ABSTRACT
Bacteriophages (phages) are viruses that infect bacteria. The emergence of resistant bacteria has been reported worldwide. The use of phages to treat bacterial infection or to reduce bacteria load has become a new area of interest. The viral colonization of rodents represents an important source for a specific biotope. The objective of this study is to characterize the effect of novel phages isolated from rodents (Mastomys) using clinical Enterobacteria strains. Three novel phages were isolated from the intestinal tract of rodents. The phages belong to Myoviridae family. By using three Random Polymerase Chain Reactions (PCR), genomic characterization of the phages shows a high diversity that reveals several DNA bands. 3 virulent phages exhibited different patterns, indicating that the rodents’ phages are genetically unique and could be distinguished by typing-PCR. The virulence spectrum of the phages shows phages Mα3 and Mα7 have 50% (7/15) lytic activity on enteropathogenic and enterohemorrhagic Escherichia coli strains while phage Mα11 has 26.6% (4/15) lytic activity. These phages have no lytic activity on Salmonella strains. This study is the first to isolate novel phages from rodents with lytic activity. The candidate rodents’ phages (Mα3, Mα7, Mα11) can be used to control virulent E. coli in West Africa.
Keys words: Lytic phages, rodents, Escherichia coli strains, West Africa, enteropathogenic, enterohemorrhagic.
Bacterial infections are mostly involved in causing diseases in sub-Saharan countries. Goualié et al. (2019) reported the high incidence of multidrug resistant bacteria in chickens in Cote d’Ivoire. This problem is emergent because of the misuse of drugs and the free accessibility to drugs in human or in animal trials. Several studies have reported the distribution of multi-drug resistant strains isolated from humans, animals and the environment (Guessennd et al., 2013). Fagmabila et al. (2017) reported the distribution of Salmonella serovars in
commercial poultry farms in West Africa. Bacteriophages, the most ubiquitous organisms on Earth, are viruses that infect bacteria (Rohwer, 2003). These have been employed since their discovery in the development of therapeutics against infections. They are highly specific, very safe, and effective against their target pathogenic bacteria; they are easily modifiable to address new threats (Golkar et al., 2014). Regardless of the increase in antibiotics resistance worldwide, the use of phages is an alternative for the treatment of bacterial infections (Nagel et al., 2016). Many studies have reported the efficacy of phage therapy in humans, animals and the environment (Biswas et al., 2002; Wang et al., 2006; Watanabe et al., 2007; Wright et al., 2009; Abedon et al., 2011; Nale et al., 2016; Lin et al., 2017).
Phages were discovered before the development of antibiotics and their administration as pharmaceutical agents was a common practice in the pre-antibiotic era (Golkar et al., 2014). On the contrary, phage therapy is not known in sub-Saharan countries’ medical care systems (Ehui et al., 2017). Some studies have reported the abundance and isolation of lytic phages in West Africa (Essoh et al., 2015; Kakou-Ngazoa et al., 2017; Ngazoa-Kakou et al., 2018, 2019), but the applications of phage therapy are not reported. Nagel et al. (2016) suggested the need to implement phage therapy in low-income countries because it is cheap to produce and is available everywhere in rural regions. Because of the renewed interest in lytic phages as biocontrol agents, new phages are sought after as they offer the possibility of increasing bacterial strain coverage in the design and development of phage cocktails (Golkar et al., 2014). Most of the studies on phages focus on Enterobacteria phages. Enterobacteria colonize major biotopes, intestine, sewage, water bodies (Kumari et al., 2009; Park et al., 2012; Maal et al., 2015; Hamdi et al., 2017) and are abundant in sub-tropical countries (Essoh et al., 2015). Rodents are an important reservoirs of pathogens involved in zoonosis. Lozano et al. (2015) isolated multi-drug resistant Enterococcus faecalis strains in Rattus. To propose that phages are effective against several bacteria strains, we investigated the phages that infect the Enterobacteriaceae family. The main aim of this study is to evaluate the lytic activity of novel phages isolated from rodents on clinical virulent bacteria strains.
Isolation of rodent phages
Fifty samples of rodent (Mastomys) intestines were obtained from Pasteur Institute Project for the Surveillance of Zoonosis in Côte d’Ivoire (West Africa). 1-2 cm of the intestines was incubated in 1 ml PBS buffer (1X) and mixed hard for 15 min. For the amplification of the phages, the solution was passed through a filter of 0.45 µm. One ml of the filtrate was added to 3ml Luria Bethani broth containing 100 µl of bacteria host (Escherichia coli). The solution was incubated at 37°C overnight and centrifuged at 8000 rpm, for 10 min. The supernatant was filtered at 0.45 µm and the filtrate was incubated in 3 ml LB broth containing bacteria host (E. coli). The step was repeated twice for high viral load (Maal et al., 2015). Chloroform or 1% NaN3 was added to the bacteria in filtrate samples. The isolated phages were named as Mα3, Mα7 and Mα11 and stored at 4°C.
Morphology identification
Twenty five microliters of each concentrated bacteriophage suspended (minimum 107 PFU/ml) in SM buffer was spotted on top of a Formvar-carbon-coated copper grid (Ted Pella, Inc, USA). The bacteriophages were allowed to adsorb for 2 min and were stained by adding 8 µl of 2% sodium phosphotungstate (pH 7.6) (Kumari et al., 2009). The grids were observed with a Zeiss EM 900 transmission electron microscope (TEM) at 80 kV.
Virulence of phages on bacteria strains
E. coli and Salmonella strains were obtained from Institut Armand Frappier, Microbiology Unit (Laval, Canada), with some virulence factors. The bacteria were incubated in LB broth overnight at 37°C. Fresh broths of bacteria were inoculated on fresh LB-Agar. Rodents’ phages MαΑ3, MαΑ7 were propagated on E. coli DH5α and rodents’ phage Mα11 on E. coli QT22 at 37°C, for 1 h (Maal et al., 2015).
Lytic activity of phages
5-10 µl of the phage solution was inoculated on LB-Agar (0.5%) containing fresh bacteria host by OD <0.3. The plate was incubated at 37°C, overnight. The plates were sectioned in 24 quadrats. The lytic activity of the phages was evaluated by the formation of clear plaques. The bacterial strains used were provided from the Biocollection of Institut Armand Frappier, Quebec, Canada (Microbiology Unit, Dr Dozois C.).
Phage DNA Isolation
8.6 µl of the phage solution was added to DNase-mix containing Dnase I buffer 1X, 20 Unit of DNaseI (Thermo Fisher, USA) to obtain a final volume of 10 µl. The solution was incubated at 37°C and 65°C, for 30 min. After the digestion, 0.5 µl of Proteinase K (20 mg/ml) was added and incubated at 37°C for 30 min and 95°C for 15 min. The DNA was stored at -20°C.
Random PCR
Three random primer PCRs were used for the genomic analysis of the isolated phages, according to the protocol described by Gutierrez et al. (2011) and Comeau et al. (2004). Briefly, 2 µl of the phages’ DNA were added to 23 µl PCR Mix containing 12.5 µl Taq-Frogga Mix, 2 µl of 100 µM DEG Primer (R10D) or P2, or OPL5 and 8.5 µl Dnase-free water. The PCR conditions for R10D were: 95°C, 5 min; 95°C, 45 s; 40°C, 3 min; 72°C, 1 min (40 cycles); and 72°C, 10 min. The PCR conditions for Primers P2 and OPL5 were 95°C, 2 min; 95°C, 45 s; 30°C, 2 min; 72°C, 1 min (40 cycles), 26 cycles at 94°C, 45 s; 36°C, 30 s; 72°C, 60 s; and final extension of 10 min at 72°C. PCR products were separated in 1% agarose gel and the TIF image was created from UV Gel Doc system (Biorad, USA).
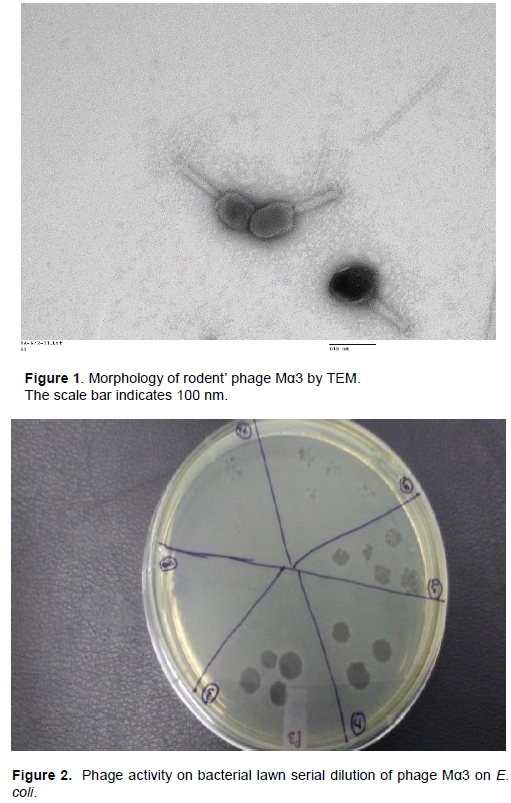
Morphology of rodents’ phages by electronic microscopy
The transmission electron microscopy (TEM) classified the morphology of the virions of Mα3, Mα7 and Mα11 as Myoviridae family. The rodents’ phages have elongated head (195 nm) and contractile tail (110-114 nm) (Figure 1).
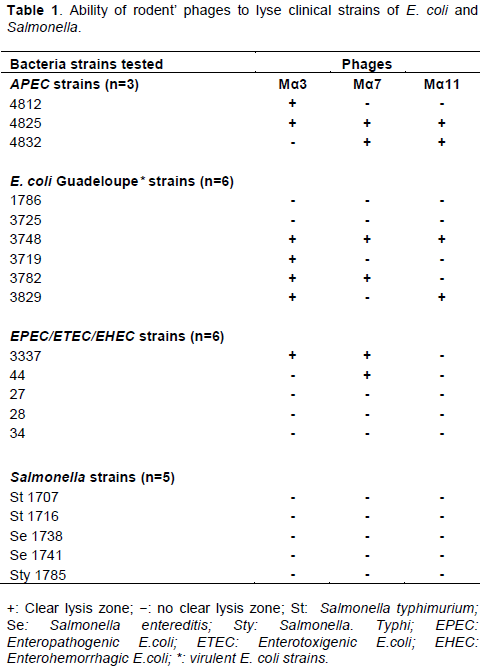
Host range of rodents’ phages on clinical strains
The host range of rodents’ phages Mα3, Mα7 and Mα11 was tested on 20 bacterial strains by spot test of diluted phage lysate. Among the 20 strains tested, phages Mα3 and Mα7 were able to infect 7 bacterial strains while phage Mα11 infected 4 strains (Table 1). The virulence spectrum shows that phages Mα3 and Mα7 have 50% (7/15) lytic activity against E. coli strains and phage Mα11 has 26.6% (4/15) lytic activity. Out of 6 EPEC/ETEC/EHEC strains, rodent phages Mα3 and Mα7 were able to lyse 4 strains (Table 1 and Figure 2). Among three APEC strains, two were sensitive to the isolated rodent phages. These phages have no lytic activity against Salmonella strains (Table 1).
Rodents’ phages typing by PCR
Random Amplified Polymorphism DNA (RAPD)-bases methods were used to establish a quick typing of the isolated phages. The results show the diversity of the isolated phages in 3 typing PCRs. With R10D primer, 3 different profiles were obtained. M3α reveals 3 bands (0.4, 0.6, 1.1 kb); 7 bands for Mα7 and 3 bands ranging from 0.2-1.100 kb for Mα11 (Figure 3). By using PCR for P2 Primer, the phages generated DNA band patterns with amplicons size ranging from 0.7-0.8 kb. Phage Mα11 shows two bands of amplification (Figure 3). The PCR used for OPL5 primer reveals a profile of 3 bands, 2 bands and one band for M3α, M7α and Mα11 respectively (Figure 3). Each phage showed a different profile regardless of the generated bands by RAPD-PCRs. All phages exhibited different patterns showing that all phages are genetically unique and could be distinguished by RAPD-PCR.
In this study, we isolated and characterized three virulent phages Mα3, Mα7 and Mα11 from intestinal tract of Mastomys. Our study reveals a new biotope for phage isolation. Several studies have isolated most Enterobacteriaceae phages from sewages, lakes and aquatic sources (Essoh et al., 2015; Maal et al., 2015; Hamdi et al., 2017). Morphological and genomic analyses revealed that all phages belong to the Myoviridae. The isolated phages have broad host ranges and were coliphages. Other studies have shown similar results with broad host range from phages isolated in sewage (Hamdi et al., 2017). Jothikumar et al. (2000) have first reported ten phages specific to Enterotoxigenic E. coli (ETEC), and this study has revealed two phages Mα3, Mα7 against ETEC strains and all tree phages Mα3, Mα7, Mα11 against APEC strains.
Several studies have reported coliphages belong to Myoviridae and Siphoviridae families. The extended host range of phages correlates with the acquisition of new fiber genes. In this study, we cannot demonstrate the phylogeny of the rodent phages, because of the lack of NGS analysis. The genomic diversity of phages by using RAPD-PCR revealed the existence of different clusters for the rodent’s phages. Previous studies have similar results to differentiate different phages families isolated from various sources by RAPD-PCR (Comeau et al., 2004; Gutierrez et al., 2011; Jothikumar et al., 2000). The lytic rodent’s phages, individually or a cocktail, may be useful for reducing contamination in food processing, or to control bacterial infections in humans. Also, our results suggest the exploration of new microbiomes from rodents to identify phages of interest in biotechnological applications and the perspectives for genomics tools.
This study describes the first isolation of rodent’s phages with lytic activity on clinical virulent bacterial strains. The rodent’s phages Mα3, Mα7 and Mα11 have E. coli as host bacteria, suggesting they belong to the family of coliphages. 96% of coliphages belong to Caudavirales. Most isolated phages were in the family of Enterobacteriaceae phages. Genomic typing by RAPD shows the divergence of isolated rodent’s phages. The effect of rodent’s phages on hemorrhagic strains was to reduce bacterial infection. The candidate rodent phages can be applied in phage therapy in West Africa to fight multi-drug strains bacteria. Analysis of morphology and Next Generation Sequencing (NGS) method should be done in the future.
The authors have not declared any conflict of interests.
The authors thank the National Surveillance of Zoonosis of Pasteur Institute, Cote d’Ivoire for the collaboration; The Government of Togo for the grant of Master 2 for Addablay Audrey, Dr Imziln B. from The University of Marrakech, Morocco; and The Institut Armand Frappier, Québec, Canada for the bacterial strains tested in this study.
REFERENCES
|
Abedon ST, Thomas-Abedon C, Thomas A, Mazure H (2011). Bacteriophage prehistory. Bacteriophage 3:174-178.
Crossref
|
|
|
|
Biswas B, Adhya S, Washart P, Paul B, Trostel AN, Powell B, Carlton R, Merril CR (2002). Bacteriophage therapy rescues mice bacteremic from a clinical isolate of vancomycin-resistant Enterococcus faecium. Infection and immunity 70(1):204-210.
Crossref
|
|
|
|
|
Comeau AM, Short S, Suttle CA (2004). The use of degenerate primed random amplification of polymorphic DNA (DPRAPD) for strain-typing and inferring the genetic similarity among closely related viruses. Journal of Virological Methods 118(2):95-100.
Crossref
|
|
|
|
|
Ehui PJ, Sekongo KW, Kouassi FK (2017). L'adhésion du personnel soignant ivoirien a la phagotherapie. International Journal of Law Education and Social Special Studies 4:1-10.
View
|
|
|
|
|
Essoh C, Latino L, Midoux C, BlouinY, Loukou G, Nguetta SPA, Lathro S, Cablanmian A, Kouassi AK, Vergnaud G, Pourcel C. (2015). Investigation of a Large Collection of Pseudomonas aeruginosa Bacteriophages Collected from a Single Environmental Source in Abidjan, Côte d'Ivoire. PLoS ONE 10(6):e0130548.
Crossref
|
|
|
|
|
Fagmabila O, Barco L, Mancin M, Kwaga J, Ngulukun SS, Zavagnin P, Lettini AA, Lorenzetto M, Abdu PA, Kabir J, Umoh J, Ricci A, Muhammad M (2017). Salmonella serovars and their distribution in Nigerian commercial chicken layer farms. PLoS ONE 0173097-3-15.
Crossref
|
|
|
|
|
Golkar Z, Bagasra O, Pace DG (2014). Bacteriophage therapy: A potential solution for the antibiotic resistance crisis. Journal of Infection in Developing Countries 8(02):129-136.
Crossref
|
|
|
|
|
Goualié BG, Akpa EE, Kakou-Ngazoa SE, 0uattara HG, Niamke S, Dosso M (2019). Antimicrobial resistance and virulence associated genes in Campylobacter jejuni isolated from chicken in Côte d'Ivoire. The Journal of Infection in Developing Countries 13(08):671-677.
Crossref
|
|
|
|
|
Guessennd NK, Ouattara MB, Ouattara ND, Nevry RK, Gbonon V, Tiekoura KB, Dosso M (2013). Étude des bactéries multirésistantes des effluents hospitaliers d'un centre hospitalier et universitaire (CHU) de la ville d'Abidjan (Côte d'Ivoire). Journal of Applied Biosciences 69:5456-5464.
Crossref
|
|
|
|
|
Gutierrez D, Martın-Platero AM, Rodrıguez A, Martınez-Bueno M, Garcıa P, Martinez B (2011). Typing of bacteriophages by randomly ampli¢ed polymorphic DNA (RAPD)-PCR to assess genetic diversity. FEMS Microbiology Letters 32:90-97.
Crossref
|
|
|
|
|
Hamdi S, Rousseau GM, Labrie SJ, Tremblay DM, Kourda RS, Slama BK, Moineau M (2017). Characterization of two polyvalent phages infecting Enterobacteriaceae. Scientific Reports 7(1):1-12.
Crossref
|
|
|
|
|
Jothikumar N, Reddy CG, Sundari, RB, Saigopal DVR (2000). Isolation of coliphages specific to enterotoxigenic E. Coli (ETEC). Journal of Environmental Monitoring 2(4):372-374.
Crossref
|
|
|
|
|
Kakou-Ngazoa ES, Kouya D, Koudou AA, Abole A, Guessend KN, Sylla A, Sina KM, Coulibaly ND, Kouassi KS, Meite S, Aoussi S, Dosso M (2017). New identification of phages in water and in fishes from Lagoon Ebrié and their application for multidrug strains in Abidjan, West Africa. World Journal of Pharmaceutical Research 6:84-89.
Crossref
|
|
|
|
|
Kumari S, Kusum H, Chhibber S (2009). Characterization of Pseudomonas aeruginosa PAO Specific Bacteriophages Isolated from Sewage Samples. America Journal of Biomedical Science 2:91-102.
Crossref
|
|
|
|
|
Lin DM, Koskella B, Lin HC (2017). Phage therapy: An alternative to antibiotics in the age of multidrug resistance. World Journal of Gastrointestinal Pharmacology and Therapeutics 8(3):162-173.
Crossref
|
|
|
|
|
Lozano C, González-Barrio D, García, JT, Ceballos S, Olea PP, Ruiz-Fons F, Torres C (2015). Detection of vancomycin-resistant Enterococcus faecalis ST6-vanB2 and E. faecium ST915-vanA in faecal samples of wild Mastomys Mastomys in Spain. Veterinary Microbiology 177(1-2):168-174.
Crossref
|
|
|
|
|
Maal KB, Delfan AS, Sharareh S (2015). Isolation and Identification of Two Novel Escherichia coli Bacteriophages and Their Application in Wastewater Treatment and Coliform's Phage Therapy. Jundishapur Journal of Microbiology 8(3):14940-14945.
Crossref
|
|
|
|
|
Nagel TE, Chan BK, De Vos D, El-Shibiny A, Kang'ethe EK, Makumi A, Pirnay J (2016). The Developing World Urgently Needs Phages to Combat Pathogenic Bacteria. Frontiers in Microbiology 7:882-887.
Crossref
|
|
|
|
|
Nale J, Spencer YJ, Hargreaves KR, Buckley AM, Trzepiński P, Douce GR, Clokie MR (2016). Bacteriophage combinations significantly reduce Clostridium difficile growth in vitro and proliferation in vivo. Antimicrobial Agents and Chemotherapy 60(2):968-981.
Crossref
|
|
|
|
|
Ngazoa-Kakou S, Philippe C, Tremblay DM, Loignon S, Koudou A, Abole A, Ngolo CD, Kouassi KS, Kouamé S M, Aoussi S, Dosso M, Moineau S (2018). Complete genome sequence of Ebrios, a novel T7virus isolated from the Ebrie Lagoon in Abidjan, Côte d'Ivoire. Genome Announcements 6(25):e00280-18.
Crossref
|
|
|
|
|
Ngazoa-Kakou S, Shao Y, Rousseau MG, Addablah AA, Tremblay DM, Hutinet G, Lemire N, Plante PL, Corbeil J, Koudou A, Soro BK, Coulibaly ND, Aoussi S, Dosso M, Moineau S (2019). Complete Genome Sequence of Escherichia coli Siphophage BRET. Microbiology Resource Announcements 8(5):e01644-18.
Crossref
|
|
|
|
|
Park M, Lee JH, Shin H, Kim M, Choi J, Kang DH, Heu S, Ryua S (2012). Characterization and Comparative Genomic Analysis of a Novel Bacteriophage, SFP10, Simultaneously inhibiting both Salmonella enterica and Escherichia coli O157:H7. Applied Environmental Microbiology 78:58-69.
Crossref
|
|
|
|
|
Rohwer F (2003). Global phages diversity. Cell 113:141.
Crossref
|
|
|
|
|
Wang J, Hu B, Xu M, Yan Q, Liu S, Zhu X, Sun Z, Tao D, Ding LE, Reed EJ, Gong J, Li QQ, Hu J (2006). Therapeutic effectiveness of bacteriophages in the rescue of mice with extended spectrum betalactamase-producing Escherichia coli bacteremia. International Journal of Molecular Medicine 17(2):347-355.
Crossref
|
|
|
|
|
Watanabe R, Matsumoto T, Sano G, Ishii Y, Tateda K, Sumiyama Y, Uchiyama J, Sakurai S, Matsuzaki S, Imai S, Yamaguchi K (2007). Efficacy of bacteriophage therapy against gut-derived sepsis caused by Pseudomonas aeruginosa in mice. Antimicrobial Agents and Chemotherapy 51:446-452.
Crossref
|
|
|
|
|
Wright A, Hawkins CH, Anggard EE, Harper DR (2009). A controlled clinical trial of a therapeutic bacteriophage preparation in chronic otitis due to antibiotic-resistant Pseudomonas aeruginosa: A preliminary report of efficacy. Clinical Otolaryngology 34(4):349-357.
Crossref
|
|