Full Length Research Paper

ABSTRACT
Multidrug resistance Plasmodium falciparum remains a significant global health problem worldwide. New alternative antimalarial drugs are urgently needed. Dioscorea membranacea Pierre. is a Thai-medicinal plant that has been shown to exhibit a wide range of pharmacological activities. The study aimed to investigate antimalarial activity and possible protein targets of action of the crude ethanolic extract of the rhizome of this plant. The in vitro antimalarial activity expressed as IC50 (concentration that inhibits the parasite growth by 50%) of the extract against 3D7 chloroquine-sensitive P. falciparum and K1 chloroquine-resistant P. falciparum clones were 10.1 (8.8-10.3) and 9.3 (9.17-9.63) µg/ml [median (range)], respectively. The cytotoxicity of against the human fibroblast cell OUMS-36T-1F was 96.4 (96.3-96.5) µg/ml. The selectivity index (SI) for the 3D7 and K1 clones was 9.5 and 10.4, respectively. Preliminary investigation of the protein targets of action in 3D7 P. falciparum clone revealed 13 up-regulated protein spots and 14 down-regulated protein spots. For further development of D. membranacea Pierre. as a promising antimalarial drug candidate, identification of these proteins by mass spectrometry and investigation of their mode of antimalarial actions are encouraged.
Key words: Malaria, proteomics, Dioscorea membranacea Pierre.
INTRODUCTION
Malaria is a vector-borne disease that remains a significant public health problem in tropical and subtropical regions of the world (World Health Organization, 2017). The emergence and spread of multidrug-resistant Plasmodium falciparum have become problematic for controlling malaria in most endemic regions of the world including Thailand. Nevertheless, antimalarial chemotherapy remains the mainstay for controlling malaria in the absence of effective vaccines. There is a pressing need for ongoing drug discovery research that will provide safe, effective, and affordable antimalarial agents. Several approaches have been applied for searching for new antimalarial drugs. Among these is investigation of natural - product-derived compounds for their potential antimalarial activity. Dioscorea membranacea Pierre. locally known in Thai as ‘Hua-Kao-Yen-Tai’, has long been used as a common ingredient in several Thai medicinal preparations including those for treatment of dermopathy, lymphopathy, inflammation, cancers, neural diseases, and leprosy (Itharat, 2010). The antimalarial activity of this plant has recently been reported by Thiengsusuk et al. (2013). The crude extract of the plant rhizome showed potent antimalarial activity with median IC50 (the concentrations that inhibit the parasite growth by 50%) values of less than 10 μg/ml against both K1 (chloroquine resistant) and 3D7 (chloroquine sensitive) P. falciparum clones. The objective of the present study was to confirm the antimalarial activity and identify possible protein targets of antimalarial action of the crude ethanolic extract of D. membranacea Pierre. (rhizome) using proteomics approach.
MATERIALS AND METHODS
Chemicals and reagents
RPMI, HEPES, and gentamicin were supplied by Gibco BRL Life Technologies (Grand Island, NY, USA). Chloroquine, SYBR Green I and 3-(4,5-dimethyl-2 thiazoyl)-2,5-diphenyl-2H-tetrazolium bromide (MTT) were purchased from Sigma-Aldrich Inc. (St. Louis, MO, USA). Ethanol was purchased from Labscan Co. Ltd. (Bangkok, Thailand).
Crude extract of D. membranacea Pierre. (rhizome), chemicals, and reagents
The crude extract of D. membranacea Pierre. was prepared by maceration method (Itharat et al., 2003). In brief, the plant rhizome was washed, cut into small pieces, air-dried, weighed, and ground into powder. The powder (100 g) was soaked in 300 ml of absolute ethanol for 3 days (1:3 w/v ratio), and the extract was filtered and evaporated under reduced pressure by rotary evaporation. The extract yield (2.93%) was weighed and stored at -20°C until it was used.
Assessment of in vitro antimalarial activity of the crude ethanolic extract of D. membranacea Pierre.
Two P. falciparum clones, that is, 3D7 and K1 were used in the study. The parasites were cultured according to the traditional method of Trager and Jensen (1976). Both were maintained in continuous culture in O+ human erythrocytes suspended in RPMI culture medium supplemented with 10% human B serum and 25 mM HEPES (at 37°C under 5% CO2, 5% O2, and 90% N2 atmosphere). The parasites were synchronized to ring stage P. falciparum using 5% sorbitol. Antimalarial activity of the crude ethanolic extract of D. membranacea Pierre. (rhizome) was assessed using SYBR Green I assay (Bennett et al., 2004a; Smilkstein et al., 2004). Highly synchronous ring stage parasite was used in each assay. An aliquot of parasite inoculum (50 ml) with 2% parasitemia and 1% hematocrit was added into each well of a 96-well microtiter plate. The 96-well drug plates were dosed with the extract at eight final concentrations as follows: 1.5625, 3.125, 6.25, 12.5, 25, 50, 100, and 200 mg/ml. The experiment was done three times in triplicate each. Data are presented as median (range) values. The IC50 values (concentrations that inhibit the parasite growth by 50%) used as indicators of antimalarial activity were determined from log-dose-response curves using the CalcusynTM version 1.1 (BioSoft, Cambridge, UK).
Assessment of cytotoxicity of the crude ethanolic extract of D. membranacea Pierre.
The normal human embryo fibroblast cell OUMS-36T-1F was purchased from Japanese Collection of Research Bioresources (JCRB) cell bank, Japan. The cell was cultured in RPMI 1640 medium (Gibco Co. Ltd., NY, USA) supplemented with 10% (v/v) heated fetal bovine serum (FBS) and 100 IU/ml of antibiotic-antimycotic solution (Gibco Co. Ltd., NY, USA). The cytotoxicity of the extract was determined by MTT assay (Mosmann, 1983). Briefly, the OUMS-36T-1F cell was seeded onto each well of the 96-well microtiter plate (10,000 cells/well) and incubated for 24 h (37°C, 5% CO2 atmosphere, and 95% humidity) before exposure to various concentrations of the extract (250, 125, 62.5, 31.25, 15.6, 7.8, 3.9, and 1.95 μg/ml). Following the 48 h incubation, the MTT reagent (20 μl of 5 mg/ml solution; Sigma Co. Ltd., MO, USA) was added into each well, and the plate was further incubated for 4 h. The culture medium of each well was discarded, and DMSO (100 μl) was added, and the plate was incubated at 25°C in the darkroom for 15 min. The absorbance was measured at 570 nm (Varioscan Flash, Thermo, Finland). Cell viability and corresponding IC50 was determined using CalcuSyn™ v2.11 software (Biosoft, Cambridge, UK). The experiment was done three times, in triplicate each. Data are presented as median (range) values. The selectivity index (SI) was defined as the ratio of IC50 of the crude ethanolic extract of D. membranacea Pierre. against malaria in the OUMS-36T-1F cell.
Morphological change of parasite cells following exposure to the crude ethanolic extract of D. membranacea Pierre.
Synchronized 3D7 P. falciparum was used in the experiment. The parasite was exposed to the crude ethanolic extract of D. membranacea Pierre. (rhizome) at the IC50 at 37°C under 5% CO2, 5% O2, and 90% N2 atmosphere for 48 h. Blood films were prepared at the following time points: 4, 8, 12, 16, 20, 24, 28, 32, 36, 40, and 48 h and stained with Giemsa (Biotechnical Thai, Bangkok, Thailand). Parasite cell morphology was observed under a light microscope (x100, Olympus, Tokyo, Japan).
Extraction of P. falciparum proteins following exposure to the ethanolic extract of D. membranacea Pierre.
Synchronized 3D7 P. falciparum was exposed to the ethanolic extract of D. membranacea Pierre. at the IC50 for 24 h. Parasite culture was harvested and the cell pellet was resuspended in 0.15% saponin and incubated on ice for 1 h to lyse red blood cells. The lysate was collected through centrifugation at 13,000 × g for 5 min (4°C) and washed three times with 1 ml of 50 mM Tris pH 7.5 until the supernatant was clear. The parasite pellet was re-suspended in 500 ml of rehydration buffer (8 M urea, 2 M thiourea, 2% CHAPS, 65 mM DTT, 1% ampholyte pH 3-10, and 1x of proteinase inhibitor). The sample was vortexed and sonicated on ice four times, 6 s each (21% amplitude, 6 s, interspersed with 9 s), followed by centrifugation at 13,000 × g for 1 h (4°C). Protein concentration was measured using Bradford reagent (BioRad Co. Ltd., California, USA) and the supernatant was subjected to analysis by 2-dimensional gel electrophoresis (2-DE). The experiment was done four times each.
2-Dimensional gel electrophoresis
The extract of the parasite protein (100 mg) was mixed with rehydration buffer (8 M urea, 1%CHAP, 15 mM dithiothreitol, and 0.001% bromophenol blue) to prepare protein mixture (125 ml) and applied onto 7 cm IPG strips (non-linear) with a pH range of 4 to 7 in an isoelectric focusing (IEF) system (PROTEAN® i12™ IEF Cell, BioRad Co. Ltd., California, USA). The IEF was initially performed at 250 v for 15 min, followed by 4,000 v for 1 h, and 4000 to 20,000 v-h. The focused strips were equilibrated in equilibration solution I (10 ml of 50 mM Tris-HCl, pH 8.8, 6 M urea, 30% glycerol, and 2% SDS) containing the reducing agent DTT (100 mg) for 10 min, followed by equilibration solution II (5 ml) containing iodoacetamide (450 mg) for additional 10 min. Finally, the strips were equilibrated with 1x electrode buffer (pH 8.3) for 10 min and loaded onto 12% SDS-polyacrylamide gel electrophoresis (SDS-PAGE) for second dimension separation. The gels were run on 1x electrode buffer (pH 8.3). The gels were fixed and stained with silver stain (BioRad Co. Ltd., California, USA) according to the manufacturer's recommendation. The 2-DE gel images were scanned and analyzed using PDQuestTM software (BioRad Co. Ltd., California, USA). At least four independent gels were analyzed for each sample group.
RESULTS
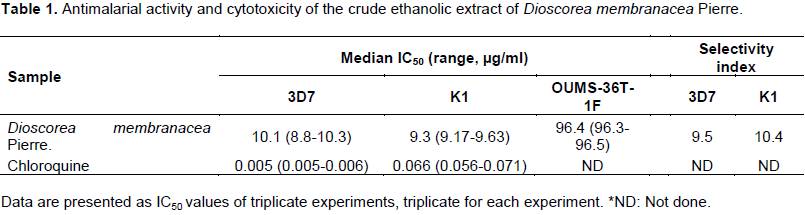
The median (range) IC50 values of the ethanolic extract of D. membranacea Pierre. for the 3D7 and K1 P. falciparum clones were 10.1 (8.8-10.3) and 9.3 (9.17-9.63) mg/ml, respectively. The cytotoxicity assay was performed using human fibroblast cell line (OUMS-36T-1F) to evaluate the selectivity of antimalarial activity. The median (range) IC50 for cytotoxicity result of the extract was 96.4 (96.3-96.5) mg/ml. The selectivity index of the extract for 3D7 and K1 P. falciparum was 9.5 and 10.4, respectively (Table 1).
The morphological change of 3D7 P. falciparum following exposure to the ethanolic extract of D. membranacea Pierre. at the IC50 was observed during the period of 4 to 48 h (Figure 1). During 4 to 12 h, the morphology of the exposed parasites was similar to the control parasite (exposed to 50% ethanol). A ring-like cytoplasm enclosing a vacuole with a dot of round nuclear chromatin projecting from the cytoplasm was observed. The stippling of Maurer's dots was found, and the cytoplasm of the parasite was thickening. The trophozoite stage showed approximately half the diameter (0.8 to 1.1 μm) of the infected red cell. During the period of 16 to 24 h, the control parasite developed to late and mature trophozoite stages. Moreover, the nuclear chromatin mass was more conspicuous but remained undivided. Following exposure to the extract on the other hand, a marked change in morphology was observed in about 50% of the parasite. The parasite was slowly grown, and the cytoplasm was shrink and dense as compared to control. This alteration was more prominent after 24 h of exposure; the trophozoites appeared relatively smaller as compared to the control with the diameter being reduced to approximately one fourth to one third. During the period of 28 to 48 h, the control parasite developed to early and mature schizont stages. The nuclear chromatin was divided and developed to 24 to 28 merozoites. Thereafter, the mature schizonts began to rupture and re-infected new red blood cells. The growth of about 50% of the exposed parasites was however arrested in the trophozoite stage and finally died.

Based on the morphological changes observed with the 3D7 P. falciparum clone following exposure to the extract, suitable time for proteomics analysis was 24 h. The protein patterns of non-exposed and extract exposed parasites are as shown in Figures 2 and 3, respectively. The protein spots were observed following separation with IPG at a pH range of 4 to 7. A total number of approximately 83 and 257 protein spots were separated throughout the gel from the control and exposed parasites, respectively. The patterns of protein spots separated from the control and exposed parasites were found to be similar, of which only 56 protein spots were likely to be the same essential proteins. On the other hand, 27 spots from control and 201 spots from exposed parasite were unmatched spots (Figure 4). The increase in protein expression of about 2- to 38-fold when compared with the control was observed in a total of 13 protein spots extracted from the exposed parasites (Table 2). The decrease in the expression to about 2 to 47% (ratio of exposed/control range from 0.002 to 0.47) of the exposed parasite was observed with about 14 protein spots (Table 3).



DISCUSSION
The antimalarial activity of the crude ethanolic extract of D. membranacea Pierre. rhizome was evaluated in 3D7 chloroquine-sensitive and K1 chloroquine-resistant P. falciparum clones in vitro. Results (IC50 values of 10.1 and 9.3 mg/ml against 3D7 and K1 clones, respectively) are in agreement with that previously reported (IC50 values of 6.2 and 5.1 mg/ml against 3D7 and K1 clones, respectively) (Thiengsusuk et al., 2013). According to the criteria proposed by Rasoanaivo et al. (2004), the IC50 of potential antimalarials from the natural products should be at least 10 mg/ml. The results confirm potent antimalarial activity of ethanolic extract of D. membranacea Pierre against P. falciparum. The selectivity of the extract against both parasite clones as compared to human normal cell was relatively high (about 10 times). In the previous study, SI of the extract was reported to be 24.2 and 29.4 for 3D7 and K1, respectively. The discrepancy could be due to the difference in sensitivity of the parasite clones to different human cell lines. Human renal epithelium cell was used in the previous study, whereas fibroblast cell line was used in the present study (Thiengsusuk et al., 2013). To further investigate for the time- and stage-specific antimalarial action, the morphological changes of 3D7 P. falciparum exposed to the ethanolic extract of D. membranacea Pierre. were observed in comparison with the control (exposed to 50% ethanol) parasite during the period of 4 to 48 h. The difference in morphology was observed in the exposed parasite starting from 16 h and was seen at 24 h. This indicates that the plant was likely to act on the growing trophozoite stage (12 to 24 h).
More than 40 Thai medicinal plants have been screened for antimalarial activity (Gale et al., 2007; Pinmai et al., 2010; Thiengsusuk et al., 2013). The promising plants include Phyllanthus emblica, Terminalia chebula, Terminalia bellerica (Pinmai et al., 2010), Plumbago indica Linn. (root), Garcinia mangostana Linn. (pericarp), Dracaena loureiri Gagnep. (stem), Piper chaba Hunt. (fruit), Myristica fragrans Houtt. (seed), Kaempferia galangal Linn. (leave), Artemisia annua Linn. (rhizome), and D. membranacea Pierre. (rhizome) (Thiengsusuk et al., 2013). The potency of antimalarial activity (IC50) of these plants range from 3 to 15 µg/ml. Maceration was applied as the method for extraction of all plants but with different solvents. In the study reported by Pinmai et al. (2010), water was used as the extraction solvent, whereas for those reported by Gale et al. (2007) and Thiengsusuk et al., (2013), ethanol was used as the extraction solvent. The selection of solvents in all studies follows the practical use in traditional medicine.
Apart from antimalarial activity, the ethanolic extract of D. membranacea Pierre. rhizome has been report to exhibit a wide range of pharmacological activities such as cytotoxic against various cancer cell lines (COR-L23, LS-174T, MCF-7, and SVK-14) (Itharat et al., 2003), anti-HIV-1 protease- and HIV-1 integrase activities (Tewtrakul et al., 2006), immunomodulatory activity (Panthong et al., 2014; Tewtrakul and Itharat, 2006). Dioscorealides, dioscoreanone discorealide B, dihydrophenanthrene are bioactive compounds isolated from its rhizome, which were responsible for these pharmacological activities (Thongdeeying et al., 2016; Itharat and Hiansai, 2012; Saekoo et al., 2010).
The proteomics analysis is one of the commonly used approaches for protein identification that has been applied in various types of research. These include the identification of biomarkers of disease pathogenesis, protein targets of action of new drug candidates, protein targets of drug resistance, and vaccine development (Cooper and Carucci, 2004; Moorthy et al., 2004; Singh et al., 2009). Proteomics is a large-scale study of proteome which includes their structures and functions. These proteins are responsible for the phenotype of cells responded to the environment and therefore, is much more complicated than the genomics approach due to a marked difference in protein types and quantity from cell to cell over time. In addition, the obstacles of proteomics research in malaria are difficulty in protein extraction and protein solubility (Rungsihirunrat et al., 2012). The commonly used technique to study proteomics is the two-dimensional gel electrophoresis (2-DE), which separates solubilized proteins in the first dimension according to their charges by using isoelectric focusing, followed by the second dimension which separates proteins according to their molecular weight (Bernard et al., 2004b; Gevaert and Vandekerckhove, 2000). In this study, insolubility of malarial membrane proteins resulted in low efficiency of separation of the protein spots. It is noted that the proteomics was performed only in the 3D7 P. falciparum clone as the change can be observed in the parasite carrying natural gene (wild-type). This ensures that the change that occurred was the effect of the extract instead of the mutation of parasite gene. The 2DE was used to analyze the expression and patterns of proteins separated from 3D7 P. falciparum clone following exposure to the extract for 24 h in comparison with the control parasite. The efficiency of protein separation was improved by cleaning up the insoluble parasite proteins by centrifugation parasite suspension (4°C) at 11,300 × g for 1 h. Abundant malaria proteins were separated at the pH gradients ranging from 4 to 7. Significantly higher number of protein spots (201 spots) was separated from the exposed as compared to the non-exposed parasite. This suggests that only a few or none of the protein was expressed during normal condition but was over-expressed following exposure of the parasite to the extract. Among the protein spots separated from parasite exposed to the plant extract, 27 protein spots were unmatched protein spots, of which 13 and 14 spots showed up- and down-regulation of expression, respectively (at least 2-fold difference in expression when compared with the control). Unfortunately the protein identification was not successful, these proteins showed hypothetical proteins with unknown function. The previous report of proteomics study from ethanolic extracts of G. mangostana against P. falciparum had been reported. The IPG strip in this study was in a wider range (pH 3 to 10) than the present study (IPG strip pH 4 to 7). This difference was due to the result of primary proteomics work, most protein spots of our study were localized at pH 4 to 7. Therefore, we designed to use the IPG strip pH 4 to 7. In this present work, only 3D7 P. falciparum clone was used in proteomic study, because the native genetic background of this clone will not interfere with the changed of protein pattern.
CONCLUSION
For further development of D. membranacea Pierre. as a promising antimalarial drug candidate, identification of these proteins by mass spectrometry and investigation of their mode of antimalarial actions are encouraged.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests
REFERENCES
|
Bennett TN, Paguio M, Gligorijevic B, Seudieu C, Kosar AD, Davidson E, Roepe PD (2004a). Novel, rapid, and inexpensive cell-based quantification of antimalarial drug efficacy. Antimicrobial Agents and Chemotherapy 48(5):1807-1810. |
|
|
Bernard KR, Jonscher KR, Resing KA, Ahn NG (2004b). Methods in functional proteomics: two-dimensional polyacrylamide gel electrophoresis with immobilized pH gradients, in-gel digestion and identification of proteins by mass spectrometry. Methods in Molecular Biology 250:263-282. |
|
|
Cooper RA, Carucci DJ (2004). Proteomic approaches to studying drug targets and resistance in Plasmodium. Current Drug Targets. Infectious Disorders 4:41-51. |
|
|
Gale GA, Kirtikara K, Pittayakhajonwut P, Sivichai S, Thebtaranonth Y, Thongpanchang C, Vichai V (2007). In search of cyclooxygenase inhibitors, anti-Mycobacterium tuberculosis and anti-malarial drugs from Thai flora and microbes. Pharmacology and Therapeutics 115(3):307-351. |
|
|
Gevaert K, Vandekerckhove J (2000). Protein identification methods in proteomics. Electrophoresis 21(6):1145-1154. |
|
|
Itharat A (2010). Comparative biological activities of five Thai medicinal plants called Hua-Khao-Yen. Thai Journal of Pharmacology 32(1):327-331. |
|
|
Itharat A, Hiransai P (2012). Dioscoreanone suppresses LPS-induced nitric oxide production and inflammatory cytokine expression in RAW 264.7 macrophages by NF-κB and ERK1/2 signaling transduction. Journal of Cellular Biochemistry 113(11):3427-3435. |
|
|
Itharat A, Plubrukarn A, Kongsaeree P, Bui T, Keawpradub N, Houghton PJ (2003). Dioscorealides and dioscoreanone, novel cytotoxic naphthofuranoxepins, and 1,4-phenanthraquinone from Dioscorea membranacea Pierre. Organic Letter 5(16):2879-2882. |
|
|
Moorthy VS, Good MF, Hill AV (2004). Malaria vaccine developments. Lancet 363:150-156. |
|
|
Mosmann T (1983). Rapid colorimetric assay for cellular growth and survival: application to proliferation and cytotoxicity assays. Journal of Immunological Methods 65:55-63. |
|
|
Panthong S, Ruangnoo S, Thongdeeying P, Sriwanthana B, Itharat A (2014). Immunomodulatory activity of Dioscorea membranacea Pierre rhizomes and of its main active constituent Dioscorealide B. BMC Complementary and Alternative Medicine 14:403. |
|
|
Pinmai K, Hiriote W, Soonthornchareonnon N, Jongsakul K, Sireeratawong S, Tor-Udom S (2010). In vitro and in vivo antiplasmodial activity and cytotoxicity of water extracts of Phyllanthus emblica, Terminalia chebula, and Terminalia bellerica. Journal of the Medical Association of Thailand 93(Suppl 7):S120-6. |
|
|
Rasoanaivo P, Deharo E, Ratsimamanga-Urverg S, Frappier F (2004). Guidelines for the Nonclinical Evaluation of the Efï¬cacy of Traditional Antimalarials. In: Traditional Medicinal Plants and Malaria, CRC Press. |
|
|
Rungsihirunrat K, Chaijaroenkul W, Siripoon N, Seugorn A, Thaithong S, Na-Bangchang K (2012). Comparison of protein patterns between Plasmodium falciparum mutant clone T9/94-M1-1(b3) induced by pyrimethamine and the original parent clone T9/94. Asian Pacific Journal of Tropical Biomedicine 2:66-69. |
|
|
Saekoo J, Dechsukum C, Graidist P, Itharat A (2010). Cytotoxic effect and its mechanism of dioscorealide B from Dioscorea membranacea against breast cancer cells. Journal of the Medical Association of Thailand 93 (12):S277-82. |
|
|
Singh M, Mukherjee P, Narayanasamy K, Arora R, Sen SD, Gupta S, Natarajan K, Malhotra P (2009). Proteome analysis of Plasmodium falciparum extracellular secretory antigens at asexual blood stages reveals a cohort of proteins with possible roles in immune modulation and signaling. The Molecular and Cellular Proteomics 8(9):2102-2118. |
|
|
Smilkstein M, Sriwilaijaroen N, Kelly JX, Wilairat P, Riscoe M (2004). Simple and inexpensive fluorescence-based technique for high-throughput antimalarial drug screening. Antimicrobial Agents and Chemotherapy 48(5):1803-1806. |
|
|
Tewtrakul S, Itharat A (2006). Anti-allergic substances from the rhizomes of Dioscorea membranacea. Bioorganic and Medicinal Chemistry 14(24):8707-8711. |
|
|
Tewtrakul S, Itharat A, Rattanasuwan P (2006). Anti-HIV-1 protease- and HIV-1 integrase activities of Thai medicinal plants known as Hua-Khao-Yen. Journal of Ethnopharmacology 105(1-2):312-315. |
|
|
Thiengsusuk A, Chaijaroenkul W, Na-Bangchang K (2013). Antimalarial activities of medicinal plants and herbal formulations used in Thai traditional medicine. Parasitology Research 112(4):1475-1481. |
|
|
Thongdeeying P, Itharat A, Umehara K, Ruangnoo S (2016). A novel steroid and cytotoxic constituents from Dioscorea membranacea Pierre against hepatocellular carcinoma and cholangiocarcinoma cells. Journal of Ethnopharmacology 194:91-97. |
|
|
Trager W, Jensen JB (1976). Human malaria parasites in continuous culture. Science 193:673-675. |
|
|
World Health Organization (2017). World malaria report 2017 Geneva: World Health Organization. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


