ABSTRACT
Many strains of Klebsiella pneumoniae can produce several virulence factors, such as siderophores, capsules, fimbrial adhesins and antibiotic resistance enzymes, which are important in the adherence, colonization, invasion and development of the infection. Therefore the aim of this study was to investigate ten virulence gens in 32 K. pneumoniae isolated from different clinical infections: urine (13), burns (12), sputum (4) and blood (3). Phenotypic methods were used to detect capsule, hypermucoviscosity, ability to form biofilm, produce siderophores, primary and confirmatory test of extended spectrum beta lactamase (ESBL). Polymerase chain reaction (PCR) technique was used to detect ten virulence genes encoding attachments (fimH, ycfM), siderophores (kfu: iron uptake system, entB: enterobactin, irp-2: yersiniabactin), capsule synthesis or invasions (rmpA, uge, wabG) and beta lactamase enzymes (SHV, TEM). Antimicrobial susceptibility was tested by disk diffusion method. According to phenotypic methods, the results demonstrated that, the capsule and siderophores production and biofilm formation were observed in 100% of isolates, while hypermucoviscosity phenomenon was observed in 62.5%. The most common virulence genes were fimH-1, ycfM and entB (100%), uge and TEM (93.75%), wabG and SHV (87.5%). Kfu and rpmA genes were found at medium rates of 65.62 and 62.5%, respectively and at lower prevalence was gene Irp-2 (37.5%). A rate of 84.375% of isolates showed a multidrug resistance (MDR) pattern, 12.5% extensive drug resistance (XDR), 3.12% pandrug resistance (PDR) and the rate of extended-spectrum β- lactamases (ESBL) producing K. pneumoniae was 62.5%.The study concluded that fimH-1, ycfM and entB genes were commonly found in all isolates, they seem to be at the basis of classic pathogenicity of K. pneumoniae. The most common virulence genes were observed in isolates from burns and blood samples. K. pneumoniae became highly resistant to antibiotics especially to 3rd generation cephalosporins and there was positive relationship between presence of virulence genes and ability of the bacterium to antibiotics resistance.
Key words: Klebsiella pneumoniae, drug resistance, virulence genes, Iraq.
Klebsiella pneumoniae is one of the most important pathogenic bacteria. It is gram negative, bacilli, non-
motile and causative agent of many diseases, such as pneumonia, urinary tract infections, bacteremia, burns and wounds infections and pyogenic liver abscesses (Rahamathulla et al., 2016). Pathogenicity of K. pneumoniae is due to the presence of many virulence genes which encode virulence factors that allow it to attack the immune system of mammalians and cause many kind of diseases. Some of these virulence factors are: biofilm formation, hypermucoviscosity, capsule synthesis, adhesions, iron uptake and lipopolysaccharides formation (Fertas-Aissani et al., 2013; Chung et al., 2015). K. pneumoniae has been found capable to resist many antibiotics especially third generation cephalosporins like cefotaxime, ceftriaxone and ceftazidime (Yeh et al., 2007). Many clinical features of K. pneumoniae infections are related with virulence gens according to number and mode of action of these genes (Wiskur et al., 2008). Recently K. pneumoniae is found causing acute liver abscess as reported in many Asian countries like China, Kuwait and Iraq (Christopher et al., 2014; Chung et al., 2015). K. pneumoniae mostly contains extended spectrum beta-lactamase genes (SHV, TEM and C-TXM) that encoded by plasmid. These genes have shown resistance to many types of antibiotics (more than three classes) which is considered as multidrug resistant bacteria (Ahmed et al., 2014). In spite of K. pneumoniae is considered as one of the most important opportunistic bacteria, knowledge of the mechanism by which this bacterium causes many diseases is still not fully understood. The aim of this study was to investigate the presence of virulence genes in 32 K. pneumoniae clinical isolates from different clinical sources.
Bacterial isolates
This study was carried out in Al-Sadder Medical City in AL-Najaf Governorate-Iraq. A total of 464 specimens from different clinical sources were collected from patients with (urinary tract infections, respiratory tract infections, burns infections and blood infections) during the period of January to October 2015. All urine and sputum samples were collected in sterile containers, a loopful of the urine and sputum samples were streaked onto the surface of MacConkey agar and blood agar (Oxoid,UK) and incubated at 37°C for 24 h. Burns samples were collected by sterile swabs and streaked onto the surface of MacConkey agar and blood agar (Oxoid, UK) and incubated at 37°C for 24 h. Five milliliter of blood samples were collected by a sterile syringes and mixed with 45 ml of brain heart infusion broth (Oxoid, UK) and incubated at 37°C for 7 days , and streaked (by sterile loop) onto the surface of blood and MacConkey agar (Oxoid, UK) and incubated at 37°C for 24 h (Collee et al.,1996). All isolates were identified according to morphological and biochemical tests (Gram stain, Growth on MacConkey Agar, Capsule stain, Catalase and oxidase test, Urease production test, Motility test, Indole production test, Methyl Red test, Voges-Proskauer test, Simmons Citrate test and Triple Sugar Iron test) (MacFaddin, 2000) and then by cultivation on Chrome agar medium (Orientation company – France). Finally all suspected isolates of K. pneumoniae were identified by using Vitek2® system (BioMerieux® -France).
Phenotypic determination of virulence factors
Capsule detection
The presence of capsule was investigated by staining with nigrosin, a loopful of overnight bacterial colony was transferred on a dry and clean slide, then gently mixed with nigrosin and allowed to dry in air, then rinsed with water , the slide was stained with methylen blue for 2 min and allowed to air dry, then the slide was gently washed with water, under light microscope the nigrosin stain provides a dark background to unstained capsule and metylene blue stain provides blue color to the cells (Soensen, 1995).
Hypermucoviscosity testing
Single colonies after culturing on Brain Heart Infusion agar plates (Oxoid, UK) were obtained and tested for their ability to form viscous strings. When a standard inoculation loop was touched onto their surface and slowly raised. The formation of string greater than 5 mm in length is indicative of hypermucoviscosity positive phenotype (Grange, 1988).
Siderophores production assay
Nutrient agar supplemented with 200 mM of 2.2’-dipyridyl was used as iron-restricted agar medium. All bacterial isolates were streaked on agar plates, and then incubated at 37°C for 24 h. Any bacterial growth was considered as positive results for ability of bacteria to siderophores production (Schwyn and Neilands, 1987).
Biofilm formation testing
Briefly, K. pneumoniae isolates were sub cultured three times in LB broth for 18 h at 37°C. The optical density was adjusted to 0.55 to 0.65 (2.106 to 8.109 CFU/mL) at 540 nm. Two hundred milliliter of the adjusted bacterial cultures were transferred to 96-well polystyrene microtiter plates and incubated for 24 h at 25°C. After addition of 25 mL of 1% crystal violet to each well, the plate was shaken and incubated for 15 min at room temperature. After removal of medium and three washes with phosphate-buffered saline, crystal violet was dissolved by the addition of ethanol, and the absorbance (Izquierdo et al., 2003).
DNA extraction
DNA was extracted by a boiling method as follows: Three to five pure and fresh colonies were suspended in 300 µl of distilled water, then cells were lysed by heating at 100°C for 20 min (in water bath), immediately the cells were placed in ice for 30 mi and the other cellular components was removed by centrifugation at 8500
rpm for 10 min. Finally the supernatant was used as the DNA template (Yang et al., 2008).
PCR detection of virulence-associated genes
PCR was used to detect genes encoding type 1 adhesins (fimH-1), outer membrane lipoprotein (ycfM), enterobactin biosynthesis (entB), yersiniabactin biosynthesis (irp-2), iron uptake system (kfu), regulator of mucoid phenotype A (rmpA), invasions (uge, wabG) and beta lactamase enzymes (SHV , TEM). All primers used are listed in Table 1. And polymerase chain reaction (PCR) thermo cycling conditions are listed in Table 2.
Antimicrobial susceptibility testing
Antibiotic susceptibility testing was performed by the Kirby-Bauer method according to the Clinical Laboratory Standards Institute (CLSI, 2014). A colony from each K. pneumoniae isolate was grown overnight in Mueller Hinton broth (Oxoid,UK) at 37°C. Bacterial cultures were adjusted to 0.5 on the MacFarland nephelometer scale (1.5 × 108 CFU/ml) and plated on Mueller Hinton agar (Oxoid, UK) by the streaking method using a sterile swab. Antimicrobial susceptibility and resistance was determined by isolate growth zone diameter according to CLSI guidelines (2014). E. coli ATCC 25922 strain was used as controls. Antibiotics disks were purchased from Bioanalyse, Turkey. All types of antibiotics are listed in Table 3.
Primary test for production of ESBL
Antibiotic susceptibility testing was done to three types of 3rd generation cephalosporins antibiotics: ceftazidime, cefotaxime, and ceftriaxone. If inhibition zone for bacterial isolates were: ≤27 mm for cefotaxime, ≤22 mm for ceftazidime and ≤25 mm for ceftriaxone, this result considered as positive result for production of extended spectrum beta lactamase (CLSI, 2006).
Confirmatory test for ESBL
Augmentin disc 30 μg was placed in the center of Mueller Hinton agar plate (Oxoid, UK). Around of three sides of augmentin disc (30 μg), a disc of ceftazidime (30 μg), ceftriaxone (30 μg) and cefotaxime (30 μg) were placed with distance of 15mm from center to center of augmentin disc. Then the plate was incubated at 37°C for 24 h.
If inhibition zone was increased towards the augmentin disc that considered as positive results for production of ESBL (
Sarojamma and
Ramakrishna, 2011).
Statistical analysis
Statistical analysis was performed with GraphPad Prism version 5 software, percentages was used for the comparison between samples of the study.
K.pneumoniae isolates
Out of the 464 total specimens there were 243 specimens (52.370%) it has been diagnosed as gram negative bacteria, while there were 196 specimens (42.241%) were gram positive bacteria and 25 specimens (5.389%) with no any growth (Table 4). A total of 243 g negative bacterial isolates were collected from different clinical sources (106 from urine, 90 from burns, 25 from sputum and 22 from blood) there were 13 isolate of K. pneumoniae (12.264%) from urine, 12 isolates (13.333%) from burns, 4 isolates (16%) from sputum and only 3 isolates (13.636%) from blood (Table 5).
Virulence factors
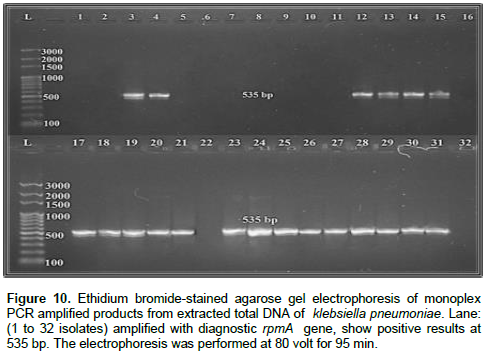
In the present study, phenotypic methods were used to detect virulence factors, the capsule, biofilm formation and siderophores productions were observed in 32 isolates (100%), hypermucoviscosity phenomenon was observed in 20 isolates (62.5%) (Figure 1). Out of total 32 isolates, 32 isolates (100%) were positive for fimh1, ycfm and entB genes (Figures 2, 3 and 4, respectively), 30 isolates (93.75%) were positive for uge and TEM genes (Figures 5 and 6 , respectively), 28 isolates (87.5%) were positive for wabG and SHV genes (Figures 7 and 8, respectively), 21 isolates (65.62%) were positive for kfu gene (Figure 9), 20 isolates (62.5%) were positive for rpmA gene (Figure 10) and 12 isolates (37.5%) were positive for Irp-2 gene (Figure 11). The prevalence and distribution of virulence factors were given in Tables 6 and 7.
Antimicrobial susceptibility
All isolates of K. pneumoniae showed resistance to AX, TIC and F (100%, 32/32), AMC and CTX (97.75%, 30/32), CRO (87.5%, 28/32), CAZ (71.875% 23/32), CIP (43.75%, 14/32), DO (40.62%, 13/32), TM and TE (34.37, 11), CN and C (31.25%, 10/32), AK (28.12% , 9/32), MEM(25% , 8/32) and IMP (18.75% 6/32). Also in this study, 84.37% (27/32) of K. pneumoniae isolates were MDR, 12.5% (4/32) were XDR and 12% (1/32) were PDR. Both primary and confirmatory test for ESBL production were positive for 62.5% (20/32) of the isolates resistant to third-generation cephalosporins (Figures 12 and 13). Resistance test to 16 antibiotic of K. pneumoniae isolates were given in Tables 8, 9 and 10.
The genes fimH-1 and ycfm were found in all of our isolates. This result is in agreement with the ubiquitous nature of this structure in K. pneumoniae as reported in Fertas-Aissani et al. (2013). Microorganisms with biofilm formation are associated with many human infections. The biofilm formation role and development by bacteria has been documented to be a crucial step in the pathogenesis of Klebsiella. The flushing action of sterile urine in urinary tract is protected from colonization by pathogen, but formation of biofilms by bacteria in urinary tract was lead to infect by chronic urinary tract infections. Biofilms and outer membrane lipoproteins can play an important role in protecting bacteria from drugs exposure when compared with other bacteria do not have these virulence factors (Bellifa et al.,2013). Therefore, there was strong relationship between resistance of antibiotics and prevalence of these virulence factors in bacteria. Bacteria with biofilm forming are generally more resistant to many antibiotics. Biofilms and outer membrane lipoproteins act as biodegradable effect on beta-lactamases antibiotics. Beta-lactamase enzymes are secreted and maintain their activity inside of biofilm matrix, and decompose beta-lactam antibiotics before these antibiotics reach the bacterial cells (Høiby et al., 2010). Antibiotics penetration inside of biofilms could be blocked by other factors, such as the presence of surfaces with negatively-charged, particularly for large polar molecules such as aminoglycosides antibiotics with positively charged, Also, trace amount of metabolic rates and limited oxygen are maybe important factors contributing to increasing resistance to fluoroquinolones, cephalosporins, aminoglycosides and beta-lactamase antibiotics (Kwon et al., 2008; Lewis, 2001).
Siderophores are bio-synthetically compounds produced and secreted by many microorganisms such as, fungi and bacteria for iron (Fe+3) uptake, and they are selective chelators. These ions are weakly soluble and almost found in all oxygenated environments. In this study, Entb gene was found in all our isolates (100%, 32/32), while irp-2 gene was found in some of our isolates (37.5%, 12/32). These results were in agreement with some previous studies showed that almost all K. pneumoniae clinical isolates were having these chelators (Podschun et al., 1993; Koczura and Kaznowski, 2003; Fertas-Aissani et al., 2013) also in this study, kfu gene was found in 21 isolates (65.62%). Enterobactin and yersiniabactin they are siderophore compounds produced by bacterial cells to uptake iron (Fe+3) from iron-binding proteins of the host, and they are have strong-affinity extracellular ferric chelators, which; they have an important role in pathogenesis and virulence of bacteria, virulence of these compounds are remain unclear; however, the expression of enterobactin induced biofilm formation (May and Okabe, 2011). But Lai et al. (2001) suggested that the expression of gene for iron-enterobactin that responsible for outer membrane receptor appears to be activated during bacterial infection.
The kfu gene is a putative pathogenic gene which codes for an iron uptake system. kfu gene associated with the purulent tissue infections ,capsule formation and virulent hypermucoviscosity phenotype. Therefore, this gene is considered as a very important gene in iron up take from host cell (Aher et al., 2012).
In this study, all our isolates had a capsule, while only 62.5% (20/32) had a hypermucoviscosity phenotype and rpmA gene. Almost K. pneumoniae produce large amounts of muco-polysaccharide mass and extra-capsular polysaccharides to produce strain with more virulent (Wiskur et al., 2008). In line with this Victor et al. (2007) reported that mucoid phenotype was seen in all isolates of K. pneumoniae that caused the invasive syndrome and in more than 90% of isolates in human with community- acquired pneumonia and in South Africa and Taiwan. Also he found that mortality of laboratory animals injected with mucoid strains was higher than that occurring in the same laboratory animals injected with non-mucoid strains.
The rmpA gene is a main factor that has an important role in virulence of K. pneumoniae strains, is controlled by plasmid with strongly mucoviscous phenotype promoted and synthesis regulator of the capsular polysaccharide (Rivero et al., 2010). The results demonstrated that out of total 32 isolates, 30 isolates (93.75%) were positive for uge gene and 28 isolates (87.5%) were positive for wabG gene.
wabG gene is less virulence factor studied, although it has been observed that many of wabG gene positive strains are from the patients with invasive and serious infections. But the mechanism of mode of action is still unclear (Turton et al., 2010). Izquierdo et al. (2003) proved that strains of K. pneumoniae with mutant wab G gene were non capsulated and less virulent in murine pneumonia model (rats). This fact proves the important role of wab G gene in pathogenicity of K. pneumoniae. Because the defect in synthesis of core lipopoly-saccharides in strains of K. pneumoniae with mutant wab-G gene were unable to induce urinary tract infection in experimental rats. Since they were unable to colonize in urinary tract in comparison with that of the normal strains. But the mutant bacterial strains were still able to perform some colonization. Regué et al. (2004) proved that the mutant strains of K. pneumoniae (without uge gene) were non-virulent in laboratory animals; this fact proves the important role of uge gene in pathogenicity of K. pneumoniae.
The antibiotic susceptibility analysis of the isolates showed that 100% (32/32) were resistance to amoxicillin, ticarcillin and nitrofurantoin. On the other hand, 84.37% (27/32) of isolates were MDR, 12.5% (4/32) were XDR and 12% (1/32) were PDR. The presumptive test for ESBL production was positive for 65.5% (20/32). The genes TEM and SHV were 93.75% (30/32) and 87.5% (28/32). These resistance patterns can be due to the expression of different enzymes such as extended-spectrum beta lactamases. Multi-drug resistances are often associated with extended spectrum beta-lactamase producing bacteria, that is, resistance to other classes of drugs like aminoglycosides and quinolones (Roshan et al., 2011). Isolates of K. pneumoniae may be have natural resistance to amoxicillin, ampicillin and ticarcillin but not to ESBL antibiotics. The resistance to Extended Spectrum Beta-Lactamase could happen class A chromosome beta-lactamase SHV and TEM genes are expressed (Mendonca and Ferreira, 2009). K. pneumoniae plasmids contain many B-lactamase genes including those encoding AmpC beta-lactamases, extended-spectrum B-lactamases, inhibitor resistant, metallo enzymes, SHV and TEM beta-lactamases. These enzymes make bacteria enable to resistant to different antimicrobial agents including Imipenem, Meropenem, 3rd generation cephalosporins and others (Moland et al., 2003; Poirel et al., 2004; Essack et al., 2009). Padilla et al. (2010) reported that resistance mechanisms such as efflux pump can contribute to virulence of Klebsiella, and there was positive relationship between production of ESBL with virulence factors (Wiskur et al., 2008). As conclusions fimH-1, ycfM and entB genes were commonly found in all isolates, they seem to be at the basis of classic pathogenicity of K. pneumoniae. The most common virulence genes were observed in clinical sources of burns and blood infections. K. pneumoniae became highly resistant to antibiotics especially to 3rd generation cephalosporins. There was positive relationship between resistance to antibiotics and prevalence of virulence factors in K. pneumoniae.
Study the prevalence of another virulence gene in k. pneumoniae such as kpn, traT and iroN. And study the prevalence of virulence genes in another bacterium like serratia and streptococcus pneumoniae.
This work entitled (Virulence factors and antibiotic susceptibility patterns of multidrug resistance K. pneumoniae isolated from different clinical infections) was funded by the Faculty of Science University of Kufa department of biology.
The authors are grateful to staff of microbiology laboratory in AL-sadder medical hospital, Iraq, for providing the clinical samples.
REFERENCES
|
Aher T, Roy A, Kumar P (2012). Molecular Detection of Virulence Genes Associated with Pathogenicity of Klebsiella spp. isolated from the respiratory tract of apparently healthy as well as sick goats. Isr. J. Vet. Med. 67(4):249-52.
|
|
|
|
Ahmed OI, Soha AE, Tamer MA, Iman ZA (2014). Detection of bla SHV and bla CTX-M genes in ESBL producing Klebsiella pneumoniae isolated from Egyptian patients with suspected nosocomial infections. Egypt. J. Med Hum. Genet .14:277-283.
Crossref
|
|
|
|
Bellifa S, Hassaine H, Balestrino D, Charbonnel N, M'hamedi I, Terki IK, Lachachi M, Didi W, Forestier C (2013). Evaluation of biofilm formation of Klebsiella pneumoniae isolated from medical devices at the University Hospital of Tlemcen, Algeria. Afr. J. Microbiol. Res. 7(49):5558-5564.
|
|
|
|
Brisse S, Fevre C, Passet V (2009). Virulent clones of Klebsiella pneumoniae: identification and evolutionary scenario based on genomic and phenotypic characterization. PLoS One. 4(3):e4982.
Crossref
|
|
|
|
Chung TH, Karkey A, Pham TD, Boinett CJ, Cain AK, Ellington M (2015). A high-resolution genomic analysis of multidrug resistant hospital outbreaks of Klebsiella pneumoniae. EMBO Mol. Med. 7(3):227-39.
Crossref
|
|
|
|
Christopher AB, Michelle P, Virginia LM (2014). Klebsiella: A long way to go towards understanding this enigmatic jet-setter. F1000Prime Reports. 6:64.
|
|
|
|
Clinical Laboratory Standards Institute CLSI (2006). Performance standards for antimicrobial susceptibility testing, in Proceedings of the 16th International Supplement (M100-S16), National Committee for Clinical Laboratory Standards, Wayne, PA, USA.
|
|
|
|
Clinical and Laboratory Standards Institute (CLSI) (2014). Performance Standards for Antimicrobial Susceptibility Testing; 22ed. Informational Supplement. PA, USA 32(3).
|
|
|
|
Ensor VM, Jamal W, Rotimi VO, Evans JT, Hawkey PM (2009). Predominance of CTX-M-15 extended spectrum beta-lactamases in diverse Escherichia coli and Klebsiella pneumoniae from hospital and community patients in Kuwait. Int. J. Antimicrob. Agents 33:487-489.
Crossref
|
|
|
|
Essack SY, Hall L. MC, Livermore DM (2004). Klebsiella pneumoniae isolate from South Africa with multiple TEM, SHV and AmpC beta-lactamases. Int. J. Antimicrob. Agents. 23:398-400.
Crossref
|
|
|
|
Fertas-Aissani R, Messai Y, Alouache S, Bakour R (2013). Virulence profiles and antibiotic susceptibility patterns of Klebsiella pneumonia strains isolated from different clinical specimens. Pathol. Biol. 61(5):209-216.
Crossref
|
|
|
|
Grange JM (1988). Mycobacteria and human disease. Edward Arnold, London.
|
|
|
|
Høiby N, Bjarnsholt T, Givskov M, Molin S, Ciofu O (2010). Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents. 35:322-32.
Crossref
|
|
|
|
Izquierdo L, Coderch N, Piqué N, Bedini E, Corsaro MM, Merino S, Fresno S, Tomás JM, Regué M (2003).The Klebsiella pneumoniae wabG gene: role in biosynthesis of the core lipopolysaccharide and virulence. J. Bacteriol. 185(24):7213-7221.
Crossref
|
|
|
|
Koczura R, Kaznowski A (2003). Occurrence of the Yersinia high-pathogenicity island and iron uptake systems in clinical isolates of Klebsiella pneumoniae. Microb. Pathog. 35:197-02.
Crossref
|
|
|
|
Kwon AS, Park GC, Ryu SY, Lim DH, Lim DY, Choi CH (2008). Higher biofilm formation in multidrug-resistant clinical isolates of Staphylococcus aureus. Int. J. Antimicrob Agents. 32:68-72.
Crossref
|
|
|
|
Lai YC, Peng HL, Chang HY (2001). Identification of genes induced in vivo during Klebsiella pneumoniae CG43 infection. Infect. Immun . 69(11):7140-5.
Crossref
|
|
|
|
Lewis K (2001). Riddle of biofilm resistance. Antimicrob Agents Chemother. 45:999-1007.
Crossref
|
|
|
|
Ma LC, Fang CT, Lee CZ, Shun CT and Wang JT (2005). Genomic heterogeneity in K. pneumoniae strains is associated with primary pyogenic liver abscess and metastatic infection. J. Infect. Dis. 192:117-128.
Crossref
|
|
|
|
MacFaddin JF (2000). Biochemical tests for identification of medical bacteria . (3rd edition).The Williams and Wilkins-Baltimor. USA.
|
|
|
|
May T, Okabe S (2011). Enterobactin is required for biofilm development in reducedgenome Escherichia coli. Environ. Microbiol. 13:3149-62.
Crossref
|
|
|
|
Mendonca N, Ferreira E (2009). Genetic diversity of genes encoding OKP and LEN b-lactamases produced by clinical Klebsiella pneumoniae strains in Portugal. Diagn. Microbial. Infect. Dis. 63:334-38.
Crossref
|
|
|
|
Moland ES, Hanson ND, Herrera VL, Black JA, Lockhart T, Hossain J, Johnson JA, Goering RV, Thomson KS (2003). Plasmid-mediated, carbapenem-hydrolysing B-lactamase, KPC-2, in Klebsiella pneumoniae isolates. J. Antimicrob. Chemother. 51:711-14.
Crossref
|
|
|
|
Padilla E, Llobet E, Dome’nech-Sa’nchez A, Martı’nez-Martı’nez L, Bengoechea JA, Albertı´ S (2010). Klebsiella pneumoniae AcrAB efflux pump contributes to antimicrobial resistance and virulence. Antimicrob. Agents Chemother. 54:177-183.
Crossref
|
|
|
|
Podschun R, Sievers D, Foscher A, Ullmann U (1993). Serotypes, hemagglutinins, siderophore synthesis, and serum resistance of Klebsiella isolates causing human urinary tract infections. J. Infect Dis.168:1415-1421.
Crossref
|
|
|
|
Poirel L, He´ ritier C, Tolu¨n V, Nordmann P (2004). Emergence of oxacillinase-mediated resistance to imipenem in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 48:15-22.
Crossref
|
|
|
|
Rahamathulla MP, Harish BN, Mataseje L, Mulvey MR (2016). Carbapenem resistance mechanisms among blood isolates of Klebsiella pneumoniae and Escherichia coli. Afr. J. Microbiol. Res. 10(2):45-53.
Crossref
|
|
|
|
Regué M, Hita B, Piqué N, Izquierdo L, Merino S, Fresno S, Benedí VJ, Tomás JM (2004). A Gene uge is essential for Klebsiella pneumoniae virulence. Infect Immun. 72(1):54-61.
Crossref
|
|
|
|
Rivero A, Gomez E, Alland D, Huang D, Chiang T (2010). K2 serotype Klebsiella pneumoniae causing a liver abscess associated with infective endocarditis. J. Clin. Microbiol. 48(2):639-641.
Crossref
|
|
|
|
Roshan M, Ikram A, Mirza IA, Malik N, Abbasi A, Alizai SA (2011). Susceptibility pattern of extended spectrum ß- lactamase producing isolates in various clinical specimens. J. Coll. Phys. Surge Pak. 21(6):342-346.
|
|
|
|
Sarojamma V, Ramakrishna V (2011). Prevalence of ESBL-producing Klebsiella pneumoniae isolates in Tertiary Care Hospital. ISRN Microbiology 5 p.
Crossref
|
|
|
|
Schubert S, Cuenca S, Fischer D, Heesemann J (2000). High-pathogenicity island of Yersinia pestis in Enterobacteriaceae isolated from blood cultures and urine samples: prevalence and functional expression. J. Infect Dis. 182:1268-1271.
Crossref
|
|
|
|
Schwyn B, Neilands JB (1987). Universal chemical assay for the detection and determination of siderophores. Anal. Biochem. 160:47-56.
Crossref
|
|
|
|
Soensen UBS (1995). Pneumococcal polysaccaride antigens: capsules and C-polysaccaride. Danish. Med. Bull. 42:47-53.
|
|
|
|
Turton JF, Perry C, Elgohari S, Hampton CV (2010). PCR characterization and typing of Klebsiella pneumoniae using capsular type-specific, variable number tandem repeat and virulence gene targets. J. Med. Microbiol. 59(5):541-547.
Crossref
|
|
|
|
Victor LY, Dennis SH, Wen CK, Asia S, Keith PK (2007). Virulence characteristics of Klebsiella and clinical manifestations of K. pneumoniae bloodstream infections. Emer. Infect. Dise .13(7): 986-93.
Crossref
|
|
|
|
Wiskur BJ, Hunt JJ, Callegan MC (2008). Hypermucoviscosity as a virulence factor in experimental Klebsiella pneumoniae endophthalmitis. Invest. Ophthalmol. Vis. Sci. 49:4931-4938.
Crossref
|
|
|
|
Yang JL, Wang MS, Cheng AC, Pan KC, Li CF, Deng SX (2008). A simple and rapid method for extracting bacterial DNA from intestinal microflora for ERIC-PCR detection. World J. Gastroenterol. 14(18):2872-2876.
Crossref
|
|
|
|
Yeh KM, Kurup A, Siu LK, Koh YL, Fung CP, Lin JC, Chen TL, Chang FY, Koh TH (2007). Capsular serotype K1 or K2, rather than magA and rmpA, is a major virulence determinant for Klebsiella pneumoniae liver abscess in singapore and Taiwan J. Clin. Microbiol. 45(2):466-471.
Crossref
|