Full Length Research Paper

ABSTRACT
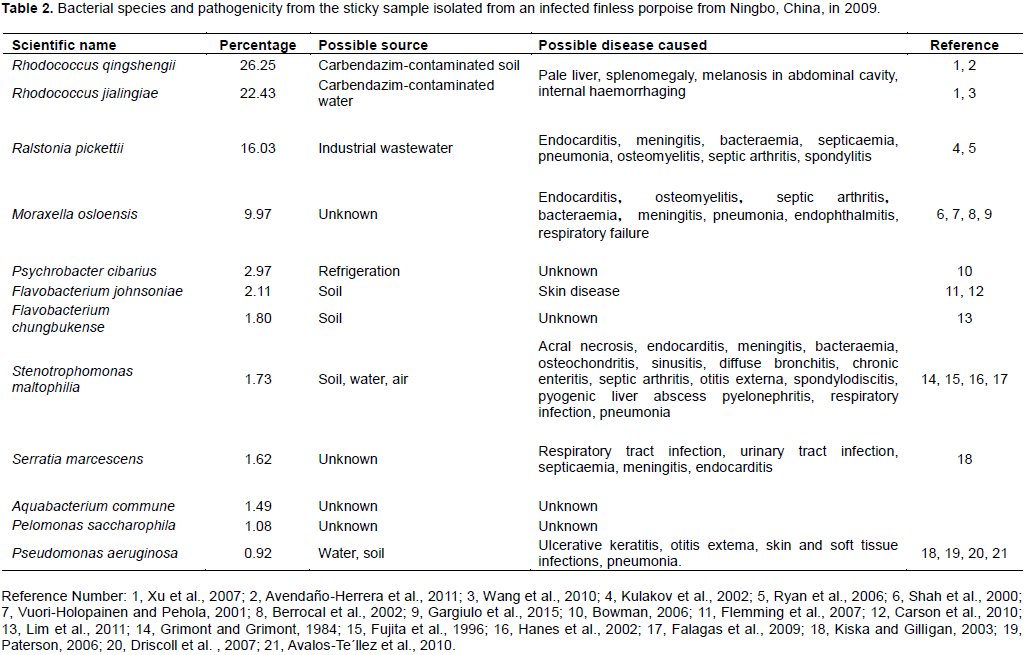
Two East Asian finless porpoises (Neophocaena asiaeorientalis sunameri) in Ningbo, East China Sea, China, were observed to be bacterially infected between the fat layer and muscle layer. The microbial communities in pus samples were identified by metagenome sequencing 16S in the PacBio platform and explored the relationship between emaciation and bacterial infection. The present paper was the first report on bacterial diversity in infected tissue pus of finless porpoise. In total, 101 bacterial species were identified, and the top nine species were Rhodococcus qingshengii (26.25%), Rhodococcus jialingiae (22.43%), Ralstonia pickettii (16.03%), Moraxella osloensis (9.97%), Psychrobacter cibarius (2.97%), Flavobacterium johnsoniae (2.11%), Flavobacterium chungbukense (1.80%), Stenotrophomonas maltophilia (1.73%) and Serratia marcescens (1.62%). These main bacteria could cause various diseases or metabolic disorder, such as endocarditis, meningitis, bacteraemia, septicaemia, and so on; they also involve microbiome metabolism of amino acids (12.47%), carbohydrates (11.58%), xenobiotic biodegradation (7.81%), lipids (6.01%) and energy (4.98%). Regardless of the type of disease metabolic disorder will firstly cause body weight loss. In conclusion, these bacteria could cause diseases or metabolic disorders, resulting in emaciation of East Asian finless porpoises. Admittedly, pathogenic assay is furthermore needed to determine the mechanisms regarding the pathological phenomena.
Key words: Bacterial diversity, East Asian finless porpoises, Metagenome sequencing 16S.
INTRODUCTION
MATERIALS AND METHODS
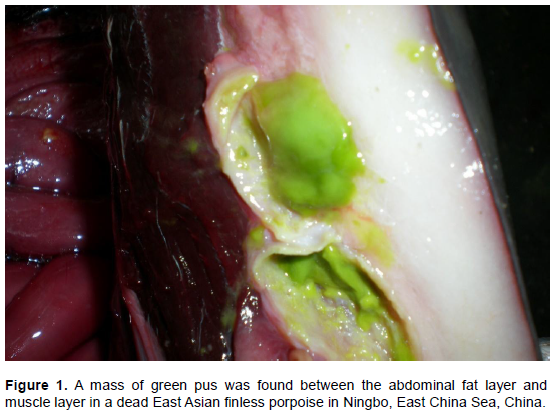
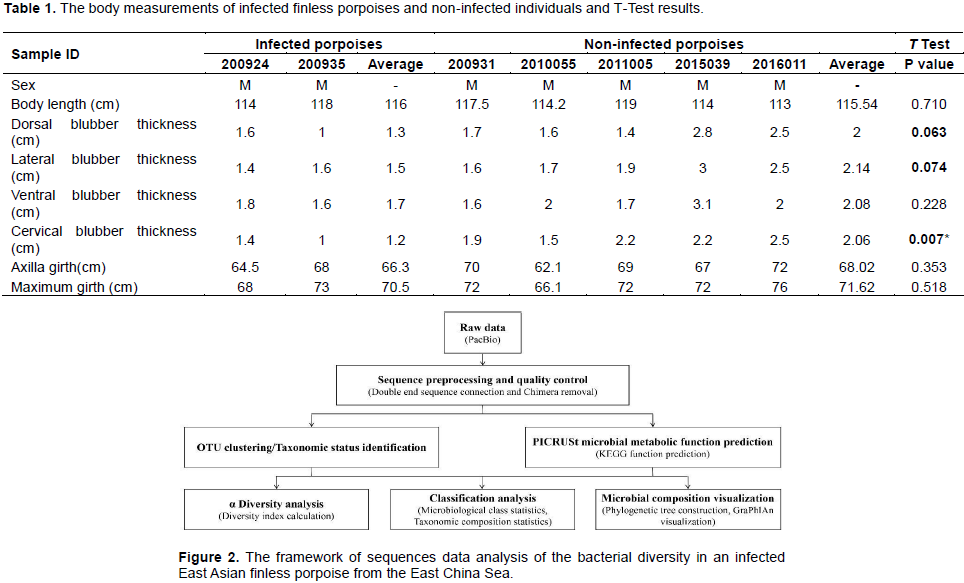
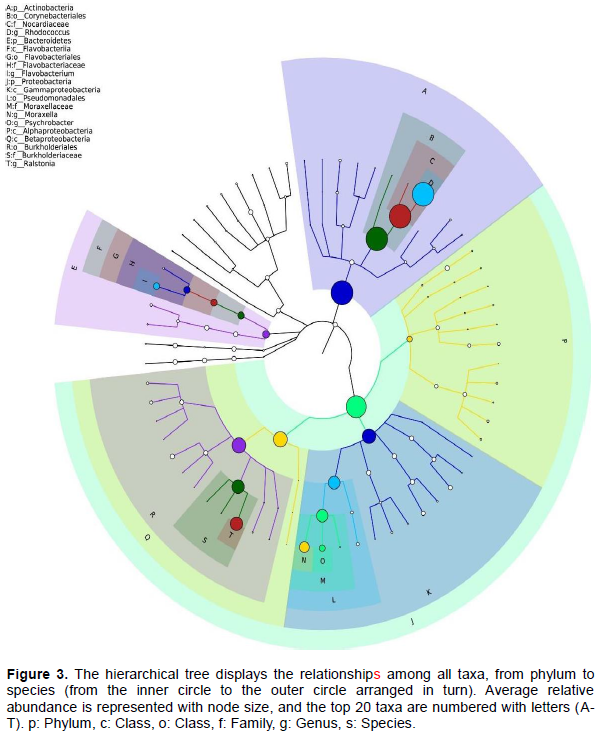
RESULTS
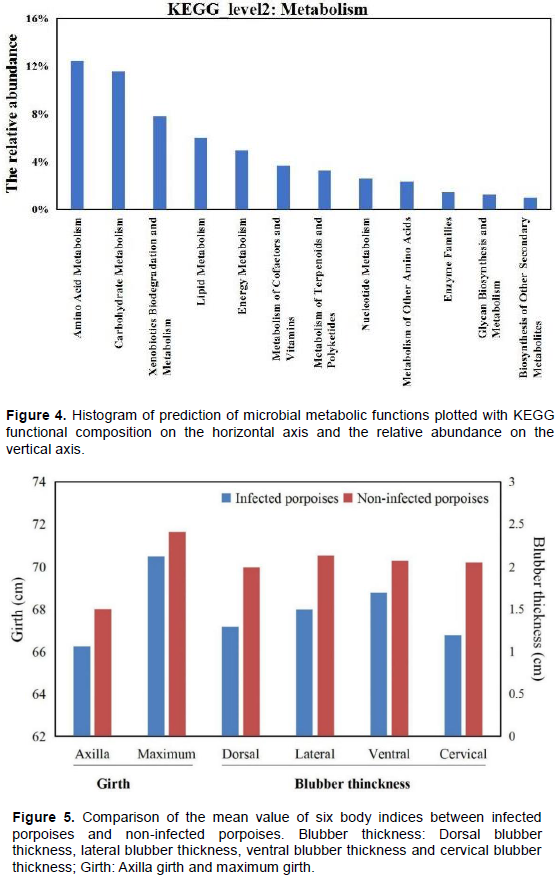
DISCUSSION
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
REFERENCES
|
Amir A, Zeisel A, Zuk O, Elgart M, Stern S, Shamir O, Turnbaugh PJ, Soen Y, Shental N (2013). High-resolution microbial community reconstruction by integrating short reads from multiple 16S rRNA regions. Nucleic Acids Research 41(22):e205. |
|
|
Asnicar F, Weingart G, Tickle TL, Huttenhower C, Segata N (2015). Compact graphical representation of phylogenetic data and metadata with GraPhlAn. Peer Journal 3:e1029. |
|
|
Avalos-Te'llez R, Sua'rez-Gu¨emes F, Carrillo-Casas EM, Herna'ndez-Castro R (2010). Bacteria and yeast normal microbiota from respiratory tract and genital area of bottlenose dolphins (Tursiops truncatus). Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology 2:666-673. |
|
|
Avendaño-Herrera R, Balboa S, Doce A, Ilardi P, Lovera P, Toranzo AE, Romalde JL (2011). Pseudo-membranes on internal organs associated with Rhodococcus qingshengii infection in Atlantic salmon (Salmo salar). Veterinary Microbiology 147:200-204. |
|
|
Bayer T, Neave MJ, Alsheikh-Hussain A, Aranda M, Yum LK, Mincer T, Hughen K, Apprill A, Voolstra CR (2013). The microbiome of the Red Sea coral Stylophora pistillata is dominated by tissue-associated Endozoicomonas bacteria. Applied and Environmental Microbiology 80:4759-4762. |
|
|
Blaxter M, Mann J, Chapman T, Thomas F, Whitton C, Floyd R, Abebe E (2005). Defining operational taxonomic units using DNA barcode data. Philosophical Transactions of the Royal Society 360:1935-1943. |
|
|
Bowman JP (2006). The genus Psychrobacter. In: Dworkin M, Falkow S, Rosenberg E, Schleifer KH, Stackenbrandt E, eds. The Prokaryotes. New York: Springer pp. 920-930. |
|
|
Buck JD, Stephen S (2010). The occurrence of potentially pathogenic vibrios in marine mammals. Marine Mammal Science 2:319-324. |
|
|
Caporaso JG, Kuczynski J, Stombaugh J, Bittinger K, Bushman FD, Costello EK, Fierer N, Peña AG, Goodrich JK, Gordon JI, Huttley GA, Kelley ST, Knights D, Koenig JE, Ley RE, Lozupone CA, McDonald D, Muegge BD, Pirrung M, Reeder J, Sevinsky JR, Turnbaugh PJ, Walters WA, Widmann J, Yatsunenko T, Zaneveld J, Knight R (2010). Qiime allows analysis of high-throughput community sequencing data. Nature Methods 7:335-336. |
|
|
Carson J, Schmidtke LM, Munday BL (2010). Cytophaga johnsonae: a putative skin pathogen of juvenile farmed barramundi, Lates calcarifer Bloch. Journal of Fish Diseases 16:209-218. |
|
|
Chao A (1984). Nonparametric Estimation of the Number of Classes in a Population. Scandinavian Journal of Statistics 11:265-270. |
|
|
Charuvaka A, Rangwala H (2011). Evaluation of short read metagenomic assembly. BMC Genomics 12:S8. |
|
|
Cole JR, Wang Q, Cardenas E, Fish J, Chai B, Farris RJ, Kulam-Syed-Mohideen AS, McGarrell DM, Marsh T, Garrity GM, Tiedje JM (2009). The Ribosomal Database Project: improved alignments and new tools for rRNA analysis. Nucleic Acids Research 37:D141-D145. |
|
|
Dawid AP (1993). Stopping rules and estimation for recapture debugging with unequal failure rates. Biometrika 80:193-201. |
|
|
Desantis TZ, Hugenholtz P, Larsen N, Rojas M, Brodie EL, Keller K, Huber T, Dalevi D, Hu P, Andersen GL (2006). Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Applied and Environmental Microbiology 72:5069-5072. |
|
|
Driscoll JA, Brody SL, Kollef MH (2007). The epidemiology, pathogenesis and treatment of Pseudomonas aeruginosa infections. Drugs 67:351-368. |
|
|
Dunn JL, Buck JD, Robeck TR (2001). Bacterial Diseases of Cetaceans and Pinnipeds. CRC Handbook in Marine Mammal Medicine, Second edition pp. 309-335. |
|
|
Edgar RC (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26:2460-2461. |
|
|
Falagas ME, Kastoris AC, Vouloumanou EK, Dimopoulos G (2009). Community-acquired Stenotrophomonas maltophilia infections: a systematic review. European Journal of Clinical Microbiology and Infectious Diseases 28:719-730. |
|
|
Flemming L, Rawlings D, Chenia H (2007). Phenotypic and molecular characterisation of fish-borne Flavobacterium johnsoniae-like isolates from aquaculture systems in South African. Research in Microbiology 158:18-30. |
|
|
Fujita J, Yamadori I, Xu G, Hojo S, Negayama K, Miyawaki H, Yamaji Y, Takahara J (1996). Clinical features of Stenotrophomonas maltophilia pneumonia in immunocompromised patients. Respiratory Medicine 90:35. |
|
|
Gargiulo C, Pham VH, Hai NT, Nguyen KCD, Davey KF, Abe K (2015). A case report of an elderly patient with respiratory failure caused by Moraxella osloensis infection. Anaesthesia 1(1):17-21. |
|
|
Gaydos JK, Iii KCB, Osborne RW, Dierauf L (2004). Evaluating potential infectious disease threats for southern resident killer whales, Orcinus orca: a model for endangered species. Biological Conservation 117:253-262. |
|
|
Grimont PAD, Grimont F (1984). Family I. Enterobacteriaceae. Genus VIII. Sermtia Bizio 1823. In: Krieg NR, Holt JG, eds. Bergey's Manual of systematic bacteriology, Baltimore: Williams and Wilkins 1:477-484. |
|
|
Hanes SD, Demirkan K, Tolley E, Boucher BA, Croce MA, Wood GC, Fabian TC (2002). Risk factors for late-onset nosocomial pneumonia caused by Stenotrophomonas maltophilia in critically ill trauma patients. Clinical Infectious Diseases 35:228-235. |
|
|
Harper CMG, Dangler CA, Xu S, Feng Y, Shen Z, Sheppard B, Stamper A, Dewhirst FE, Paster BJ, Fox JG (2000). Isolation and characterization of a helicobacter sp. from the gastric mucosa of dolphins, Lagenorhynchus acutus and Delphinus delphis. Applied and Environmental Microbiology 66:4751-4757. |
|
|
Harris NB, Rogers DG (2001). Septicemia associated with Stenotrophomonas maltophilia in a West African dwarf crocodile (Osteolaemus tetraspis subsp. tetraspis). Journal of Veterinary Diagnostic Investigation 13:255-258. |
|
|
Hou Q, Xu H, Zheng Y, Xi X, Kwok LY, Sun Z, Zhang H, Zhang W (2015). Evaluation of bacterial contamination in raw milk, ultra-high temperature milk and infant formula using single molecule, real-time sequencing technology. Journal of Dairy Science 98:8464-8472. |
|
|
Jauniaux TP, Brenez C, Fretin D, Godfroid J, Haelters J, Jacques T, Kerckhof F, Mast J, Sarlet M, Coignoul FL (2010). Brucella ceti infection in harbor porpoise (Phocoena phocoena). Emerging Infectious Diseases 16:1966-1968. |
|
|
Kiska DL, Gilligan PH (2003). Pseudomonas. In: Murray PR, Baron EJ, Jorgensen JH, Pfaller MA, Yolken RH, eds. Manual of clinical microbiology. 8th ed. Vol. 1. Washington, DC: American Society for Microbiology Press pp. 719-728. |
|
|
Kulakov LA, Mcalister MB, Ogden KL, Larkin MJ, O'Hanlon JF (2002). Analysis of bacteria contaminating ultrapure water in industrial systems. Applied and Environmental Microbiology 68:1548-1555. |
|
|
Langille MGI, Zaneveld J, Caporaso JG, McDonald D, Knights D, Reyes JA, Clemente JC, Burkepile DE, Thurber RLV, Knight R, Beiko RG, Huttenhower C (2013). Predictive functional profiling of microbial communities using 16S rRNA marker gene sequences. Nature Biotechnology 31(9):814-821. |
|
|
Lee WJ, Hase K (2014). Gut microbiota-generated metabolites in animal health and disease. Nature Chemical Biology 10:416-424. |
|
|
Lim CS, Oh YS, Lee JK, Park AR, Yoo JS, Rhee SK, Roh DH (2011). Flavobacterium chungbukense sp. nov., isolated from soil. International Journal of Systematic and Evolutionary Microbiology 61:2734-2739. |
|
|
Liu Y (2009). The analyses of genetic diversity MHC-DQB exon 2 of finless porpoises in Bohai and north Yellow Sea. Shandong University Master Dissertation pp. 1-57. |
|
|
Olsen AB, Birkbeck TH, Nilsen HK, MacPherson HL, Wangel C, Myklebust C, Laidler LA, Aarflot L, Thoen E, Nygård S, Thayumanavan T, Colquhoun DJ (2006). Vaccine-associated systemic Rhodococcus erythropolis infection in farmed atlantic salmon Salmo salar. Diseases of Aquatic Organisms 72:9-17. |
|
|
Paterson DL (2006). The epidemiological profile of infections with multidrug-resistant Pseudomonas aeruginosa and Acinetobacter species. Clinical Infectious Diseases 43:43-48. |
|
|
Pootakham W, Mhuantong W, Yoocha T, Putchim L, Sonthirod C, Naktang C, Thongtham N, Tangphatsornruang S (2017). High resolution profiling of coral-associated bacterial communities using full-length 16S rRNA sequence data from PacBio SMRT sequencing system. Scientific Reports 7:2774. |
|
|
Quast C, Pruesse E, Yilmaz P, Gerken J, Schweer T, Yarza P, Peplies J, Glöckner FO (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Research 41(Database issue), pp. 590-596. |
|
|
Rhoads A, Au KF (2015). PacBio Sequencing and its applications. Genomics Proteomics Bioinformatics 13:278-289. |
|
|
Ryan MP, Pembroke JT, Adley CC (2006). Ralstonia pickettii: a persistent gram-negative nosocomial infectious organism. Journal of Hospital Infection 62:278-284. |
|
|
Salipante SJ, Kawashima T, Rosenthal C, Hoogestraat DR, Cummings LA, Sengupta DJ, Harkins TT, Cookson BT, Hoffman NG (2014). Erratum to Performance comparison of Illumina and ion torrent next-generation sequencing platforms for 16S rRNA-based bacterial community profiling. Applied and Environmental Microbiology 80:7583-7591. |
|
|
Schmidt PA, Bálint M, Greshake B, Bandow C, Römbke J, Schmitt I (2013). Illumina metabarcoding of a soil fungal community. Soil Biology and Biochemistry 65:128-132. |
|
|
Shah SS, Ruth A, Coffin SE (2000). Infection due to Moraxella osloensis: case report and review of the literature. Clinical Infectious Diseases 30:179-181. |
|
|
Shannon CE (1948). A Mathematical Theory of Communication. The Bell System Technical Journal 27:379-423. |
|
|
Sharp KH, Distel D, Paul VJ (2012). Diversity and dynamics of bacterial communities in early life stages of the Caribbean coral Porites astreoides. The ISME Journal 6:790-801. |
|
|
Simpson EH (1949). Measurement of Diversity. Nature 163:688. |
|
|
Sun JX, Wang Y, Liu D, Wang LX (2011). An ambiguous concept: clarification on conditioned pathogen. Microbiology China 38:1584-1586. |
|
|
Vuori-Holopainen E, Pehola H (2001). Reappraisal of lung tap: Review of an old method for better etiologic diagnosis of childhood pneumonia. Clinical Infectious Diseases 32(5):715-726. |
|
|
Wang Z, Xu J, Li Y, Wang K, Wang Y, Hong Q, Li WJ, Li SP (2010). Rhodococcus jialingiae sp. nov., an actinobacterium isolated from sludge of a carbendazim wastewater treatment facility. International Journal of Systematic and Evolutionary Microbiology 60:378-381. |
|
|
Xu JL, He J, Wang ZC, Wang K, Li WJ, Tang SK, Li SP (2007). Rhodococcus qingshengii sp. nov., a carbendazim-degrading bacterium. International Journal of Systematic and Evolutionary Microbiology 57:2754-2757. |
|
|
You SJ (2005). The microorganism community in an aerobic denitrification membrane bioreactor by 16S rDNA based molecular biotechnology. Environmental Technology 26:767-772. |
|
|
Zhou K (2004). Fauna Sinica, Mammlia. Vol. 9: Cetacea, Carnivora: Phocoidea, Sirenia.Beijing: Science Press. |
|
|
Zhou X, Guang X, Sun D, Xu S, Li M, Seim I, Jie W, Yang L, Zhu Q, and Xu J. Gao Q, Kaya A, Dou Q, Chen B, Ren W, Li S, Zhou K, Gladyshev VN, Nielsen R, Fang X, Yang G (2018). Population genomics of finless porpoises reveal an incipient cetacean species adapted to freshwater. Nature Communications 9:1276. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


