Full Length Research Paper

ABSTRACT
There are many conflicting reports around the phenytoin (PHT)-induced genotoxic effect especially in the in-vitro studies. PHT was claimed to cause genotoxic effect by the oxidative stress of its metabolic intermediates. However, by reviewing the distribution and activity of the enzymes responsible for PHT metabolism, we found that PHT is rarely metabolized by human lymphocytes. So that, we will use isolated cultured human lymphocytes to determine which is genotoxic, PHT itself or its metabolites? PHT 60 μg/ml were added to lymphocytes before and after metabolic activation by S9. Also, this study will investigate the possible antioxidant genoprotective effects of Thymoquinone (TQ) 1 μM and Curcumin (CMN) 15 μM on the chromosomal injury induced by PHT or its metabolites. After the end of culture period, the effects of PHT on the lymphocytes were investigated by measuring levels of chromosomal aberrations (CAs); mitotic index (MI); reduced glutathione (GSH); malondialdehyde (MDA); and 8-hydroxydeoxyguanosine (8-OH-dG). Only PHT after metabolic activation caused oxidative genotoxic effects which were significantly ameliorated by TQ more than CMN. Hence, the present study is the first to record that PHT without metabolic activation in isolated human lymphocytes from non epileptic donors cause dose dependant direct toxic effect rather than genotoxic effect.
Key words: Phenytoin, thymoquinone, curcumin, genotoxic
INTRODUCTION
MATERIALS AND METHODS
RESULTS
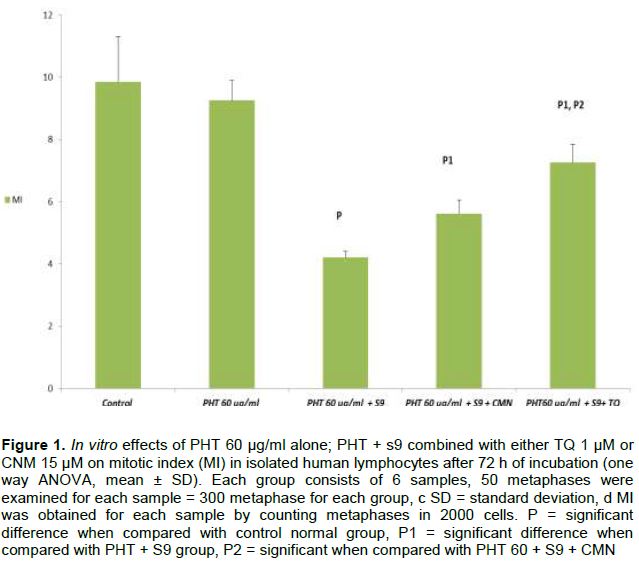
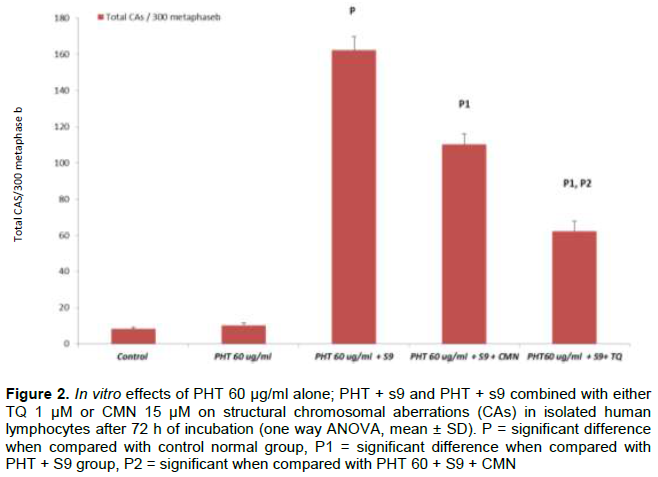
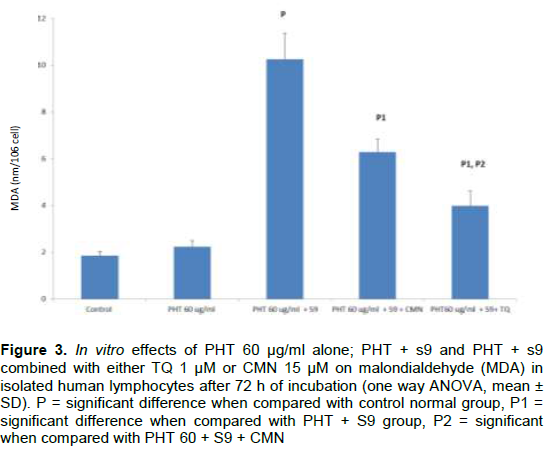
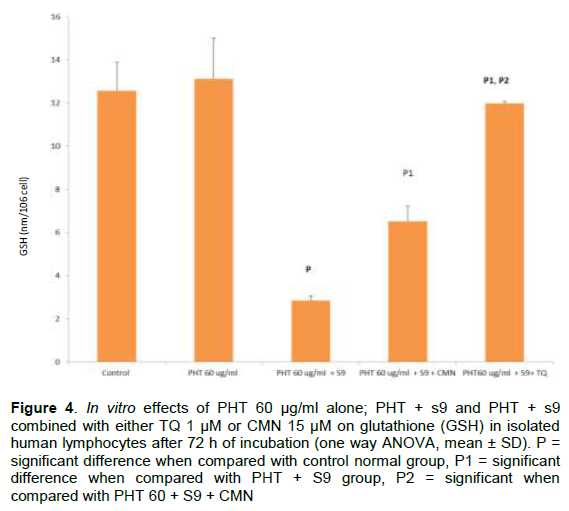
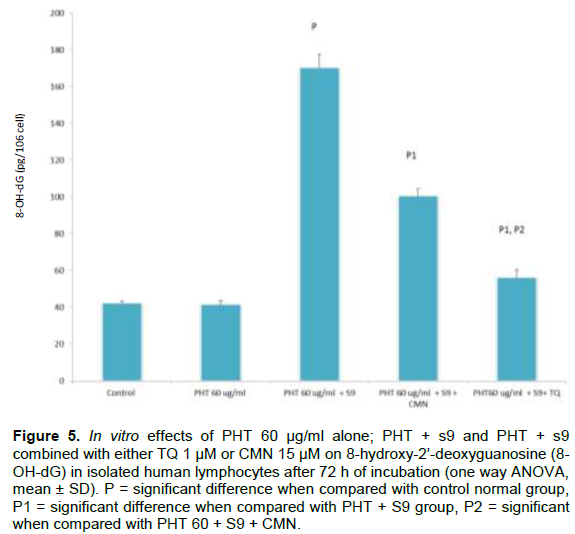
DISCUSSION
CONFLICT OF INTERESTS
REFERENCES
|
Abou Gabal AA, Essawy AE, Abdel-Moneim AM, Hame SS, Elzergy AA (2007). The protective effect of black seed (Nigella sativa) against carbon tetrachloride-induced chromosomal aberrations and ultrastructural changes of bone marrow cells. Arab J. Biotechnol. 10(2):275-288. |
|
|
Aboul-Ela EI (2002). Cytogenetic studies on Nigella sativa seeds extract and thymoquinone on mouse cells infected with schistosomiasis using karyotyping. Mutat. Res. 516(1-2):11-7. |
|
|
Al-Jassabi S, Azirun MS (2010). Phenytoin-Induced Hepatic 8-Hydroxy-deoxyguanosine in DNA of Balb/C Mice and Its Reduction by Curcumin. Mohd . Am-Eurasian J. Toxicol. Sci. 2(3):129-133. |
|
|
Anderson ME (1985). Tissue glutathione. In: Greenwald, R., A., ed. Handbook of methods for oxygen radical research. Boca Raton: CRC Press. pp. 317-323. |
|
|
Anzenbacher P, Anzenbacherová E (2001). Cytochromes P450 and metabolism of xenobiotics. Cell Mol. Life Sci. 58(5-6):737-47. |
|
|
Badary OA, Abd-Ellah MF, El-Mahdy MA, Salama SA, Hamada FM (2007). Anticlastogenic activity of thymoquinone against benzo(a)pyrene in mice. Food Chem. Toxicol. 45(1):88-92. |
|
|
Badary OA, Taha RA, Gamal el-Din AM, Abdel-Wahab MH (2003). Thymoquinone Is a Potent Superoxide Anion Scavenger. Drug Chem. Toxicol. 26:87-98. |
|
|
Bajpayee M, Pandey AK, Parmar D, Dhawan A (2005). Current Status of Short-Term Tests for Evaluation of Genotoxicity, Mutagenicity, and Carcinogenicity of Environmental Chemicals and NCEs. Toxicol. Mech. Methods 15(3):155-80. |
|
|
Basta-Kaim A, Budziszewska B, Lasoń W (2008). Effects of antiepileptic drugs on immune system. Przegl Lek. 65(11):799-802. |
|
|
Beulter E, Duron O, Kelly BM (1963). Improved method for the determination of blood glutathione. J. Lab. Clin. Med. 61:882-8. |
|
|
Calderón-Ezquerro C, Sánchez-Reyes A, Sansores RH, Villalobos-Pietrini R, Amador-Mu-oz O, Guerrero-Guerra C, Calderón-Segura ME, Uribe-Hernández R, Gómez-Arroyo S (2007). Cell proliferation kinetics and genotoxicity in lymphocytes of smokers living in Mexico City. Hum. Exp. Toxicol. 26(9):715-22. |
|
|
Carrano AV, Natarajan AT (1988). International Commission for Protection Against Environmental Mutagens and Carcinogens. ICPEMC publication no. 14. Considerations for population monitoring using cytogenetic techniques. Mutat. Res. 204(3):379-406. |
|
|
Corona-Rivera A, Urbina-Cano P, Bobadilla-Morales L, Vargas-Lares Jde J, Ramirez-Herrera MA, Mendoza-Magaua ML, Troyo-Sanroman R, Diaz-Esquivel P, Corona-Rivera JR (2007). Protective in vivo effect of curcumin on copper genotoxicity evaluated by comet and micronucleus assays. J. Appl. Genet. 48(4):389-96. |
|
|
Daba MH, Abdel-Rahman MS (1998). Hepatoprotective activity of thymoquinone in isolated rat hepatocytes. Toxicol. Lett. 95:23-29. |
|
|
Dailey LA, Imming P (1999). 12-Lipoxygenase: classification, possible therap-eutic benefits from inhibition and inhibitors. Curr. Med. Chem. 6(5):389-98. |
|
|
Draper W, Hadley M (1990). Indirect determination of oxygen free radicals. Methods Enzymol. 186:421-431. |
|
|
Durante M, Furusawa Y, Gotoh E (1998). A simple method for simultaneous interphase-metaphase chromosome analysis in biodosimetry. Int. J. Radiat. Biol. 74(4):457-62. |
|
|
El Hamss R, Idaomar M, Alonso-Moraga A, Mu-oz Serrano A (2003). Antimutagenic properties of bell and black peppers. Food Chem. Toxicol. 41(1):41-7. |
|
|
El-Sheikh AA, Morsy MA, Hamouda AH (2016). Protective Mechanisms of Thymoquinone on Methotrexate-induced Intestinal Toxicity in Rats. Pharmacogn. Mag. 12(Suppl 1):S76-81. |
|
|
EroÄŸlu HE (2011). The cytogenetic effects of black tea and green tea on cultured human lymphocytes. Braz. Arch. Biol. Technol. 54(6)1159-1165. |
|
|
Galloway SM, Armstrong MJ, Reuben C, Colman S, Brown B, Cannon C, Bloom AD, Nakamura F, Ahmed M, Duk S, Rimpo J, Margolin BH, Resnick MA, Anderson B, Zeiger E (1987). Chromosome aberrations and sister chromatid exchanges in Chinese hamster ovary cells: Evaluation of 108 chemicals. Environ. Mol. Mutagen. 10(10):1-175. |
|
|
Gökce EC, Kahveci R, Gökce A, Cemil B, Aksoy N, Sargon MF5, Kısa Ü, ErdoÄŸan B, Güvenç Y, Alagöz F, Kahveci O (2016). Neuroprotective effects of thymoquinone against spinal cord ischemia-reperfusion injury by attenuation of inflammation, oxidative stress, and apoptosis. J. Neurosurg. Spine 24(6):949-59. |
|
|
Ibrahim ZS, Alkafafy ME, Ahmed MM, Soliman MM (2016). Renoprotective effect of curcumin against the combined oxidative |
|
|
International Agency for Research on Cancer (IARC) (1996). Some Pharmaceutical Drugs; Phenytoin. IARC Monogr. Eval. Carcinog. Risks Hum. 66:175-237. |
|
|
Iqbal M, Okazaki Y, Okada S (2009). Curcumin attenuates oxidative damage in animals treated with a renal carcinogen, ferric nitrilotriacetate (Fe-NTA): implications for cancer prevention. Mol. Cell. Biochem. 324(1-2):157-64. |
|
|
Jacobsen NW, Halling-Sorensen B, Birkved FK (2008). Inhibition of human aromatase complex (CYP19) by antiepileptic drugs. Toxicol. In Vitro. 22(1):146-53 |
|
|
Kannan TP, Quah BB, Azlina A, Samsudin AR (2006). Mitotic Index and Chromosomal Analyses for Hydroxyapatite Implantation in Rabbits. Arch. Orofacial Sci. 1:15-20. |
|
|
Kaul A, Kalla NR, Goyle S (2001). The modulatory effect in genotoxic responses due to age and duration of PHT-therapy inepileptic patients. Teratog. Carcinog. Mutagen. 21(2):135-49. |
|
|
Khader M, Bresgen N, Eckl PM (2009). In vitro toxicological properties of thymoquinone. Food Chem. Toxicol. 47(1):129-33. |
|
|
Kim PM, Winn LM, Parman T, Wells PG (1997). UDP-glucuronosyl-transferasemediated protection against in vitro DNA oxidation and micronucleus formation initiated by phenytoin and its embryotoxic metabolite 5-(p-hydroxyphenyl)-5- phenylhydandoin. J. Pharmacol. Exp. Ther. 280:200-209. |
|
|
Kindig D, Garriott ML, Parton JW, Brunny JD, Beyers JE (1992). Diphenylhydantoin is not genotoxic in a battery of short-term cytogenetic assays. Teratog. Carcinog. Mutagen. 12(1):43-50. |
|
|
Kubow S, Wells PG (1989). In vitro bioactivation of phenytoin to a reactive free radical intermediate by prostaglandin synthetase, horseradish peroxidase, and thyroid peroxidase. Mol. Pharmacol. 35(4):504-11. |
|
|
Leonard A, de Meester C, Fabry L, de Saint- Georges L, Dumont P (1984). Lack o f muta- genicity of diphenylhydantoin in in vitro short-term tests. Mutat. Res. 137:79-88. |
|
|
Maclouf J, Grassi J, Pradelles P (1987). Development of enzyme-immunoassay techniques for the measurement of eicosanoids: In Walden TL Jr, Hughes HN (eds.), Prostaglandin and Lipid Metabolism in Radiation Injury, Plenum Press, Rockville. pp. 355-364. |
|
|
Malhotra A, Nair P, Dhawan DK (2012). Curcumin and resveratrol in combination modulates benzo(a)pyrene-induced genotoxicity during lung carcinogenesis. Hum Exp Toxicol. 31(12):1199-206. |
|
|
Mays DC, Pawluk LJ, Apseloff G, Davis WB, She ZW, Sagone AL, Gerber N (1995). Metabolism of phenytoin and covalent binding of reactive intermediates in activated humanneutrophils. Biochem. Pharmacol. 50(3):367-80. |
|
|
National Toxicology Program (NTP) (1993). Toxicology and Carcinogenesis Studies of 5,5-Diphenylhydantoin (CAS No. 57-41-0) (Phenytoin) in F344/N Rats and B6C3F1 Mice (Feed Studies). Natl. Toxicol. Program Technol. Rep. Ser. 404:1-303. |
|
|
Parman T, Chen G, Wells PG (1998). Free radical intermediates of phenytoin and related teratogens. Prostaglandin H synthase-catalyzed bioactivation, electron paramagnetic resonance spectrometry, and photochernical product analysis. J. Biol. Chem. 273(39):25079-88. |
|
|
Poddar S, Talukder G, Sharma A (2004). Chromosome Damage Induced by Ferric Chloride in Human Peripheral Lymphocytes. Int. J. Hum. Genet. 4(4):261-264. |
|
|
Ponnala S, Rao KP, Chaudhury JR, Ahmed J, Rama Rao B, Kanjilal S, Hasan Q, Das UN (2009). Effect of polyunsaturated fatty acids on diphenyl hydantoin-induced genetic damage in vitro and in vivo. Prostaglandins Leukot Essent Fatty Acids. 80(1):43-50. |
|
|
Poojan S, Kumar S, Verma V, Dhasmana A, Lohani M, Verma MK (2015). Disruption of Skin Stem Cell Homeostasis following Transplacental Arsenicosis; Alleviation by Combined Intake of Selenium and Curcumin. PLoS One 10(12):e0142818. |
|
|
Pradelles P, Grassi J, Maclouf J (1985). Enzyme immunoassays of eicosanoids using acetylcholinesterase as label: An alternative to radioimmunoassay. Anal. Anal. Chem. 57(7):1170-3. |
|
|
Sankar P, Telang AG, Ramya K, Vijayakaran K, Kesavan M, Sarkar SN (2014). Protective action of curcumin and nano-curcumin against arsenic-induced genotoxicity in rats in vivo. Mol. Biol. Rep. 41(11):7413-22. |
|
|
Sbrana I, Zaccaro L, Lascialfari D, Ceccherini I, Loprieno N (1984). Human lymphocytes assay: cyclophosphamide metabolic activation by S9 system with low cytotoxicity. Mutat. Res. 130(6):411-6. |
|
|
Schaumann BA, Winge VB, Pederson M, Kuskowski MA (1990). Comparative effects of phenytoin and/or phenobarbital treatment on sister chromatid exchange. Epilepsia 31:453-457. |
|
|
Sezzano P, Raimondi A, Arboix M, Pantarotto C (1982). Mutagenicity of diphenyl- hydantoin and some of its metabolites towards Salmonella typhinzurium strains. Mutat. Res. 3:219-228. |
|
|
Siddique YH, Ara G, Beg T, Gupta J, Afzal M (2010). Assessment of cell viability, lipid peroxidation and quantification of DNA fragmentation after the treatment of anticancerous drug mitomycin C and curcumin in cultured human blood lymphocytes. Exp. Toxicol. Pathol. 62(5):503-8. |
|
|
Snyder RD (2009). An Update on the Genotoxicity and Carcinogenicity of Marketed Pharmaceuticals with Reference to In Silico Predictivity. Environtl. Mol. Mutagen. 50:435-450. |
|
|
Snyder RD, Greenb JW (2001). A review of the genotoxicity of marketed pharmaceuticals. Mutat. Res. 488:151-169. |
|
|
Soga Y, Nishimura F, Ohtsuka Y, Araki H, Iwamoto Y, Naruishi H, Shiomi N, Kobayashi Y, Takashiba S, Shimizu K, Gomita Y, Oka E (2004). CYP2C polymorphisms, phenytoin metabolism and gingival overgrowth in epileptic subjects. Life Sci. 74(7):827-34. |
|
|
Stacey NH, Klaassen CD (1981). Copper toxicity in isolated rat hepatocytes. Toxicol. Appl. Pharmacol. 58:211-220. |
|
|
Tawfik SS, Abouelella AM, Shahein YE (2013). Curcumin protection activities against γ-rays-induced molecular and biochemical lesions. BMC Res. Notes 6:375. |
|
|
Tay SP, Cheong SK, Hamidah NH, Ainoon O (1998). Flow cytometric analysis of intracellular myeloperoxidase distinguishes lymphocytes, monocytes and granulocytes. Malays. J. Pathol. 20(2):91-4. |
|
|
Ugur S, Ulu R, Dogukan A, Gurel A, Yigit IP, Gozel N, Aygen B, Ilhan N (2015). The renoprotective effect of curcumin in cisplatin-induced nephrotoxicity. Ren. Fail. 37(2):332-6. |
|
|
Watson RR (1992). In in Vitro Methods of Toxicology. CRC Press. CRC Press, Boca Raton, Ann Arbor, London, Tokyo. pp. 204-216. |
|
|
Winn LM, Kim PM, Nickoloff JA (2003). Oxidative stress-induced homologous recombination as a novel mechanism for phenytoin-initiated toxicity. J. Pharmacol. Exp. Ther. 306(2):523-7. |
|
|
Witczak M, Ferenc T, Lopaczyńska D, Nowakowska D, Kociszewska I, Wilczyński J (2008). The effect of antiepileptic drugs administered in pregnancy on micronucleus frequency in cordblood lymphocytes. Int. J. Occup. Med. Environ. Health 21(1):67-71. |
|
|
Woodruff RC, Mason JM, Valencia R, Zimmering S (1985). Chemical mutagenesis testing in Drosophila. V. Results o f 53 coded compounds tested for the National Toxicology Program. Environ. Mutagen. 7(5):677-702. |
|
|
Zegura B, Lah TT, Filipic M (2004). The role of reactive oxygen species in microcystin-LR-induced DNA damage. Toxicology 200(1):59-68. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


