Full Length Research Paper

ABSTRACT
ð›¼-Naphthylisothiocyanate (ANIT) is known to provoke liver damage with intrahepatic cholestasis. This ANIT is accepted to be important for assessing the effect of medicine instigated cholestasis. This assessment investigated the effect of green tea (Camellia sinensis) in contrast to that of Vitamin C (VC) against the hepatotoxicity of ANIT. Rats were orally treated with green tea portion (GT50, 75, 100 mg/kg) and CV (250 mg/kg) following 12 h of ANIT mixture (75 mg/kg). Rats were killed 24 h after treatment. Rats treated with ANIT demonstrated hepatocyte damage and cholestasis appeared as changes in serum biomarker levels, among others; increase in entire cholesterol, triglycerides, phospholipids, and lipid peroxide, and furthermore, an extension in hepatic lipid peroxide, a decline in glutathione and myeloperoxidase activity and a decrease in hepatic superoxide dismutase. The utilization of GT (75 mg/kg) to rats treated with ANIT blocked hepatic cell mischief and cholestasis and alleviated these serum and hepatic biochemical changes, while the use of GT (50 or 100 mg/kg) was less impressive. Both VC and ANIT-treated rats kept up a vital separation from liver cell hurt, yet not cholestasis, and reduced serum lipid peroxide, hepatic lipid peroxide, and myeloperoxidase action. These results demonstrate that the GT guarantees ANIT-affected liver harm (Cholestasis in the rat) more satisfactorily than VC.
Key words: Antioxidant, ð›¼-Naphthylisothiocyanate, cholestasis, green tea, Vitamin C.
INTRODUCTION
Green tea is the most expended drink and can deter the impact of cancer in vivo (Yang et al., 2009; Fujiki et al., 2018) Green tea (Camellia sinensis) has cell fortifying and weakening properties. The protective impact was credited to the cell reinforcement limit of the concentrate (Erba et al., 1999; Somia et al., 2017) and likely because of the gift of hydrogen by green tea polyphenols (Anderson et al., 2001). Correspondingly, green tea extricate anticipated H2O2-actuated cell demise just like manner bladder malignancy and urothelial cells (Coyle et al., 2008). Polyphenols in green tea are believed to be in charge of the protection impacts of disease. Every day utilization of polyphenols from green tea is high in a few nations (Lorenzo et al., 2016; Fujiki et al., 2018). The phenolic mixes of green tea with the most astounding focus are gallic acid, gallocatechin, catechin, epicatechin, epigallocatechin, epicatechin gallate, epigallocatechin gallate, p-coumaroylquinic corrosive and gallocatechin-3-gallate (Shishikura and Khokhar, 2005; Kwon et al., 2015; Riley et al., 2018). Green tea likewise contains dense and hydrolyzable tannins (Okuda and Ito, 2011; McAlpine et al., 2016) and has the most astounding centralization of polyphenols contrasted with different teas, including epigallocatechin gallate (EGCG), which might be the reason green tea can prompt apoptotic cell passing in malignant growth than different teas (Lin et al., 2003; Riley et al., 2018; Miyata et al., 2018). Green tea polyphenols have been exhibited against malignant growth action in various investigations that could be intervened by the antioxidant or pro-oxidant system in various rats models of human disease (Davalli et al., 2012; Naponelli et al., 2017; Fujiki et al., 2018). These polyphenols, for example, EGCG hinders cell feasibility and actuate apoptosis in various malignant growth cell lines, for example, osteogenic sarcoma (Ji et al., 2006), lymphoblastoid cells (Noda et al., 2007), leukemia cells (Nakazato et al., 2005), melanoma cells (Nihal et al., 2005), immune system microorganisms (Li et al., 2000), antibladder cancer (Miyata et al., 2018) and laryngeal carcinoma (Lee et al., 2010). Green tea polyphenols have appeared to lessen the separation of malignant growth cells into slower multiplying cells (Zhou et al., 2004). Be that as it may, the ramifications of oxidative or cell antioxidant instruments is vague; it is as yet indistinct whether green tea has a defensive impact against intense liver damage with cholestasis in vivo in rats.
α-Naphthylisothiocyanate (ANIT) is a substance usually used to start cholestasis by harming epithelial cells of the bile conduits and causing serious cholangitis and intrahepatic cholestasis (Yan et al., 2017;Wu et al., 2017; Han et al 2018). A one of a kind treatment of rats explored different avenues regarding α-naphthylisothiocyanate (ANIT) which can harm the liver with intrahepatic cholestasis (Plaa and Clerical, 1976; Kossor et al., 1993; Golbar et al., 2017). It is imagined that this hepatic injury with ANIT-initiated cholestasis is valuable for examining the procedures associated with medication instigated cholestasis, since hepatic sores and cholestasis exist on account of the application of certain explicit medications (e.g. Erythromycin estolate, Chlorpromazine, and so forth) to rats and people by emulating the application of ANIT to rats (Kossor et al., 1993; Yan et al., 2017;Han et al.,2018). ANIT instruments with cholestasis have been proposed; however, it has not yet been completely explained. It has been prescribed that lessened hepatic glutathione (GSH) be added to the movement of hepatic inclusion related with ANIT-prompted cholestasis because of its capacity to deliver a reversible ANIT-S conjugate which is basic for the vehicle of ANIT into the bile where it is discharged at high fixations and most likely lethal (Roth and Dahm, 1997; Yan et al., 2017; Wu et al., 2017). Likewise, it has been proposed that neutrophil-intervened aggravation might be added to the enhancement of ANIT-initiated cholestatic liver damage in rats (Roth and Dahm, 1997). Besides, in rats treated once with ANIT, it was affirmed that lipid peroxidation instigated by responsive oxygen species (ROS) made by methods for attacking neutrophils in the liver tissues is engaged with the movement of liver sores with cholestasis (Kongo et al., 1999; Cullen et al., 2016). Moreover, it has appeared in ANIT-treated rats that the intrusion of the hepatic disease hindering action framework adds to enhanced cholestasis-related liver harm (Ohta et al., 2001; Nakamura et al., 2013; Wu et al., 2017).
The destructive ascorbic acid, also called vitamin C (VC), is prominent for applying antioxidant and reducing exercises (Sorice et al., 2014). This investigation demonstrates that green tea extricates control weight actuated liver harm, in any event to a lesser degree, through its cell reinforcement and calming exercises identified with VC (Reddy et al., 2017). Furthermore, VC has been proven to hinder the harm to liver cells (Yang et al., 2018), yet not cholestasis, in rats treated with a solitary ANIT treatment, at any rate to some degree, through its cancer prevention agent and calming exercises (Ohta et al., 2006; Han et al., 2018). In this vein, this investigation inspected the guarded impact of green tea extricate on ANIT-actuated liver damage and cholestasis in rats contrasted with that of VC.
MATERIALS AND METHODS
Chemical
α-Naphthylisothiocyanate (ANIT), 3,3', 5,5'- tetramethylbenzidine (TMB), cow-like serum egg whites, p-coumaric corrosive, RRR-α-tocopherol (α-Toc), superoxide dismutase (Grass) decontaminated from ox-like erythrocytes, yeast glutathione reductase, NADPH, chlorogenic corrosive, epigallocatechin gallate (EGCG), L-ascorbic corrosive, cinnamic corrosive, chrysin, N, N-dimethylformamide (DMF), α'- dipyridyl, 5.5 Dithiobis (2-nitrobenzoic corrosive) (DTNB), ethylenediaminetetraacetic corrosive (EDTA), Folin-Ciocalteu reagent (FCR), gallic corrosive, kaempferol, quercetin, glutathione decrease (GSH), 2-thiobarburic acid, trichloroacetic acid (TCA), Tween 80 and different synthetic concoctions were bought from Sigma (St. Louis, Missouri, USA); α-Toc and delta-tocopherol standard utilized for the assurance of the CV.
Preparation of GT extract and determination its composition
A green tea extricate (GT) was set up by drenching about 30 g of dried green tea leaves homogenized in 100 ml of 95% ethanol at 90°C for 60 min. The cooled blend was sifted through a 0.45 μm millipore nylon channel before the examination. The sifted test was dried at 45°C.
The polyphenol substance of the GT was controlled by the Folin-Ciocalteau colorimetric system referenced by Ahn et al. (2004). The essential constituents of GT were decayed by HPLC as shown by the method portrayed by Friedman et al. (2006). HPLC was performed utilizing a Hitachi display 665-II fluid chromatograph with an autosampler (Demonstrate 655A-40, Hitachi Ltd., Tokyo, Japan). The section was (250 mm x 4.0 mm i.d.). The section temperature of 30°C was kept up utilizing a Shimadzu CTO-10vp segment heater (Shimadzu, Kyoto, Japan). The slope of the versatile stage was a blend of 100% acetonitrile and 20 mM KH2PO4. The stream rate was 1 mL/min. The photodiode cluster UV-VIS locator (SPD-10Avp model, Kyoto, Japan) was set somewhere in the range of 200 and 600 nm. The tea extricates (10 μL) was infused into the section. Recognition of every catechins, theaflavins, alkaloid, and artepillin C in the GT extricate were performed at 280 nm and the constituent of every substance was assessed. The constituent of strong parts was assessed at 14.3%. The catechins and the segments of the other two fundamental green tea, caffeine and theanine, are confirmed as just the extricated concentrates of 58, 25 and 12 mg/g of catechins, caffeine, and theanine, referenced by Chang et al. (2000).
Experimental animals and protocol
Six-week-old male rats were acquired from King Fahad Exploration Center, King Abdul Aziz Higher Institution, Jeddah, Saudi Arabia. The rats were housed in temperature-controlled pens (25 ± 2°C) and relative stickiness (52 ± 5%) with 12 h of light. The rats were kept with free access to rats sustenance and faucet water for seven days. All rats got kind consideration as per Umm Al-Qura Higher Institution rules.
The rats were isolated into 6 factions (n = 5 each) as shown below:
i) Control faction: The rats did not get ANIT, GT or VC.
ii) ANIT faction: The rats were dealt with just with ANIT.
iii) ANIT + GT faction (50): The rats were treated with ANIT and managed after GT application at a quantity of 50 mg/kg.
iv) ANIT + GT faction (75): The rats were treated with ANIT and BPEE at a quantity of 75 mg/kg was applied hereafter.
v) ANIT + GT faction (100): The rats were treated with ANIT and along these lines managed by GT at a quantity of 100 mg/kg.
vi) ANIT + VC faction: The rats were treated with ANIT and post-managed with VC. ANIT has been broken down in olive oil. All rats were fasted for 15 h before treatment and got water voluntarily amid the tests. Seven-week-old rats from all factions got an intraperitoneal (I.P.) infusion of ANIT at a portion of 75 mg for each kg of body weight, as portrayed by Kongo et al. (1999), Ohta et al. (2001, 2006). The control faction and alternate factions were treated with a similar volume of olive oil. Following 12 h of starting an infusion of ANIT or olive oil, the rats treated with ANIT were partitioned into 4 factions; Three factions of them were orally attended to, with GT at various quantities (50, 75 and 100 mg/kg) of green tea extricate with ethanol. The last faction of ANIT was treated with VC at a quantity of 250 mg/kg body weight. The quantity of VC utilized in this examination was resolved dependent on information introduced by Kaida et al. (2010). Following 24 h, the treated rats were yielded for histological examination and biochemical investigation.
Sample preparation
24 h after the main infusion of ANIT or vehicle, each rat was gauged and relinquished under ether anesthesia, and the blood was expelled from the mediocre vena cava. The serum was disconnected from the blood gathered by centrifugation. Not long after the killing, every liver was washed with super cold 0.9% NaCl to empty the blood into the tissues, at that point weighed subsequent to being wiped on a channel paper. The liver and serum tests were kept at - 80°C until its utilization.
Serum biomarkers investigation
Alanine aminotransferase (ALT) and aspartate aminotransferase (AST) in serum were directed utilizing a transaminase II test pack. γ-Glutamyl transpeptidase (γ-GTP) was settled in the serum utilizing the γ-GTP C-Test pack. The bilirubin and bile acid totals were inspected utilizing the Bilirubin BII-Test and Aggregate bili basic analysis units, individually. Serum adds up to cholesterol (T-Chol), while triglycerides and phospholipids were directed by the cholesterol test E test, the triglyceride test G packs, and the phospholipid C test, individually. The packs were obtained from Wako, Richmond, VA, USA. Lipid peroxide (LPO) was assessed using serum fluorometry by the methodology of thiobarbituric acid, as depicted by Yagi (1979). The excitation and transmission wave lengths of this gauge were 515 and 553 nm. The estimation of LPO in serum is communicated as malondialdehyde proportionate (MDA).
Determination of hepatic biomarkers
The liver was gathered and isolated from every trial faction and weighed independently. The heaviness of the liver was assessed by their relative weight (g/100 g B.W). Disconnected hepatic tissue was homogenized in 9 volumes of super cold 50 mM Tris-HCl support (pH 7.4), containing 1 mM EDTA, to set up a 10% homogenate utilizing a FisherbrandTM Q700 sonicator (Fisher, Hampton, New Hampshire, USA). In the wake of keeping the remaining on the ice for one moment, the homogenate was centrifuged at 4°C (10,000xg) for 20 min. A microdialysis device was utilized to dialyze the supernatant against 100 volumes of a comparative transporter at 4°C for 1 h (Fisher, Hampton, New Hampshire, USA). For estimation of GSH, ð›¼-Toc, and LPO, the subsequent liver homogenate was utilized. The DTNB procedure of Sedlak and Lindsay (1968) was utilized to gauge the dimension of GSH in liver homogenate. HPLC was utilized to assess α-Toc in the liver homogenate utilized as an interior standard, as demonstrated by Kamiya (2005). The spectrophotometer was utilized to evaluate LPO in a homogenate utilizing the thiobarbituric acid strategy depicted by Ohkawa et al. (1979). The estimation of liver LPO accounted for is like that of MDA reciprocals. Hepatic SOD movement was settled utilizing a business SOD test unit (Sigma, St. Louis, Missouri, USA). The techniques utilized by Cohen (1970) and Kamata et al. (1994) were utilized to decide catalase (Feline) and Se-glutathione peroxidase (Se-GSH-px), individually. Myeloperoxidase (MPO) was assessed as depicted by Suzuki et al. (1983), where MPO was utilized as a record of neutrophil invasion in tissues. The action of the MPO in the liver tissue supernatant was controlled by brooding of the supernatant at 60°C for 2 h to construct recuperation of MPO in liver tissues as revealed by Schierwagen et al. (1990). MPO movement in the warmed liver tissue test was evaluated by estimating the oxidation of the hydrogen peroxide-subordinate TMB at 37°C. The TMB was decayed to N, N-dimethylformamide. One unit (U) of this compound is described as estimating the catalyst, causing a change of the absorbance of 1.0 every moment at 655 nm. The protein in the supernatant was evaluated utilizing the Protein Test Fast unit (Wako, Richmond, VA, USA). The protein level in the liver tissue supernatant was resolved and the cow-like serum egg whites were utilized as a benchmark.
Histological study
Rat liver specimens treated with ANIT and GT or VC and untreated control rats were gathered and settled in phosphate cradled saline for 10 h with 10% formalin. The specimen was washed with faucet water, and afterward got dried out in liquor. The tissue was inserted in paraffin and cut into 5-7 μm thick areas. They were mounted on glass slides and recolored with hematoxylin and eosin (H&E). The histological perceptions were inspected under an optical magnifying lens.
Statistical analysis
The outcomes acquired are communicated as mean ± standard deviation (S.D.). The measurable investigation was resolved to utilize StatView statistical programming. Each mean was thought about by one-way analysis of variance (ANOVA). Numerous correlations between factions were then performed utilizing the Tukey-Kramer test, and the distinction of P < 0.05 was viewed as measurably huge.
RESULTS
Effects of GT and VC on liver cell damage and cholestasis
Serum ALT and AST demonstrate hepatocyte harm. In the ANIT faction, γ-GTP activity, a collection of bilirubin and bile acid obsession, and cholestasis were considerably higher than in the control faction (Figures 1 and 2). In the ANIT + GT (50), ANIT + GT (75) and ANIT + GT (100) factions, ANIT-driven serum ALT, AST, and γ-GTP action augmentations were recognized, while ANIT instigated serum delta ALT and AST movement was considerably weakened in the ANIT + VC faction (Figures 1A and 2B). The impact of ANIT + GT (50) on the expansion in ANIT-instigated serum ALT was in a general sense higher than in the ANIT + GT (75) and ANIT + GT (100) factions. The impact of ANIT-incited increments in AST and γ-GTP movement in the ANIT + GT (50) gathered was more articulated than in the ANIT + GT (75) and ANIT + GT (100) factions (Figures 1B and 2A). The impact of ANIT + GT (75) on the compression of expanded ANIT-prompted serum ALT and AST movement was tantamount to that of the ANIT + VC faction (Figure 1). In the ANIT + GT (75) faction, ANIT-prompted increments in serum add up to bilirubin and aggregate bile acid focuses were essentially weakened (Figures 2B and 2C). In the ANIT + GT (75) faction, ANIT-instigated increments in serum total bilirubin and bile acid obsession were basically restricted (Figures 2B and 2C). Be that as it may, in the ANIT + GT (50) faction, the ANIT-prompted increment in serum total bilirubin and aggregate bile acid obsession was not constricted (Figures 2B and 2C). In spite of the fact that the expansion in serum adds up to bilirubin fixation actuated by ANIT which was totally constricted in the ANIT + GT (100) faction, the weakening impact of ANIT + GT (100) totals was basically as low as that of ANIT + GT (75) (P < 0.05) (Figures 2B and 2C). In the ANIT + VC faction, the ANIT-initiated increment in serum add up to bilirubin and accumulated bile acid obsession was not constrained at all (Figures 2B and 2C).


Effects of GT and VC on live histological changes
In the control faction, the ANIT faction, the ANIT + GT (75) faction, and the ANIT + VC faction, the hematoxylin and eosin recolored liver regions were exposed to histopathological changes, necrosis, and aggravation tests. The control aggregate demonstrated the ordinary histological structure and no progressions were watched. Be that as it may, the faction treated with ANIT indicated huge necrotic and degenerative changes with outrageous provocative cell penetration. The ANIT + GT (75) and ANIT + VC bunches demonstrated a critical decrease in necrotic and degenerative changes with less fiery cell invasion (Figure 3). In addition, the histological changes in the ANIT + GT (75) aggregate was to some degree like the ANIT + VC faction (Figures 3C and 3D).

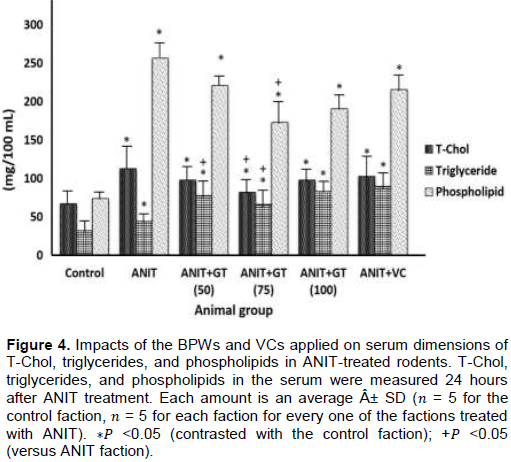
Effects of GT and VC on serum lipid profile concentrations
Serum T-Chol, triglyceride and phospholipid fixations were fundamentally lifted in the ANIT faction contrasted with the control faction (Figure 4). In the faction treated with ANIT + GT (75), an expansion in ANIT enlistment was seen in serum T-Chol, and triglyceride and phospholipid levels were altogether diminished (Figure 4). Nonetheless, in the ANIT + GT (50) faction, just ANIT enlistment with expanding serum triglyceride focus was essentially weakened. No noteworthy impact on ANIT-prompted serum T-Chol, triglyceride and phospholipid fixations were seen in the ANIT + GT (100) and ANIT + VC factions (Figure 4).
Effects of GT and VC on relative liver weight
The relative liver load of the ANIT faction was essentially higher than that of the control faction (Figure 5). In the ANIT GT (75) faction, ANIT initiated a critical decline in relative liver weight, while in ANIT GT (50), ANIT GT (100) no huge impact was seen on the relative liver weight gain and the ANIT VC faction (Figure 5).

Hepatic LPO concentrations and serum affected by GT and VC
Serum and liver LPO focuses were altogether higher in the ANIT faction than in the control rats (Figure 6). In the ANIT GT (75), ANIT GT (100) and ANIT VC factions, serum ANIT-prompted increments, and liver LPO focuses were essentially weakened, yet no critical impacts were found in the ANIT GT (50) faction (Figure 6). The decrease in serum and liver LPO focuses in the ANIT GT (75) faction was like that in the ANIT VC faction, however, essentially higher than in the ANIT GT (100) faction (P < 0.05) (Figure 6).

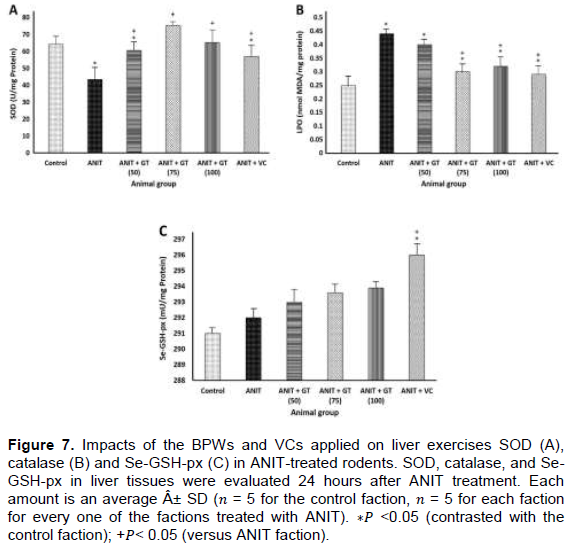
Hepatic antioxidant enzyme systems affected by GT and VC
In spite of the fact that there was no noteworthy contrast in liver CAT and Se-GSH-px movement between the two factions, the liver SOD action of the ANIT faction was considerably lower than that of the control faction (Figure 7). In the ANIT GT (50), ANIT GT (75), ANIT GT (100) and ANIT VC factions, an ANIT-instigated decrease in liver SOD action was fundamentally lessened, despite the fact that the ANIT GT (75) aggregate exhibited this. The best constriction was found in the three ANIT GTAW factions (Figure 7A). Besides, the liver SOD movement of the ANIT GT (75) or ANIT GT (100) faction did not vary from the liver SOD action of the control faction (Figure 7A). The ANIT GT (50), ANIT GT (75) and ANIT VC factions had no noteworthy impact on liver catalase action; however, the compound action of the ANIT GT (100) faction was essentially decreased (Figure 7B). The ANIT GT (50), ANIT GT (75) or ANIT GT (100) factions had no huge impact on liver Se-GSH-px movement, yet the ANIT VC amass had a noteworthy increment in this action (Figure 7C).
Hepatic antioxidant concentrations affected by GT and VC
The glutathione fixation in the ANIT faction was considerably higher than that in the control rats, in any case, there was no huge contrast in liver α-Toc focus (Figure 8). In the ANIT GT (50), ANIT GT (75) or ANIT GT (100) factions, ANIT-prompted increments in liver GSH fixations were generously diminished, while the ANIT VC mass had no critical impact on liver GSH focuses (Figure 8A). The ANIT GT (50), ANIT GT (75) and ANIT GT (100) factions had no noteworthy impact on liver α-Toc fixations; however, the liver α-Toc focuses were essentially and altogether expanded in the ANIT VC faction (Figure 8B).

Effects of GT and VC on neutrophil infiltration
Liver MPO action was essentially higher in the ANIT faction than in the control faction (Figure 9). In the ANIT GT (75) and ANIT VC factions, ANIT-actuated increments in the liver MPO movement were fundamentally weakened, albeit neither the ANIT GT (50) faction nor the ANIT GT (100) aggregate had any critical impact on liver MPO action (Figure 9). The decrease in the ANIT-initiated increment in hepatic MPO action was essentially less in the ANIT GT (75) faction than in the ANIT VC faction (P < 0.05) (Figure 9). Liver MPO impacts are a lot higher in ANIT totals than in the control faction (Figure 9). In the ANIT GT (75) and ANIT VC faction, the expansion in ANIT preparation in liver MPO practice was totally constricted, in spite of the fact that the ANIT GT (50) total and the ANIT GT (100) mass did not have any perceptible impact on the extension of liver MPO. (Figure 9). In the ANIT GT (75) polymerization, the impact of ANIT-instigated increment in liver MPO action was commonly not as much as that of ANIT VC total (P < 0.05) (Figure 9).

DISCUSSION
In the GT, epigallocatechin gallate (EGCG), quercetin (Qu), caffeine (Cf), theanine, theobromine, and catechin (C) are available in the investigation. The water extraction planning of green tea (C. sinensis) was affirmed by HPLC examination, and the substance of (EGCG) and catechin (C) were the most noteworthy among the substance of the six parts examined. This outcome is predictable with past reports (Kravchenko et al., 2011; Lan-Sook et al., 2014; Lorenzo and Munekata, 2016; Fujiki et al., 2018).).
It has been reported that rats treated with ANIT (75 mg/kg, IP) have liver damage and cholestasis, as estimated by serum ALT and AST levels, hepatocyte damage file, γ-GTP, add up to bilirubin and aggregate bile acid, files of biliary cell harm and cholestasis at 24 h, yet not at 12 h of treatment (Ohta et al., 2006; Nakamura et al., 2013). In the present investigation, ANIT-treated rats were given a solitary oral portion of GT or VC 12 h after ANIT treatment, as this deferred application of GT or VC was viewed as valuable for giving the impact of the concentrate or close clinical conditions. On account of vitamins, there is cholestasis of ANIT-actuated liver harm. Changes in hepatocyte harm and serum ALT and AST action were diminished after the application of GT at a portion of 50, 75 or 100 mg/kg 12 h after ANIT treatment. Subsequently, GT can be utilized as a cell reinforcement against ANIT-initiated hepatocyte harm in rats. Be that as it may, the defensive impact of GT was higher at its portion of 75 mg/kg than at the portion of 50 or 100 mg/kg, demonstrating that the defensive impact of GT on ANIT-incited hepatocyte damage was lessened at 100 mg/kg. Rats regulated with ANIT at a rate of 12 h after treatment (250 mg/kg) diminished the expansion in serum ALT and AST action, and these impacts were like GT (75 mg/kg). This outcome is like the outcomes announced by Nakamura et al. (2013) who detailed an expansion in ALT and AST levels in VC after treatment with ANIT. The consequences of the present investigation demonstrate that ANIT causes an expansion in serum T-Chol, triglyceride and phospholipid focus 24 h after treatment, and a huge increment in serum phospholipid fixation in most treated rats, an outcome like that obtained by Yoshizumi et al. 2005; Wu et al., 2018).
Liver LPO has appeared to add to the advancement of liver harm caused by cholestasis in rats treated once with ANIT. Also, it was reported that rat treated with ANIT (75 mg/kg, IP) demonstrated a huge increment in liver and serum LPO fixations following 24 h of treatment, despite the fact that there was a huge increment in LPO focus in the liver, yet no critical increment in the liver. Serum of ANIT-treated rats was found to have occurred 12 h after treatment (Zimmerman et al., 1997; Guangjun et al., 2002; Silvia et al., 2004; Ohta et al., 2006; Rahal et al., 2014). In the present examination, a solitary oral portion of GT (75 or 100 mg/kg) regulated to ANIT following 12 h of treatment brought about a noteworthy decline in liver and serum LPO fixations, notwithstanding that the GT portion which was 75 to 100 mg/kg was increasingly viable. As detailed by Kaida et al. (2010), controlled VC (250 mg/kg) caused a huge decline in liver and serum LPO focuses 24 h after ANIT treatment.
It has been shown in rats treated with ANIT (75 mg/kg) that liver SOD action diminished following 24 h, and otherwise without following 12 h of treatment, while liver CAT and Se-GSH-px movement expanded at 12 h in spite of an expansion. CAT and Se-GSH-px movement came back to the dimension of untreated control rats following 24 h (Ohta et al., 1999, 2001;Wu et al., 2018). In the present examination, a solitary oral application of GT (50, 75 or 100 mg/kg) for ANIT treatment 12 h after ANIT treatment brought about a critical decline in liver SOD movement found following 24 h, however with GT (75 mg/kg). The most astounding impact was accomplished and the decrease in hepatic SOD action was completely reestablished to the dimension of untreated control rats. The regulated GT (50 or 75 mg/kg) had no impact on liver CAT movement discovered 24 h after ANIT treatment, whereas the application of GT (100 mg/kg) caused a huge decline in liver CAT action. All dosages of GT had no impact on liver Se-GSH-px movement discovered 24 h after ANIT treatment. Interestingly, the application of VC (250 mg/kg) to ANIT-treated rats did not influence liver CAT movement, but rather caused a huge increment in liver Se-GSH-px action, despite the fact that application of VC somewhat lessened ANIT-incited liver decrease of SOD exercises. In this manner, the oral application of GT was found to enhance the SOD related disturbed hepatic enzymatic cancer prevention agent barrier framework in ANIT-treated rats.
Rats treated once with ANIT (75 mg/kg, IP) had an expanded liver GSH focus 24 h after treatment, and liver α-Toc fixations did not change at 12 and 24 h (Ohta et al., 1999, 2001, 2006). Moreover, the liver GSH grouping of the ANIT-treated rats given GT (75 or 100 mg/kg) did not contrast from the untreated control rats. It has been recommended that liver GSH applies a causal or lenient impact in ANIT-incited rat cholestasis liver damage by shaping a reversible GSH conjugate of ANIT in hepatocytes and transporting the GSH conjugate of ANIT into bile. Separation of free ANIT and GSH was done according to Roth and Dahm (1997). As per Jean et al. (1995), an expansion in bile GSH and ANIT focuses occurred before an increment in liver GSH fixation in rats orally treated with ANIT (100 mg/kg). In this way, GT directed to ANIT-treated rats can possibly improve the discharge of GSH from liver tissue into the circulatory system, bringing about a decline in the expansion in GSH fixation in liver tissue (Basu et al., 2013).
Green tea is a drink that is famous everywhere throughout the world and has numerous pharmacological impacts, for example, anti-proliferative, anti-cancer impacts, anti-mutagenic, and is increasingly imperative for this investigation. It is known to apply a mitigating activity by hindering neutrophil penetration and the creation of ROS in actuated neutrophils (Kwon et al., 2015; Sharifzadeha et al., 2017; Torello et al., 2018; Miyata et al., 2018). It has been reported that the polyphenol segment in green tea extricates, along these lines, has a few times higher cancer prevention agent movement than vitamin C and E (Rice-evans et al., 1995; Sharifzadeha et al., 2017). Vitamin C has anticancer action in different tumor cell lines. Be that as it may, its particular system of activity stays obscure. Nonetheless, VC is known to go about as a mitigating specialist by restraining the generation of ROS in actuated neutrophils and neutrophil penetration (Alpekin et al., 1998; Bozonet et al., 2015). Penetrating neutrophils in rat liver tissue treated with ANIT have been shown to assume a key job in the improvement of ANIT-instigated cholestatic liver damage (Roth and Dahm, 1997; Kongo et al., 1999). Kongo et al. (1999) and Ohta et al. (2006) have demonstrated that neutrophil penetration in rat liver tissue treated with ANIT (75 mg/kg) is expanded 12 h after treatment, such as before the rise.
Cholestatic causes liver harm and increments further following 24 h. In the present examination, a solitary oral application of ANIT-treated rat GT (75 mg/kg) at 12 h after treatment brought about a critical decline in liver MPO movement, which is a pointer of tissue neutrophil penetration (Krawisz et al., 1984). In spite of the fact that the equivalent regulated GT (50 or 100 mg/kg) had no huge impact on the expansion in liver MPO movement, it was found at 24 h (Mullane et al., 1985). As recently detailed by Kaida et al. (2010), regulated VC (250 mg/kg) likewise created a huge lessening of the expansion in liver MPO movement, in spite of the fact that the controlled VC was more powerful than the GT applied at a quantity of 75 mg/kg. These outcomes are in great concurrence with the histological perceptions of the above hepatocytes. In this manner, the oral application of GT (75 mg/kg) was found to repress neutrophil invasion into the liver tissue of ANIT-treated rats, as on account of oral VC. Initiated neutrophils have been shown to intervene in lipid peroxidation by the generation of ROS by NADPH oxidase in cells (Zimmerman et al., 1997). It has additionally been demonstrated that MPO intercedes on lipid peroxidation within the sight of hydrogen peroxide and halide particles (Stelmaszynska et al., 1992; Han et al., 2018). Subsequently, these discoveries enable us to bring up that osmotic neutrophils in rat liver tissue treated with ANIT by the oral application of GT may work against oxidative harm related to intemperate ROS creation, potentially through mitigating within the sight of the oral VC impact. Orally controlled GT may add to its defensive impact against oxidative harm caused by neutrophils invaded in ANIT-treated rat liver by repressing the movement of MPO in penetrating neutrophils. During this investigation, GT (75 mg/kg) directed to ANIT-treated rats were shown to restrain neutrophil invasion, enabling it to deliver H2O2 into liver tissue by means of enacted NADPH oxidase, steady with different examinations (Ohta et al., 2001; Okado and Fridovich, 2001; Gottfredsen et al., 2013; Forester and Lambert, 2011).
Epigallocatechin gallate (EGCG) is available as a noteworthy segment of GT utilized in this examination. EGCG has been responsible for application of cancer prevention agent impacts by searching ROS and hindering LPO (Senthil et al., 2008; Tachibana, 2009; Lambert and Elias, 2010; Fujiki et al., 2018). Along these lines, it was recommended that the lessening impact of GT application in the expansion of LPO fixation and SOD movement in ANIT-treated rat liver might be essential because of the oxidative activity of EGCG present in the concentrate.
CONCLUSION
The results shown in this study indicated that a single oral administration of GT to rats treated once with ANIT before the onset of apparent liver damage with cholestasis protects against liver damage with cholestasis, although this protective effect of GT diminishes at its high dose. The present results also suggested that the protective effect of GT against ANIT-induced liver damage with cholestasis could be due to the antioxidant, anti-inflammatory, anti-hyperlipemic, and anti-hypertrophic actions of the extract. The orally administered GT (75 mg/kg) was found to be more effective in protecting against ANIT-induced liver damage with cholestasis than the similarly administered VC (250 mg/kg). However, further investigation is needed to clarify the exact mechanism underlining the protective effect of GT against liver damage with cholestasis in rats treated with ANIT.
CONFLICT OF INTERESTS
The author has not declared any conflict of interests.
REFERENCES
|
Ahn MR, Kumazawa S, Hamasaka T, Bang KS, Nakayama T (2004). Antioxidant activity and constituents of propolis collected in various areas of Korea. Journal of Agricultural and Food Chemistry 52(24):7286–7292. |
|
|
Alpekin N, Seçkin S, Dogru-Abbasoglu S, Koçak-Toker N, Çevikbas U, Uysai M (1998). Effect of vitamin C on glutathione and lipid peroxide levels in rats exposed to water-immersion restraint stress. Medical Science Research 26:595-597. |
|
|
Anderson RF, Fisher LJ, Hara Y, Harris T, Mak WB, Melton LD, Packer JE (2001). Green tea catechins partially protect DNA from (.) OH, radical-induced strand breaks and base damage through fast chemical repair of DNA radicals. Carcinogenesis 22(8):1189-1193. |
|
|
Basu A, Betts AM, Mulugeta A, Tong C, Newman E, Lyons TJ (2013). Green tea supplementation increases glutathione and plasma antioxidant capacity in adults with the metabolic syndrome. Nutrition Research 33(3):180-187. |
|
|
Bozonet SM, Carr AC, Pullar JM, Vissers MC (2015). Enhanced human neutrophil vitamin C status, chemotaxis, and oxidant generation following dietary supplementation with vitamin C-rich SunGold kiwifruit. Nutrients 7(4):2574-2588. |
|
|
Chang CJ, Chiu KL, Chen YL, Chang CY (2000). Separation of catechins from green tea using carbon dioxide extraction. Food Chemistry 68(1):109-113. |
|
|
Cohen G, Dembiec D, Marcus J (1970). Measurement of catalase activity in tissue extracts. Analytical Biochemistry 34(1):30-38. |
|
|
Coyle CH, Philips BJ, Morrisroe SN, Chancellor MB, Yoshimura N (2008). Antioxidant effects of green tea and its polyphenols on bladder cells. Life Sciences 83(1-2):12-18. |
|
|
Cullen JM, Faiola B, Melich DH, Peterson RA, Jordan HL, Kimbrough CL, Miller RT (2016). Acute Alpha-Naphthylisothiocyanate-induced Liver Toxicity in Germfree and Conventional Male Rats. Toxicologic Pathology 44(7):987-997. |
|
|
Davalli P, Rizzi F, Caporali A, Pellacani D, Davoli S, Bettuzzi S, Brausi M, D'Arca D (2012). Anticancer Activity of Green Tea Polyphenols in Prostate Gland. Oxidative Medicine and Cellular Longevity (2012): ID 984219, 18 p. |
|
|
Erba D, Riso P, Colombo A, Testolin G (1999). Supplementation of Jurkat T cells with green tea extract decreases oxidative damage due to iron treatment. Journal of Nutrition (129):2130-2134. |
|
|
Forester SC, Lambert JD (2011). Antioxidant effects of green tea. Molecular Nutrition and Food Research 55(6):844-854. |
|
|
Fujiki H, Watanabe T, Sueoka E, Rawangkan A, Suganuma M (2018). Cancer Prevention with Green Tea and Its Principal Constituent, EGCG: from Early Investigations to Current Focus on Human Cancer Stem Cells.Molecules and Cells 41(2):73-82. |
|
|
Golbar HM, Izawa T, Bondoc A, Wijesundera KK, Tennakoon AH, Kuwamura M, Yamate J (2017). Attenuation of alpha-naphthylisothiocyanate (ANIT)-induced biliary fibrosis by depletion of hepatic macrophages in rats. Experimental and Toxicologic Pathology 69(4):221-230. |
|
|
Gottfredsen RH, Larsen UG, Enghild JJ, Petersen SV (2013). Hydrogen peroxide induces modifications of human extracellular superoxide dismutase that results in enzyme inhibition. Redox Biology 1(1):24-31. |
|
|
Guangjun N, Yuanlin C, Baolu Z (2002). Protective effects of green tea polyphenols and their major component, (–)-epigallocatechin-3-gallate (EGCG), on 6-hydroxydopamine-induced apoptosis in PC12 cells. Redox Report 7(3):171-177. |
|
|
Han H, Xu L, Xiong K, Zhang T,Wang Z (2018).Exploration of Hepatoprotective Effect of Gentiopicroside on Alpha-Naphthylis othiocyanate-Induced Cholestatic Liver Injury in Rats by Comprehensive Proteomic and Metabolomic Signatures.Cellular Physiology and Biochemistry 49:1304-1319. |
|
|
Jean PA, Bailie MB, Roth RA (1995). 1-Naphthylisothiocyanate- induced elevation of biliary glutathione. Biochemical Pharmacology 49(2):197-202. |
|
|
Ji SJ, Han DH, Kim JH (2006). Inhibition of proliferation and induction of apoptosis by EGCG in human osteogenic sarcoma (HOS) cells Archives of Pharmacal Research 29(5):363-368. |
|
|
Kaida S, Ohta Y, Imai Y, Kawanishi M (2010). Protective effect of L-ascorbic acid against oxidative damage in the liver of rats with water-immersion restraint stress. Redox Report 15(1):11-19. |
|
|
Kamata T, Akasaka K, Ohrui H, Meguro H (1994). Fluorometric Assay of Glutathione Peroxidase Activity in Liver and Plasma with N(9-Acridinyl) maleimide. Bioscience, Biotechnology and Biochemistry 58(5):878-880. |
|
|
Kamiya Y, Ohta Y, Imai Y, Arisawa T, Nakano H (2005). A critical role of gastric mucosal ascorbic acid in the progression of acute gastric mucosal lesions induced by compound 48/80 in rats. World Journal of Gastroenterology 11(9):1324-1332. |
|
|
Kongo M, Ohta Y, Nishida K, Sasaki E, Harada N, Ishiguro I (1999). An association between lipid peroxidation and -naphthylisothiocyanate-induced liver injury in rats. Toxicology Letters 105(2):103-110. |
|
|
Kossor DC, Meunier PC, Handler JA, Sozio RS, Goldstein S (1993). Temporal relationships of changes in hepatobiliary function and morphology in rats following -naphthylisothiocyanate (ANIT) administration. Toxicology and Applied Pharmacology 119(1):108-114. |
|
|
Kravchenko LV, Trusov NV, Aksenov IV, Avren'eva LI, Guseva GV, Lashneva NV, Tutel'ian VA (2011). Effects of green tea extract and its components on antioxidant status and activities of xenobiotic metabolizing enzymes of rats. Vopr Pitan 80(2):9-15. |
|
|
Krawisz JF, Sharon P, Stenson WF (1984). Quantitative assay for acute intestinal inflammation based on myeloperoxidase activity. Assessment of inflammation in rat and hamster models. Gastroenterology 87(6):1344-1350. |
|
|
Kwon SJ, Lambert JD, Yang CS, Hong J (2015). Role of Reactive Oxygen Species from the Green Tea Catechin, (−)-Epigallocatechin-3-gallate in Growth Modulation of Intestinal Cells. Food Science and Biotechnology 24(4):1541-1548. |
|
|
Lambert JD, Elias RJ, (2010). The antioxidant and pro-oxidant activities of green tea polyphenols: a role in cancer prevention. Archives of Biochemistry and Biophysics 501(1):65-72. |
|
|
Lan-Sook L, Sang-Hee K, Young-Boong K, Young-Chan K (2014). Quantitative Analysis of Major Constituents in Green Tea with Different Plucking Periods and Their Antioxidant Activity Molecules 19(7):9173-9186. |
|
|
Lee JH, Jeong YJ, Lee SW, Kim D, Oh SJ, Lim HS, Oh HK, Kim SH, Kim WJ, Jung, JY (2010). EGCG induces apoptosis in human laryngeal epidermoid carcinoma Hep2 cells via mitochondria with the release of apoptosis-inducing factor and endonuclease G. Cancer Letters 290(1):68-75. |
|
|
Li HC, Yashiki S, Sonoda J, Lou H, Ghosh SK, Byrnes JJ, Lema C, Fujiyoshi T, Karasuyama M, Sonoda S (2000). Green tea polyphenols induce apoptosis in vitro in peripheral blood T lymphocytes of adult T-cell leukemia patients.Japanese Journal of Cancer Research 91(1):34-40. |
|
|
Lin YS, Tsai YJ, Tsay JS, Lin JK (2003). Factors affecting the levels of tea polyphenols and caffeine in tea leaves. Journal of Agricultural and Food Chemistry 51(7):1864-1873. |
|
|
Lorenzo JM, Munekata PES (2016). Phenolic compounds of green tea: Health benefits and technological application in food. Asian Pacific Journal of Tropical Biomedicine 6(8):709-719. |
|
|
McAlpine M, Ward WE (2016). Influence of Steep Time on Polyphenol Content and Antioxidant Capacity of Black, Green, Rooibos, and Herbal Teas. Beverages 2(17):13-21. |
|
|
Miyata Y, Matsuo T, Araki K, Nakamura Y, Sagara Y, Ohba K, Sakai H (2018). Anticancer Effects of Green Tea and the Underlying Molecular Mechanisms in Bladder Cancer. Medicines (Basel) 5(3):87. |
|
|
Mullane K, Kraemer R, Smith B (1985). Myeloperoxidase Activity as a Quantitative Assessment of Neutrophil Infiltration Into Ischemic Myocardium. Journal of Pharmacological Methods 14(3):157-167. |
|
|
Nakamura T, Ohta Y, Ohashi K, Ikeno K, Watanabe R, Tokunaga K, Harada N, (2013). Protective Effect of Brazilian Propolis against Liver Damage with Cholestasis in Rats Treated with -Naphthylisothiocyanate. Evidence-Based Complementary and Alternative Medicine pp. 1-14. |
|
|
Nakazato T, Ito K, Miyakawa Y, Kinjo K, Hozumi N, Ikeda Y, Kizaki M (2005). Catechin, a green tea component, rapidly induces apoptosis of myeloid leukemic cells via modulation of reactive oxygen species production in vitro and inhibits tumor growth in vivo. Haematologica 90(3):317-325. |
|
|
Naponelli V, Ramazzina I, Lenzi C, Bettuzzi S, Rizzi F (2017). Green Tea Catechins for Prostate Cancer Prevention: Present Achievements and Future Challenges. Antioxidants 6(2):26. |
|
|
Nihal M, Ahmad N, Mukhtar H, Wood GS (2005). Anti-proliferative and proapoptotic effects of (–)-epigallocatechin-3-gallate on human melanoma: Possible implications for the chemoprevention of melanoma. International Journal of Cancer 114(4):513-521. |
|
|
Noda C, He J, Takano T, Tanaka C, Kondo T, Tohyama K, Yamamura H, Tohyama Y (2007). Induction of apoptosis by epigallocatechin-3-gallate in human lymphoblastoid B cells. Biochemical and Biophysical Research Communications 362(4):951-957. |
|
|
Ohkawa H, Ohishi N, Yagi K (1979). Assay for lipid peroxides in animal tissues by the thiobarbituric acid reaction. Analytical Biochemistry, 95(2):351-358. |
|
|
Ohta Y, Kongo M, Kishikawa T (2001). Effect of melatonin on changes in hepatic antioxidant enzyme activities in rats treated with -naphthylisothiocyanate. Journal of Pineal Research 31(4):370-377. |
|
|
Ohta Y, Kongo M, Sasaki E, Harada N (1999). Change in hepatic antioxidant defense system with liver injury development in rats with a single -naphthylisothiocyanate intoxication. Toxicology 139(3):265-275. |
|
|
Ohta Y, Kongo-Nishimura M, Imai Y, Matsura T, Kitagawa A, Yamada K (2006). -Tocopherol protects against -naphthylisothiocyanate-induced hepatotoxicity in rats less effectively than melatonin. Chemico-Biological Interactions 161(2):115-124. |
|
|
Okado-Matsumoto A, Fridovich I (2001). Subcellular distribution of superoxide dismutases (SOD) in rat liver: Cu, Zn-SOD in mitochondria. Journal of Biological Chemistry 276(42):38388-38393. |
|
|
Okuda T, Ito H (2011). Tannins of Constant Structure in Medicinal and Food Plants-Hydrolyzable Tannins and Polyphenols Related to Tannins. Molecules 16(3):2191-2217. |
|
|
Plaa GL, Priestly BG (1976). Intrahepatic cholestasis induced by drugs and chemicals. Pharmacological Reviews 28(3):207-273. |
|
|
Potter JD (1997). Cancer prevention: epidemiology and experiment. Cancer Letters 114(1-2):7-9. |
|
|
Rahal A, Kumar A, Singh V, Yadav B, Tiwari R, Sandip CS, Dharma K (2014). Oxidative Stress, Prooxidants, and Antioxidants: The Interplay. BioMed Research International 2014. |
|
|
Reddy MA, Kumar BK, Boobalan G, Kasi Reddy M, Kumar CSV, Gopala Reddy A, Lakshman M (2017). Hepatoprotective Potential of Green Tea Extract against Experimental Hepatotoxicity in Rats. Indian Journal of Pharmaceutical Sciences 79(1):58-64. |
|
|
Rice-evans CA, Miller NJ, Bolwell PG, Bramley PM, Pridham JB (1995). The relative antioxidant activities of plant-derived polyphenolic flavonoids. Free. Radiation Research Society 22(4):375-383. |
|
|
Riley C, Yasmeen E, Michael DM, William G, Wendy EW (2018). Total Polyphenol Content and Antioxidant Capacity of TeaBags: Comparison of Black, Green, Red Rooibos, Chamomile and Peppermint over Different Steep Times. Beverages 4(5):1-13. |
|
|
Roth RA, Dahm LJ (1997). Neutrophil- and glutathione-mediated hepatotoxicity of - naphthylisothiocyanate.Drug Metabolism Reviews 29(1-2):153-165. |
|
|
Schierwagen C, Bylund-Fellenius AC, Lundberg C (1990). An improved method for quantification of tissue PMN accumulation measured by myeloperoxidase activity. Journal of Pharmacological Methods 23(3):179-186. |
|
|
Sedlak J, Lindsay RH (1968). Estimation of total, protein-bound, and nonprotein sulfhydryl groups in tissue with Ellman's reagent. Analytical Biochemistry 25:192-205. |
|
|
Senthil Kumaran V, Arulmathi K, Srividhya R, Kalaiselvi P (2008). Repletion of antioxidant status by EGCG and retardation of oxidative damage induced macromolecular anomalies in aged rats. Experimental Gerontology 43(3):176-183. |
|
|
Sharifzadeha M, Ranjbarb A, Hosseini A, Khanavid M (2017). The Effect of Green Tea Extract on Oxidative Stress and Spatial Learning in Streptozotocin-diabetic Rats. Iranian Journal of Pharmaceutical Research 16(1):201-209. |
|
|
Shishikura Y, Khokhar S (2005). Factors affecting the levels of catechins and caffeine in tea beverage: estimated daily intakes and antioxidant activity. Journal of the Science of Food and Agriculture 85(12):2125-2133. |
|
|
Silvia M, Orly W, Tamar A, Moussa BHY (2004). Cell signaling pathways in the neuroprotective actions of the green tea polyphenol (-)-epigallocatechin-3-gallate: implications for neurodegenerative diseases. Journal of Neurochemistry 88(6):1555-1569. |
|
|
Somia L, Cláudia M, Radja D, Djamila Z, Paulo O, Albert R, Abderrezak D, Fadila B, Samir B (2017). Protective Effect of Green Tea (Camellia sinensis (L.) Kuntze) against Prostate Cancer: From In Vitro Data to Algerian Patients. Evidence-Based Complementary and Alternative Medicine |
|
|
Sorice A, Guerriero E, Capone F, Colonna G, Castello G, Costantini S (2014). Ascorbic acid: its role in the immune system and chronic inflammation diseases. Mini-Reviews in Medicinal Chemistry 14(5):444-452. |
|
|
Stelmaszynska T, Kukovetz E, Egger G, Schaur RJ (1992). Possible involvement of myeloperoxidase in lipid peroxidation. International Journal of Biochemistry 24(1):121-128. |
|
|
Suzuki K, Ota H, Sasagawa S (1983). An assay method for myeloperoxidase in human polymorphonuclear leukocytes. Analytical Biochemistry 132(2):345-352. |
|
|
Tachibana H (2009). Molecular basis for cancer chemoprevention by green tea polyphenol EGCG. Forum of Nutrition 61:156-169. |
|
|
Torello CO, Shiraishi RN, Della Via FI, Castro TCL, Longhini AL, Santos I, Bombeiro AL, Silva CLA, Queiroz MLS, Rego EM, Saad STO (2018). Reactive oxygen species production triggers green tea-induced anti-leukemic effects on acute promyelocytic leukaemia model. Cancer Letters 1(414):116-126. |
|
|
Wu J, Li Y, Li YY, Dai Y, Li W, Zheng M, Shi Z, Shi R, Wang T, Ma B, Liu P, Yue-Ming Ma Y (2017). Huangqi Decoction Alleviates Alpha-Naphthylisothiocyanate Induced Intrahepatic Cholestasis by Reversing Disordered Bile Acid and Glutathione Homeostasis in Mice. Frontiers in pharmacology 8(938):1-14. |
|
|
Yagi K (1976). A simple fluorometric assay for lipoperoxide in blood plasma," BiochemiaMedica 15:212-216. |
|
|
Yan J, Xie G, Liang C, Hu Y, Zhao A, Huang F, Hu P, Liu P, Jia W, Wang X (2017). Herbal medicine Yinchenhaotang protects against α-naphthylisothiocyanate-induced cholestasis in rats. Scientific Reports 7(1):4211. |
|
|
Yang CS, Wang, X, Lu G,andPicinich SC (2009). Cancer prevention by tea: animal studies, molecular mechanisms, and human relevance. Nature Reviews Cancer 9(6):429-439. |
|
|
Yang J, Shin J, Seo S, Kim S, Lee S, Shin E (2018). Effects of Antioxidants in Reducing Accumulation of Fat in Hepatocyte. International journal of molecular sciences 19(9):2563. |
|
|
Yoshizumi K, Nishioka, N, Tsuji T (2005). The xanthine oxidase inhibitory activity and hypouricemia effect of the propolis in rats. Journal of the Pharmaceutical Society of Japan 125(3):315-321. |
|
|
Zhou B, Pan J, Dai F, Zhao CY, Zhang LP, Wei QY, Yang L, Zheng RL, Liu ZL (2004). Redifferentiation of human hepatoma cells induced by green tea polyphenols. Research on Chemical Intermediates 30(6):626-635. |
|
|
Zimmerman JJ, Ciesielski W, Lewandoski, J (1997). Neutrophil-mediated phospholipid peroxidation assessed by gas chromatography-mass |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


