval or cylindrical (Figure 1A), produced on short monophialides (Figure 1B) or in false heads (Figure 1C). Macroconidia were short to medium, falcate to straight and usually with three to five septate (Figure 1D). Chlamydospores were produced abundantly in terminal (Figure 1E) and intercalary chains (Figure 1F) in 3 to 6 week-old cultures. All these correspond to the critical morphological features of F. oxysporum species cited by Leslie and Summerell (2006).
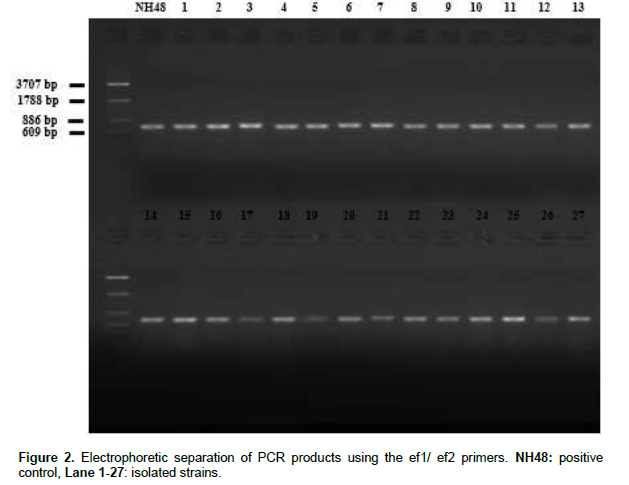
The (TEF) PCR products generated using the ef1 and ef2 primers were analyzed on agarose electrophoresis gel (Figure 2), an amplicon of an approximately 700 bp was produced with all strains. The Blastn search for similarities in GenBank database indicated between 99 and 100% identity while the blast against Fusarium-ID database indicated between 99.85 and 100% identity for all isolates. Therefore, the species name assigned was according to the closest match with known species. The combination of these two tools gave similar results and allowed us to confirm the identity of our isolates, as follow: twenty three strains identified as F. oxysporum, three strains as Fusarium solani and one strain as Fusarium redolens (Table 2).
The pathogenicity test conducted on 15 days old plants with 3 replicates per strain showed that twenty two strains identified by their translation elongation factor 1-alpha gene as F. oxysporum were pathogenic and have produced severe crown and root rot as the positive control NH48 (Figure 3B 1-2). The longitudinal section of stems showed brown discoloration of the vascular system extending no more than 10 to 15 cm above the soil as a consequence of the pathogen invasion (Figure 3C). Infected plants also showed a proliferation of adventitious roots on the first 10 cm of stem above the soil line (Figure 3B3). These symptoms are typical of F. oxysporum f.sp. radicis-lycopersici (FORL) as described by Jarvis and Shoemaker (1978), and different, both in varietal reaction and symptomatology from the Fusarium wilt caused by F. oxysporum f.sp. lycopersici (FOL); for the latter, above ground symptoms are commonly apparent, plants are severely stunted and wilted and some of them collapsed and died within 3-4 weeks. Plants showed severe vascular discoloration extending well up the stem until the petiole of the lower leaves, this contrasted with the discoloration in crown and root rot which was limited to the lower stem near the soil line. Indeed, since then Yamamoto et al. (1974) described the Fusarium crown and root rot of tomato in Japan and named the pathogen as a new race of F. oxysporum f.sp. lycopersici, there has been some disagreement with this designation because the symptoms are not those typical of Fusarium wilt but of the Fusarium crown and root rot (Jarvis et al., 1977; Rowe et al., 1977; Nutter et al., 1978). These results suggest that the twenty two F. oxyporum strains belong to the forma specialis F. oxysporum f. sp. radicis-lycopersici with 66.67% as the highest disease incidence and 44.44% for the lowest one (Table 3).

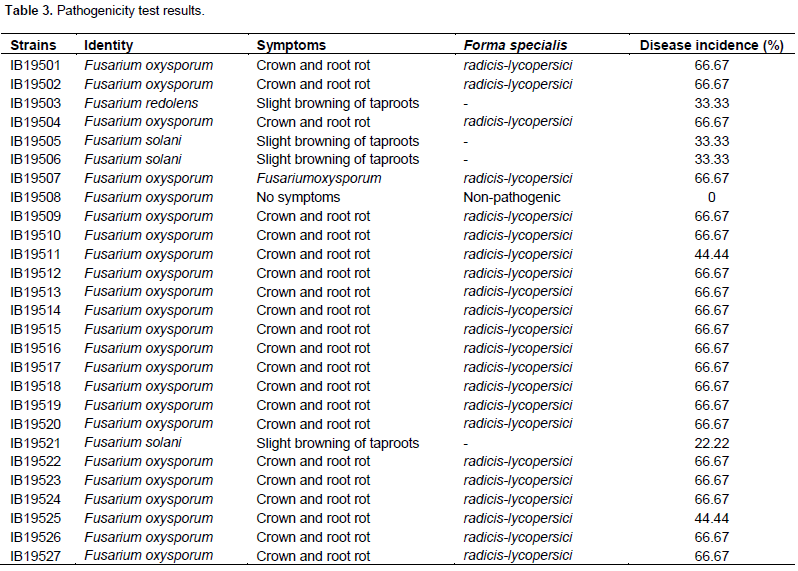
However, the strains IB19505, IB19506 and IB19521 identified as F. solani and IB19503 identified as F. redolens have produced a slight browning of taproots and showed between 22.22 and 33.33% of disease incidence (Table 3). The strain IB19508, initially identified as F. oxysporum did not cause any symptoms and is therefore considered as non-pathogenic. The re-isolation of pathogens from stems, crowns and roots of each plant showing symptoms, confirmed that it is the same starting strains that were used to prepare initial inoculums and allowed us to establish a causative relationship between isolates and the disease. This, confirm the Koch's postulates.
CONCLUSION
The Mediterranean countries of the Maghreb, mainly Algeria where cultivation of tomatoes is very important, are facing an increase in Fusarium wilt and Fusarium crown and root rot. This scourge is favored by climate changes like rising temperatures throughout the year and increasing humidity which promote fungal growth. As a part of our study, the identity of the fungus was confirmed using the translation elongation factor 1-alpha gene and the pathogenicity of the isolates was revealed when tomato plants were tested. The obtained results allowed us to distinguish strains belonging to F. redolens and F. solani species in addition to strains belonging to F. oxysporum which produced typical symptoms of Fusarium crown and root rot and a non-pathogenic F. oxysporum strain, that could be used as a potential biological control agent to protect tomato crops from these pathogens and also to limit the use of pesticides, which fits perfectly with a respectful environment approach.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Altschul SF, Madden TL, Schäffer AA, Zhang J, Zhang Z, Miller W, Lipman DJ (1997). Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 25:3389-3402. |
|
|
Beckman CM (1987). The nature of wilt disease. APS Press. St Paul. Minnesota. USA. 175 p. |
|
|
Benhamou N, Rey P, Cherif M, Hockenhul J, Tirilly Y (1997). Treatment with the mycoparasite Pythium oligandrum triggers induction of defence-related reactions in tomato roots when challenged with Fusarium oxysporum f. sp. radicis-lycopersici. Phytopathology 87:108-121. |
|
|
Booth C (1971). The genus Fusarium. Commonwealth Mycological Institute. Kew. Surrey. England. p237. |
|
|
Davet P, Rouxel F (1997). Détection et isolement des champignons du sol. INRA. Paris. France. p203. |
|
|
Di Primo P, Cartia G, Katan T (2001). Vegetative compatibility and heterokaryon stability in Fusarium oxysporum f.sp. radicis-lycopersici from Italy. Plant. Pathol. 50:371-382. |
|
|
Edel-Hermann V, Gautheron N, Steinberg C (2011). Genetic diversity of Fusarium oxysporum and related species pathogenic on tomato in Algeria and other Mediterranean countries. Plant. Pathol. 61:787-800. |
|
|
FAO (2013). Food and agriculture organization of the United Nations: statistics division. View. Retrieved 25 January 2015. |
|
|
Geiser DM, Jimenez-Gasco M, Kang S, Makalowska I, Veeraraghavan N, Ward TJ, Zhang N, Kuldau GA, O'Donnell K (2004). FUSARIUM-ID v. 1.0: A DNA sequence database for identifying Fusarium. Eur. J. Plant. Pathol. 110:473-479. |
|
|
Gerlach W, Nirenberg HI (1982). The genus Fusarium: a pictorial atlas. Mitt. Biol. Bundesanst. Ld. u. Forstw. Berlin-Dahlem. 209:1-406. |
|
|
Harrow SA, Ferrokhi-Nejad R, Pitman AR, Scott IAW, Bentley A, Hide C, Cromey MG (2010). Characterization of New Zealand Fusarium populations using a polyphasic approach differentiates the F. avenaceum/F. acuminatum/F. tricinctum species complex in cereal and grassland systems. Fungal. Biol. 114:293-311. |
|
|
Jarvis WR (1988). Fusarium crown and root rot of tomatoes. Phytoprotection. 69:49-64. |
|
|
Jarvis WR, Dirks VA, Johnson PW, Thorpe HJ (1977). No interaction between root knot nematode and Fusarium foot and root rot of greenhouse tomato. Plant. Dis. Rep. 61:251-254. |
|
|
Jarvis WR, Shoemaker RA (1978). Taxonomic status of Fusarium oxysporum causing foot and root rot of tomato. Phytopathology. 68:1679-1680. |
|
|
Kim JT, Park IH, Hahm YL, Yu SH (2001). Crown and root rot of greenhouse tomato caused by Fusarium oxysporum f. sp. radicis-lycopersici in Korea. Plant. Pathol. J. 17:290-294. |
|
|
Koike ST, Gladders P, Paulus AO (2006). Vegetable diseases: A color handbook. Academic Press. Elsevier. London. UK. 320 p. |
|
|
Kristensen R, Torp M, Kosiak B, Holst-Jensen A (2005). Phylogeny and toxigenic potential is correlated in Fusarium species as revealed by partial translation elongation factor 1 alpha gene sequences. Mycol. Res. 109:173-186. |
|
|
Kumar S, Dudley J, Nei M, Tamura K (2008). MEGA: A biologist-centric software for evolutionary analysis of DNA and protein sequences. Brief. Bioinform. 9:299-306. |
|
|
Leslie JF, Summerell BA (2006). The Fusarium laboratory manual. Blackwell Publishing. Iowa. USA. 388 p. |
|
|
Mc Ginnis S, Madden TL (2004). BLAST: At the core of a powerful and diverse set of sequence analysis tools. Nucleic. Acids. Res. 32:20-25. |
|
|
Menzies JG, Jarvis WR (1994). The infestation of tomato seed by Fusarium oxysporum f. sp. radicis-lycopersici. Plant. Pathol. 43:378-386. |
|
|
Nelson PE, Tousson TA, Marasas WFO (1983). Fusarium species: An illustrated manual for Identification. Pennsylvania State University Press. University Park. PA. USA. 206 p. |
|
|
Nutter FW, Warren CG, Wells OS, MacHardy WE (1978). Fusarium foot and root rot of tomato in New Hampshire. Plant. Dis. Rep. 62:976-978. |
|
|
O'Donnell K, Kistler HC, Cigelnik E, Ploetz RC (1998). Multiple evolutionary origins of the fungus causing Panama disease of banana: Concordant evidence from nuclear and mitochondrial gene genealogies. Proc. Natl. Acad. Sci. USA. 95:2044-2049. |
|
|
Rapilly F (1968). Les techniques de mycologie en pathologie végétale. Ann. Epiphyt. INRA. Paris. France. 102 p. |
|
|
Rowe RC, Farley JD, Colpin DL (1977). Air-borne spore dispersal and recolonization of steamed soil Fusarium oxysporum in tomato greenhouse. Phytopathology 67:1513-1517. |
|
|
Snyder WC, Hansen HN (1940). The species concept in Fusarium. Am. J. Bot. 27:64-67. |
|
|
Song W, Zhou L, Yang C, Cao X, Zhang L, Liu X (2004). Tomato Fusarium wilt and its chemical control strategies in a hydroponic system. Crop. Prot. 23:243-247. |
|
|
Steinkellner S, Mammerler R, Vierheilig H (2005). Microconidia germination of the tomato pathogen Fusarium oxysporum in the presence of root exudates. J. Plant. Inter. 1:23-30 |
|
|
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013). MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 30:2725-2729. |
|
|
Vakalounakis DJ, Fragkiadakis GA (1999). Genetic diversity of Fusarium oxysporum isolates from cucumber: Differentiation by pathogenicity, vegetative compatibility and RAPD fingerprinting. Phytopathology 89:161-168. |
|
|
Yamamoto I, Komada H, Kuniyasu K, Saito M, Ezuka A (1974). A new race of Fusarium oxysporum f. sp. lycopersici inducing root rot of tomato. Proc. Kansai. Plant. Prost. Soc. 16:17-29. |
|
|
Yli-Matilla T, Paavaeen-Huhtala S, Bulat SA, Alekhina IA, Nirenberg HI (2002). Molecular, morphological and phylogenetic analysis of the Fusarium avenaceum / F. arthrosporioides / F. tricinctum species complex: A polyphasic approach. Mycol. Res. 106:655-669. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


