Full Length Research Paper

ABSTRACT
In the dairy industry, the biofilms formed by spore forming bacteria are not well characterized. Microscopic methods are crucial for the study of biofilm structural and architectural features. Here, a simple surface-associated non-submerged model combined to environmental scanning electron microscope (ESEM) imaging was used for the study of Bacillus cereus and Geobacillus spp. dairy biofilms. To evaluate the utility of this approach, non-submerged biofilms were compared to those developed in situ on stainless steel coupons introduced inside milk processing lines. Results reveal that both B. cereus and Geobacillus spp. are able to form specific biofilm characteristics on non-immersed surfaces, notably an original dispersion style not previously described. Non-submerged biofilms in vitro are elaborate three-dimensional or extensive complex structures well resolved in ESEM and comparable to dairy biofilms in situ. The non-submerged surface-associated biofilm combined to ESEM imaging revealed a relevant model for the study of dairy biofilms.
Key words: Biofilms, Geobacillus, Bacillus cereus, non-submerged surfaces, environmental scanning electron microscope (ESEM) imaging.
INTRODUCTION
In the dairy industry, the biofilms formed on equipment surfaces are recognized to be a major source of contamination of processed milk and dairy products with both spoilage and pathogenic bacteria (Austin and Bergeron, 1995; Marchand et al., 2012; Gopal et al., 2015), and are difficult to remove even with acceptable cleaning procedures (Bremer et al., 2006). For several years, numerous studies focused on optimizing cleaning-in-place (CIP) systems, by using different biofilm models (Bénézech et al., 2002; Parkar et al., 2004; Faille et al., 2013; Kumari and Sarkar, 2014). General systems for the study of dairy biofilms often comprise complex settings that differ from laboratory to laboratory. This illustrates the need for techniques that rapidly and accurately characterize these biofilms and provide reproducible data with regard to their prevention and control. Biofilms that form at air-liquid interfaces are relevant models for the study of biofilm formation in aerobic spore forming bacteria such as Bacillus cereus (Wijman et al., 2007) and the thermophilic Geobacillus or Anoxybacillus (Zhao et al., 2013). Another simple model is the microorganism carrier-surface method previously described by Maris (1992) for testing the effectiveness of sanitizers. The formation of non-submerged biofilms on open surfaces is a practical method that was also used in other biofilm studies (Sommer et al., 1999; Leriche and Carpentier, 2000).
Microscopic methods are crucial for analysis of surface-associated communities. High-resolution imaging such as scanning electron microscopy (SEM), or confocal laser scanning microscopy (CLSM) proved to be powerful tools for biofilm structure deciphering, but they require extensive sample preparation (SEM) or image analysis (CLSM) and are not suitable for routine use. An easier microscopic approach is the use of environmental scanning electron microscopy (ESEM), which allows the observation of microbial biofilms in their native state without fixation, dehydration or metal coating (Little et al., 1991; Alhede et al., 2012). The main advantages of biofilm ESEM imaging are, indeed, the visualization of the highly hydrated organic matrix surrounding the bacteria, and minimal processing of samples.
In order to have, in a simple and rapid way, an insight into authentic biofilms that develop on dairy processing equipment surfaces, this study was carried out. A set of 13 strains of B. cereus and 10 strains of Geobacillus spp. of dairy origin were used to form biofilms on non-immersed stainless steel coupons, according to the method of Maris (1992). Non-submerged biofilms were further readily observed in ESEM. To evaluate the utility of this approach, non-submerged biofilms were compared to dairy biofilms in situ, inside milk processing lines.
MATERIALS AND METHODS
Bacterial strains culture and media
B. cereus were selected strains of genotypes that recurred for several years in a pasteurized milk processing line (Malek et al., 2013) whilst Geobacillus spp. are recent isolates from a monthly sampling performed in the same processing line (Table 1). Pipe-line surfaces were sampled with sterile swabs and dilution series made in TSE (0.1% trypton, 0.9% NaCl), plated on trypton-soy agar (TSA) medium (Sigma-Aldrich) and incubated at 55°C. Identification to Geobacillus spp. was performed according to thermophilic bacilli characteristics summarized in Burgess et al. (2010). Both B. cereus and Geobacillus spp. dairy isolates were characterized by high biofilm-forming potential on stainless steel coupons as revealed by plate count in agar media, respectively at 30 and 42°C (data not shown).
Preparation of stainless steel coupons and spore suspension
Stainless steel chips (AISI 304 L, 2 × 2 cm) were treated according to the protocol described by Peng et al. (2001). Spore suspensions of B. cereus and Geobacillus spp. were respectively obtained on Luria-Bertani agar (LB) and tryptic-soy-agar (TSA) media (Sigma-Aldrich), according to the method of Simmonds et al. (2003), and prior to use, they were washed one time and suspended in saline (0.15 M) at pH 7.4.
Biofilm formation on non-submerged stainless steel coupons
Biofilms were performed on stainless steel chips, as previously described (Maris, 1992). Briefly, 100 µL of B. cereus or Geobacillus spp. spore suspensions (107 - 109 spores.mL-1) were deposited onto clean and sterilized stainless steel coupons and allowed to adhere respectively at 30°C, and 45°C in a humidity-saturated atmosphere, for 3 h. Coupons were further washed with distilled water to remove weakly adhered cells, and then 100 µL of biofilm culture medium was poured over the adhesion area. Two culture media were used, the highly nutritious TSB medium (Sigma-Aldrich) and comparatively lesser nutritious nutrient broth Conda Pronadisa medium. Fouling coupons were incubated at appropriate temperatures for 20 to 24 h. In the case of the spore deficient strain B. cereus BC3 (Table 1), vegetative cells obtained from an overnight culture in LB broth, were harvested by centrifugation, washed with saline solution, and used to form biofilm on stainless steel coupon as described for spore suspensions.
Biofilm formation in co-culture
For biofilm formation in co-culture, spore suspensions (107 spores.mL-1, v/v) of a strain of B. cereus (BC15, or BC9) and a strain of Geobacillus spp. (HP, or EP) were mixed in an eppendorf tube and then a drop of 100 µL was deposited onto stainless steel chips as described previously. The coupons were incubated at 45°C in a humidity-saturated atmosphere. This temperature which revealed suitable for biofilm formation by these Geobacillus spp. dairy isolates also suits for the mesophilic B. cereus goups III and IV previously described to grow between 15 and 45°C (Guinebretière et al., 2008).
Biofilm formation in situ inside milking pipes
Milk processing line surfaces from which B. cereus and Geobacillus spp. strains originated, were investigated for the formation of biofilms. The observation of biofilms in-situ requires the installation of tools or devices into processing lines. To achieve this in an easier manner, stainless steel coupons (1 × 2.5 cm) were pierced at their ends making either one or two holes of 3 mm diameter, cleaned and sterilized as described previously, and suspended, in several areas inside milking pipes, notably pre and post-pasteurization sections. After a week, the coupons were removed from pipes, 7 h after cleaning-in-place (CIP) procedures, that is, just before a new production run. The CIP regimen followed at the investigated dairy plant consists of alkaline wash (2% NaOH) for 10 min at 80°C and acid wash (1% HNO3) for 5 min at 70°C.
Microscopy
All the above described biofilm carrying stainless steels coupons were gently rinsed with distilled water at the end of incubation time and examined in a 100 TM Hitachi environmental scanning electron microscope, at pressure in microscope chamber of 4 Torr.
RESULTS
Non-submerged 20 h old biofilms produced on stainless steel coupons by B. cereus or Geobacillus spp. dairy isolates were elaborate compact or more extensive complex structures, well resolved in ESEM. A selection of the best replicated images of the biofilms produced by representative strains of B. cereus (BC2, BC4, BC15, BC1, BC5, BC16 and BC3) and representative strains of Geobacillus spp. (LPM, H11, HP, EP and H17) was chosen to be presented here. For both bacteria, ESEM images showed either intact mature biofilms or dispersion process. In B. cereus, dispersion occurred in nutrient broth, whereas TSB medium supported the development of substantial mature biofilms. In comparison, biofilm dispersal was observed in both media for Geobacillus spp. strains. It should be noted that this simple biofilm non-submerged model, produced wide structure diversity, and was efficient for analysis of biofilms at various developmental stages.
Dispersion from B. cereus biofilms
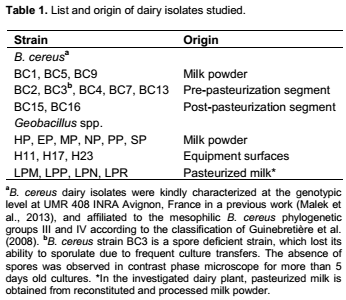
Figure 1 shows dispersion from luxurious biofilms which consisted of dense layers of rod-shaped cells interspersed with extracellular polymeric substances (EPS) material better visualized at higher resolution (Figure 1d). Dispersion occurred either from the outer surfaces of high colony diameter mushroom-like biofilms (Figure 1a) or through the formation of hollow central cavities characteristic of seeding dispersal (Figure 1c). Individual vegetative cells or small clusters of cells, rarely spores, were released from these biofilms and observed invading the substratum. At more advanced stages of dispersal process (Figure 1e and f), spores probably located in deeper areas within the biofilm, were released. The end of biofilm dispersal was imaged as cell-free EPS debris (Figure 1f).
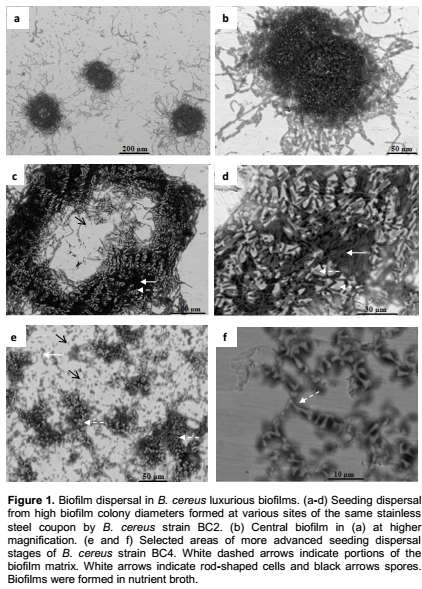
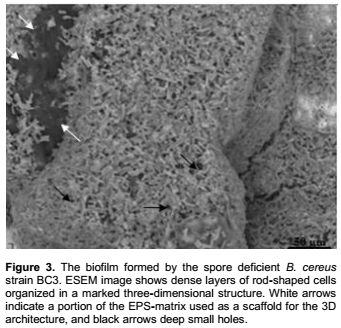
Similarly, several matrix-rich biofilms were imaged in ESEM at the dispersion stage (Figure 2). The mosaic biofilm shown in Figure 2a was characterized by irregular matrix blocks stacked on each other, delimiting large voids from which spores were released. Spores also dispersed from artistically shaped biofilm matrix (Figure 2c to d) and beside spores, vegetative cells were released from the pillar-like biofilm (Figure 2c), as shown following biofilm breaking (Figure 2d). In this biofilm, it appears that both vegetative cells and spores were rather embedded within the amorphous dark grey EPS-material, visualized under light grey pillar-like structures. This is clearly suggested in Figure 2d, where void areas resulting from the dissolution of dark grey material are observed, without obvious alteration of the pillar-like structures, at this stage. These results clearly indicate that spores were more present in matrix-rich B. cereus biofilms, in comparison to the above described luxurious biofilms. In addition the biofilm formed by the spore deficient B. cereus strain BC3, consisted of dense layers of rod-shaped cells, organized in a marked three-dimensional architecture without obvious spores and with nearly no visible EPS-matrix (Figure 3).

Mushroom-like and heterogeneous structures of B. cereus biofilms
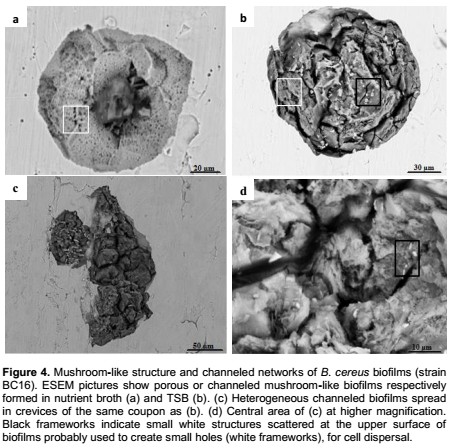
In comparison to nutrient broth, B. cereus developed densely packed complex biofilms in the highly nutritious TSB medium. Figure 4b shows a complex biofilm characterized by a heterogeneous mushroom-like structure, penetrated by interstitials voids and large deep water channels. ESEM images clearly depicted this remarkable network of well-defined channels which either extended from the top to the bottom of the biofilm (Figure 4b) or occurred at its surface (Figure 4c). It was also easy to view the marked widening and profound depth of these galleries. We can also see other heterogeneous topographical characteristics of these complex mature biofilms where cells were entirely encased in the EPS matrix. In contrast, Figure 4a showed another mushroom-like biofilm characterized by a compact porous structure, formed in comparatively more limited nutritious medium (Nutrient broth).
Dispersion from Geobacillus spp. biofilm
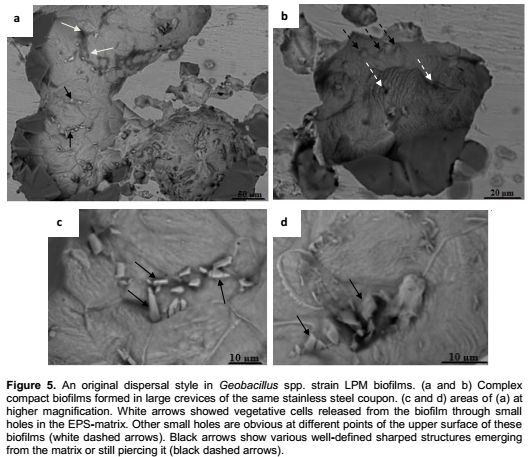
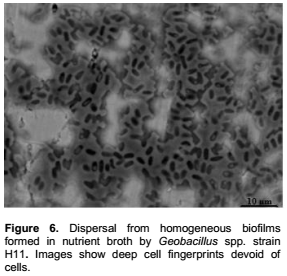
Geobacillus spp. strains developed densely packed mature biofilms undergoing an original dispersing style as well as extensive spread structures. Figure 5a showed large heterogeneous biofilms developed in crevices. Central parts of this biofilm were surrounded with large pieces of homogeneous EPS-matrix. Various geometrically shaped structures, better visualized at higher resolution (Figure 5c and d), were scattered at the upper surface of the biofilm. In this heterogeneous and closed structure, vegetative rod-shaped cells were released through small holes. At the upper surface of another part of this biofilms we can also see the EPS-matrix elevated by some hidden items in a piercing process (Figure 5b). This procedure is clearly understood, in Figure 5c and d. The emerging materials are slightly sharp tools, which pierced the EPS matrix, enabling cells to escape, from the biofilm. Figure 6 shows more conventional dispersion styles resulting from matrix degrading or through the formation of central voids in extensive structures (Figure 7) as well as detachment of small portions from a heterogeneous octopus-like biofilm (Figure 8).


Non-submerged mixed-species biofilm
Mixed-species biofilms formed by a mixture of B. cereus strain BC15 and Geobacillus spp. strain HP were heterogeneous mosaics (Figure 9), invoking the structure of the biofilm formed individually by B. cereus strain BC15 (Figure 2b). It seems that, in co-culture at 42°C, B. cereus was best able to grow in biofilm than Geobacillus spp.

Biofilms in situ inside milking pipes
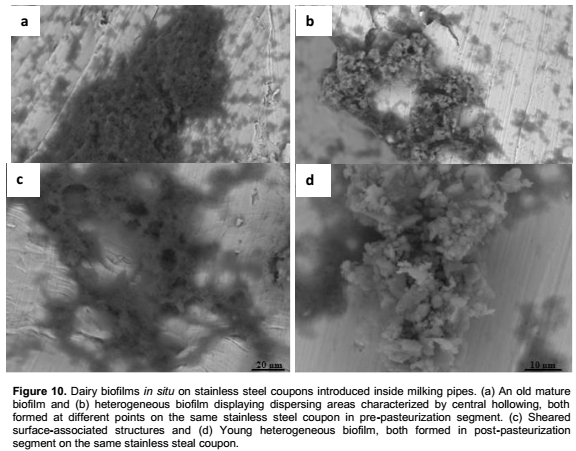
The biofilms formed on suspended stainless steel coupons installed during a week, in lines carrying pasteurized milk are extensive structures characterized by amorphous matrix, and high surface coverage (Figure 10). In post-pasteurization segments these surface-associated structures seem to be more influenced by the flux effect than in pre-pasteurization segments. Younger biofilms developed in both segments probably between two production runs (Figure 10b and d).
Another highly distinct biofilm structure formed on stainless steel coupons installed at another site of the pre-pasteurization section is shown in Figure 11b and c. Straight or curved rod-shaped matrix characterized these biofilms, which strikingly resemble non-submerged in vitro biofilms formed by strains of Geobacillus spp. in nutrient broth (Figure 11a). It seems that all these structures, probably formed under unsuitable conditions may be devoid of living cells. The similarities recorded for biofilms formed in situ and those formed in vitro, highlights the usefulness of the non-submerged assay for the study of biofilms.

DISCUSSION
This study proposes a method that relies on the use of environmental scanning electron microscopy combined with a simple cultivation, technique which has proved to be compatible with high resolution imaging, for the study of biofilm structures. ESEM provided excellent resolution of dairy biofilms formed in vitro and in situ inside industrial milking pipes, and revealed structures not previously described for B. cereus or Geobacillus spp. biofilms. It is noticeable that ESEM images of B. cereus non-submerged biofilms are, at some extent, comparable to those obtained in SEM or CLSM for immersed biofilms formed by other bacteria (Gulot et al., 2002; Parsek and Fuqua, 2004), highlighting the usefulness of this model for the study of biofilms.
Results showed that both B. cereus and Geobacillus spp. biofilms underwent rapid dispersion within 20 h or lesser, in comparison with previous work (Wijman et al., 2007). This indicates that this biofilm non-submerged model supported a luxurious growth resulting in rapid dispersal. In addition, dispersion occurred in the way called seeding dispersal or central hollowing, previously described for Pseudomonas aeruginosa biofilms cultured in flowing systems (Boles et al., 2005; Kirov et al., 2007). Indeed, non-submerged biofilms developed under static conditions were substantial structures with high colony diameters of > 80 µm, previously reported as a threshold required for hollow cavity formation to occur in P. aeruginosa biofilms (Purevdorj-Gage et al., 2005). The structural similarities recorded for biofilms formed in batch system and biofilms in dynamic systems increases the score of the non-submerged assay as an efficient biofilm cultivation method. Interestingly, central hollowing was also found in complex biofilms developed in dairy processing lines. Other conventional dispersal processes observed in non-submerged biofilms, included sloughing, erosion and cell release from mosaic or uniform matrix biofilms.
Non-submerged biofilms formed by strains of Geobacillus spp. displayed an original dispersion style not previously described. Cells were released through small holes performed in the matrix using sharped tools. This strategy to leave the biofilms was also observed in B. cereus compact mushroom-like biofilm. Unlike recognized dispersing mechanisms, this new strategy to escape from the biofilm appears not to rely upon biochemical matrix degrading, but, on a physical process which consisted of piercing the matrix surface using well-defined geometrical sharped structures most likely of crystalline nature. Crystal structures in biofilms have mainly been ascribed to mineral formations in specific biofilms characterized by high rates of minerals (Holling et al., 2014). Such high content in minerals has neither been reported for B. cereus matrix biofilms, mainly composed of polysaccharides, proteins and eDNA (Vilain et al., 2009), nor for thermophilic bacilli biofilms. Thereby, the observed piercing tools should rather be organic formations mainly polysaccharides or/and proteins.
Detachment or dispersion is the final stage of biofilm development and an essential stage of the biofilm cycle life. This process has a crucial meaning with regard to cross contamination and disease transmission. That is why, in recent years, dispersion was considered as an interesting target for biofilm prevention and control strategies, in industrial and clinic settings (Kaplan, 2010; McDougald et al., 2011, Rabin et al., 2015). Interestingly, non-submerged biofilms combined to ESEM appears as an efficient model for analysis of biofilm dispersion and should provide additional data and measurements, since samples examined in ESEM can be used with a range of downstream methods directly after viewing (Bergmans et al., 2005). This will permit in-depth investigations of biofilm dispersal notably the identification of the piercing tools.
Major differences in biofilm structures were not only strain dependent but also related to environment factors namely culture media and adhesion sites on the stainless steel coupons. The non-submerged assay successfully illustrated this wide structural diversity in biofilms formed by sporeforming bacteria, and produced elaborate three-dimensional or more extensive structures often charac-terized by complex networks of pores or water channels. ESEM pictures confirmed recent data concerning biofilm formation in mesophilic and thermophilic sporeforming bacteria (Gopal et al., 2015; Majed et al., 2016). Bacillus biofilms were previously shown to contain both spores and vegetative populations (Lindsay et al., 2006; Pagedar and Singh, 2012). In addition, sporulation and biofilm formation have been shown to occur simultaneously in the thermophilic bacilli Anoxybacillus flavithermus (Burgess et al., 2010). In the current study, spores of B. cereus appeared more present in matrix-rich biofilms than in luxurious biofilms. Furthermore, in the biofilm formed by the spore deficient B. cereus strain BC3 (Figure 3), the amount of EPS is obviously lesser than in the obtained spore-induced biofilms. In good agreement, Simões et al. (2007) found small amount of EPS in the luxurious biofilm formed by B. cereus. Interestingly, these findings confirmed data concerning B. cereus biofilms developed in dynamic system, and provide additional evidence for the efficiency of the combination of non-submerged assay to ESEM as a model for biofilm analysis.
At the morphological level, ESEM imaging showed that, non-submerged biofilms often displayed homogeneous matrix, previously reported to be less stable compared to their heterogeneous counterparts (Anand et al., 2014), enhancing the risk of cell dissemination into processing environments. These findings are interesting cues for the dairy industry.
In good agreement with previous work (Austin and Bergeron, 1995), dairy biofilms formed inside milk processing lines, were mostly extensive structures characterized by amorphous matrix. The biofilm developed in pre-pasteurization sections, should result from a robust bacterial growth due to heavy contamination of milking pipe surfaces. Bacterial contamination of the process equipment in this dairy plant was found to occur at high levels (up to 107 cfu/cm2) (unpublished data) and such cell density was reported to result in biofilms structures consisting of several layers (Gibson et al., 1999).
Therefore, dairy biofilms are characterized by the predominance of single species of bacteria, due to selective pressure from surrounding environments (Flint et al., 1997). In the investigated dairy plant, mesophilic and thermophilic sporeformers were predominant groups of constitutive bacteria of the biofilms developed on milk processing line surfaces. Strikingly, certain biofilms in situ displayed similar structures to those developed in vitro by B. cereus or Geobacillus spp. and should be attributed to the development of one or the other but not to both organisms. Indeed, the non-submerged mixed-species biofilms consisted of widely distinct structures imaged as heterogeneous mosaic biofilms (Figure 9).
Overall, the biofilm system used in situ provided valuable data concerning the hygienic statute of the investigated dairy plant indicating the failure of conventional cleaning procedures to remove spores and biofilms. The development of young biofilms on post-pasteurization segments, after CIP procedures as well as detachment of small portions from older biofilms, highlights high cross contamination risk. In conclusion, the microorganism carrier-surface method combined to ESEM revealed a relevant model for the study of dairy biofilms. The cultivation of biofilms on non-immersed surfaces produced a diversity of B. cereus and Geobacillus spp. biofilm architectures and specific dispersion features. This model is rapid, easy to control and suitable for routine use. Moreover, considering its efficiency for testing the effectiveness of sanitation agents, this model is of major concern to the dairy industry. Non-submerged biofilms produced under static conditions were also comparable to those developed in dynamic systems as well as to biofilms in situ in dairy processing lines. This clearly suggests that data of the colonization of open surfaces should be extrapolated to closed piping systems, and overall, highlights the relevance of this approach for the study of biofilms.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Alhede Alhede M, Qvortrup K, Liebrechts R, Høiby N, Givskov M, Bjarnsholt T (2012). Combination of microscopic techniques reveals a comprehensive visual impression of biofilm structure and composition. FEMS Immunol. Med. Microbiol. 65:335-342. |
|
|
Anand S, Singh D, Avadhanula M, Marka S (2014). Development and Control of Bacterial Biofilms on Dairy Processing Membranes. Comprehensive Reviews in Food Science and Food Safety 13:18-33. |
|
|
Austin JW, Bergeron G (1995). Development of bacterial biofilms in dairy processing lines. J. dairy res. 62:509-519. |
|
|
Bénézech T, Lelièvre C, Membré JM, Viet AF, Faille C (2002). A new test method for in-place cleanability of food processing equipment. J. Food Eng. 54:7-15. |
|
|
Bergmans L, Moisiadis P, van Meerbeek B, Quirynen M, Lambrechts P (2005). Microscopic observation of bacteria: review highlighting the use of environmental SEM. Int endod J. 38:775-788. |
|
|
Boles BR, Thoendel M, Singh PK (2005). Rhamnolipids mediate detachment of Pseudomonas aeruginosa from biofilms. Mol. Microbiol. 57:1210-1223. |
|
|
Bremer PJ, Fillery S, McQuillan AJ (2006). Laboratory scale Clean-In Place (CIP) studies on the effectiveness of different caustic and acid wash steps on the removal of dairy biofilms. Int. J. Food Microbiol. 106:254-262. |
|
|
Burgess SA, Lindsay D, Flint SH (2010). Thermophilic bacilli and their importance in dairy processing. International J. food microbial. 144:215-225. |
|
|
Faille C, Bénézech T, Blel W, Ronse A, Ronse G, Clarisse M, Slomianny C (2013). Role of mechanical vs. chemical action in the removal of adherent Bacillus spores during CIP procedures. Food Microbiol. 33:149-157. |
|
|
Flint SH, Bremer PJ, Brooks JD (1997). Biofilms in dairy manufacturing plant-description, current concerns and methods of control. Biofouling 11:81-97. |
|
|
Gibson H, Taylor JH, Hall KE, Holah JT (1999). Effectiveness of cleaning techniques used in the food industry in term of removal of bacterial biofilms. J. Appl. Microbial. 87:41-48. |
|
|
Gopal N, Hill C, Ross P, Beresford TP, Fenelon MA, Cotter PD (2015). The prevalence and control of Bacillus cereus and related spore-forming bacteria in the dairy industry. Front Microbiol. 6:14-18. |
|
|
Guinebretière MH, Thompson FL, Sorokin A, Normand P, Dawyndt P, Ehling-schulz M, Svensson B, Sanchis V, Nguyen-The C, Heyndrickx M, De Vos P (2008). Ecological diversification in the Bacillus cereus group. Environ. Microbiol.10:851-865. |
|
|
Gulot E, Georges P, Brun A, Fontaine-Aupart MP, Bellon-Fontaine MN, Briandet R (2002). Heterogeneity of diffusion inside microbial biofilms determined by fluorescence correlation spectroscopy under two-photon excitation. Photochem. photobiol. 75:570-578. |
|
|
Holling N, Dedi C, Jones CE, Howthorne JA, Hanlon GW, Salvage JP, Patel BA, Barnes LM, Jones BV (2014). Evaluation of environmental scanning electron microscopy for analysis of Proteus mirabilis biofilms in situ on urinary catheters. FEMS Microbiol. Lett. 355(1):20-27. |
|
|
Kaplan JB (2010). Biofilm Dispersal: Mechanisms, Clinical Implications, and Potential Therapeutic Uses. J. Dent. Res. 89:205-218. |
|
|
Kirov SM, Webb JS, O'May CY, Reid DW, Woo JKK, Rice SA, Kjelleberg S (2007). Biofilm differentiation and dispersal in mucoid Pseudomonas aeruginosa isolates from patients with cystic fibrosis. Microbiology 153:3264-3274. |
|
|
Kumari S, Sarkar PK (2014). In vitro model study for biofilm formation by Bacillus cereus in dairy chilling tanks and optimization of clean-in-place (CIP) regimes using response surface methodology. Food Control 36:1153-1158. |
|
|
Leriche V, Carpentier B (2000). Limitation of adhesion and growth of Listeria monocytogenes on stainless steel surfaces by Staphylococcus sciuri biofilms. J. Appl. Microbiol. 88:594-605. |
|
|
Lindsay D, Brozel VS, Von Holy A (2006). Biofilm-spore response in Bacillus cereus and Bacillus subtilis during nutrient limitation. J. Food Prot. 69:1168-1172. |
|
|
Little B, Wagner P, Ray R, Pope R, Scheetz R (1991). Biofilms: an ESEM evaluation of artifacts during SEM preparation. Journal of industrial microbiology 8:213-222. |
|
|
Majed R, Faille C, Kallassy M, Gohar M (2016). Bacillus cereus Biofilms—Same, Only Different. Front. Microbiol. 7: 1054. |
|
|
Malek F, Moussa-Boudjemaa B, Aouar-Metri A, Mabrouk K (2013). Identification and genetic diversity of Bacillus cereus strains isolated from a pasteurized milk processing line in Algeria. Dairy Sci. technol. 93:73-82. |
|
|
Marchand SJ, De Block VE, De Jonghe A, Coorevits M, Heyndrick X, Herman L (2012). Biofilm Formation in Milk Production and Processing Environments; Influence on Milk Quality and Safety Comprehensive. Rev. Food Sci. Food Safety 11:133-147. |
|
|
Maris P (1992). Biofilm and disinfection of microorganism carrier-surface method. Sci. des aliments 12:721-728. |
|
|
McDougald D, Rice SA, Barraud N, Steinberg PD, Kjelleberg S (2011). Should we stay or should we go: mechanisms and ecological consequences for biofilm dispersal. Nature Review Microbiology 10:39-50. |
|
|
Pagedar A, Singh J (2012). Influence of physiological cell stages on biofilm formation by Bacillus cereus of dairy origin. Int. dairy J. 23:30-35. |
|
|
Parkar SG, Flint SH, Brooks JD (2004). Evaluation of the effect of cleaning regimes on biofilms of thermophilic bacilli on stainless steel. J. Appl. microbiol. 96:110-116. |
|
|
Parsek MR, Fuqua C (2004). Biofilms 2003: Emerging themes and challenges in studies of surface-associated microbial life. J. bacterial. 186:4427-4440. |
|
|
Peng JS, Tsai WC, Chou CC (2001). Surface characteristics of Bacillus cereus and its adhesion to stainless steel. Int. J. food microbial. 65:105-111. |
|
|
Purevdorj-Gage B, Costerton WJ, Stoodley P (2005). Phenotypic differentiation and seeding dispersal in non-mucoid Pseudomonas aeruginosa biofilms. Microbiology 151:1569-1576. |
|
|
Rabin N, Zheng Y, Opoku-Temeng C, Du Y, Bonsu E, Sintim Ho (2015). Agents that inhibit bacterial biofilm formation. Future Med. chem. 5: 647-671. |
|
|
Simmonds P, Mossel BL, Intaraphan T, Deeth HC (2003). Heat resistance of Bacillus spores when adhered to stainless steel and its relationship to spore hydrophobicity. J food protect. 66:2070-2075. |
|
|
Simões M, Cleto S, Pereira MO, Vieira MJ (2007). Influence of biofilm composition on the resistance to detachment. Water Sci. Technol. 55:473-80. |
|
|
Sommer P, Martin-Rouas C, Mettler E (1999). Influence of the adherent population level on biofilm population, structure and resistance to chlorination. Food Microbiol. 16:503-515. |
|
|
Vilain S, Pretorius JM, Theron J, Brözel VS (2009). DNA as an Adhesin: Bacillus cereus Requires Extracellular DNA To Form Biofilms. Appl. Environ. Microbiol. 75:2861-2868. |
|
|
Wijman JGE, de Leeuw PPLA, Moezelaar R, Zwietering MH, Abee T (2007). Air liquid interface biofilms of Bacillus cereus: Formation, Sporulation and Dispersion. Appl. Environ. Microbiol. 73:1481-1488. |
|
|
Zhao Y, Caspers MPM, Metselaar KI, de Boer P, Roeselers G, Moezelaar R, Groot MN, Montijn RC, Abee T, Korta R (2013). Abiotic and microbiotic factors controlling biofilm formation by thermophilic sporeformers. Appl. Environ. Microbiol. 79:5652-5660. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


