Full Length Research Paper

ABSTRACT
The objective of this study was to investigate the possible effects of caffeine on the performance of juveniles of Nile tilapia. One hundred and forty four (144) juveniles of Nile Tilapia with mass, total length and initial mean height of 2.94 ± 0.16 g, 5.19 ± 0.14 cm and 1.49 ± 0.04 cm, respectively, were used. The fishes were housed in 24 plastic boxes in a closed water recirculation system, where each box contained six fishes. A completely randomized design with six treatments and four replicates was used. The treatments tested were 0.0; 0.5; 1.0; 1.5; 2.0 and 2.5 g of caffeine/3 kg of feed. The experiment lasted for 40 days. Caffeine levels, up to and including 1 g/3 kg of diet, positively affected weight, total length, standard length, height, weight gain, feed intake and specific growth rate. Dosages higher than 1 g/3 kg resulted in negative results. Caffeine can be added in up to 1 g/3 kg of feed without negatively affecting the productive performance of juvenile Nile Tilapia fingerlings.
Key words: Production index, metabolism, fish, trimethylxanthine.
INTRODUCTION
Intensive fish farming has provided the potential for raising fishes in captivity, and plans needed to promote their management, without adversely affecting the socioeconomic conditions of the activity. Ostia et al. (2018) reported that good feeding management of the animals is of great importance, since the expenses of feeding are usually high. Carvalho et al. (2012) and Souza et al. (2013) perceived that the feeding management of fish is the highest production costs, reaching 70%. With this, the use of alternative foods has gained prominence, being able to meet the nutritional needs of the animals without affecting the quality of the diet and the final product. According to Radhakrishnan et al. (2016), it is necessary to develop alternative diets that are more economical, and consequently more feasible. Veras et al. (2016) pointed out that failures in food qualitycan lead to undesired consequences, such as: hetero-geneous batch size, weight loss and nutrient absorption inefficiency, which compromise fish performance. With this, the use of alternative foods has gained prominence, and is able to meet the nutritional needs of fish without affecting the quality of the diet and the final product.
Caffeine is a bioactive substance found in many food items. Its exaggerated consumption can cause a series of disorders to the human, as well as, beneficial effects when ingested in suitable dosages (Salinas-Rios et al., 2014). Due to its diverse physiological and behavioral actions, as well as, ample availability in the market, caffeine and its various effects have been increasingly studied (Cazarim and Ueta, 2014). Based on these results, new researches on this substance as well as the dosages indicated for species according to the purpose of the study have been developed in animals (Mahboob, 2014; Silva et al., 2016; Radhakrishnan et al., 2016), with an increase in fish production that is able to meet the demand of a growing population.
Research on caffeine and its possible effects on animals, as well as on fish, are still sparse. Due to this difficulty of access to such information, studies needed to meet this need, triggering new food alternatives. The objective of this study was to analyze the possible effects of caffeine on the performance of juveniles of Nile Tilapia.
MATERIALS AND METHODS
The experiment was conducted at the Federal Institute of Education, Science and Technology of Espírito Santo (IFES)– Alegre Campus, at the Laboratory of Nutrition and Production in Ornamental Species (LNPEO), in August and September 2015.
The feed was initially ground in a hammer mill with a 0.5 mm sieve, and subsequently the caffeine dosages were added, and subsequently homogenized. After homogenization, the feed was submitted to the pelletizer with granulometry (5 mm) and then taken to the forced ventilation oven at 60°C for 24 h.
The caffeine used in the experiment was the anhydrous U.S.P. (C8H10N402) manufactured by Jilin and Labsynth Products for Laboratory LTDA, certified by Synth (ISO14001) belonging to lot 147983, with 99% purity. The animals used came from the Reproduction and Larviculture Laboratory of IFES- Alegre Campus. The experiment was carried out in a completely randomized design, with six treatments and 4 replicates, for 40 days.
Nile Tilapia juveniles used had mass, total length and initial mean height of 2.94 ± 0.16 g, 5.19 ± 0.14 cm and 1.49 ± 0.04 cm, respectively. In order to allocate the animals, a water recirculation system was used, containing 24 plastic boxes with 56 L of total volume and 45 L of useful volume each, where each one had 6 animals, totaling 144 animals used in the experiment.
Approximately 3 kg of feed was used for each treatment. The treatments used were: T0 = control; T1 = 0.5 g; T2 = 1.0 g; T3 = 1.5 g; T4 = 2.0 g and T5 = 2.5 g caffeine/3 kg. The animals were fed three times a day (07:00, 12:00 and 17:00), offered ad libitum with omnivorous ration, containing 36% crude protein.
The water quality measurements were performed every day in the afternoon period and determined by the following parameters: dissolved oxygen, temperature, conductivity, ammonia and pH. The rooms were cleaned on alternate days, counting at least three times a week.
The enclosures were wrapped with black tarpaulin to mitigate the stress of the animals (Merighe et al., 2004), as well as shelters and structures were added in each box to minimize fights and any other abnormal behavior. In addition, eight 300W heaters were also added for temperature maintenance.
The following performance indexes were evaluated: weight (W), total growth (TG), partial growth (PG), height (H), weight gain (WG), apparent feed intake (AFI), protein efficiency rate (PER), specific growth rate (SGR), and survival (S).
For the animal body measurements, they were sedated with 10% eugenol in a dosage of 2 mL/L with the aid of a pipette, and the container used for the animals contained a volume of 16 L. After sedation and biometry, the animals were placed in a second vessel, also with 16 L, with constant aeration to recover the anesthetic procedure.
Biometry was performed three times during the experimental period: at the beginning of the experiment, after 20 days and at 40 days. To perform the biometry, the animals had fasted for 24 h. The animals were weighed on the analytical balance and measured by means of an analog caliper.
All the variables of the analyses were initially submitted to the homoscedasticity test and later to the analysis of variance and polynomial regression. The statistical program used was SAEG 9.1 (2007).
RESULTS
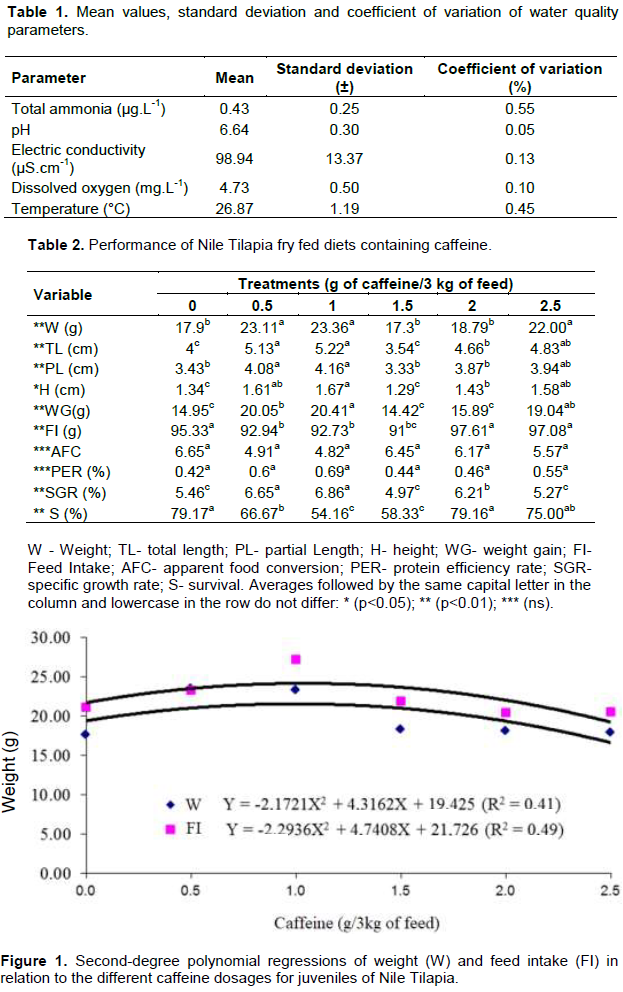
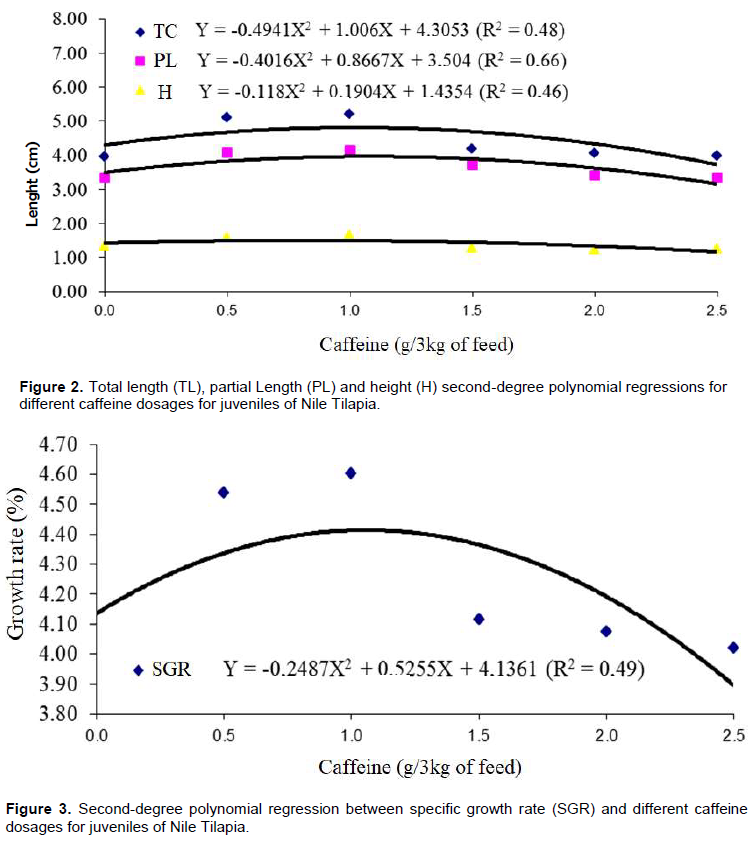
The values obtained by the physico-chemical variables of the water of the experiments are within the stipulated for tilapias (Table 1) (Salaro et al., 2006), not interfering with the obtained results. Table 2 shows the averages of the variables analyzed for the performance of Nile Tilapia fingerlings fed diets containing different levels of caffeine. The dispersion plots of the means of the treatments and the polynomial regression equations for the variables are presented in Figures 1, 2 and 3.
DISCUSSION
It can be seen that the inclusion of caffeine up to the dosage of 1 g had a positive effect on the variables analyzed. However, dosages above 1 g of caffeine had a negative effect (Lin et al., 2010; Pimenta et al., 2011). According to Gatlin et al. (2007), there is interference in feed consumption through the presence of antinutritional factors inherent in the food, as well as, caffeine (Sotelo and Alvarez, 1991), according to the dosage used. Although, the caffeine used in this research is purified, the information described corroborates with the results from inclusions above 1 g of the substance.
Braga et al. (2010), by studying Nile Tilapia (40 g), described the worst means as those derived from the diet containing cocoa meal, the latter fruit, which presents caffeine in its composition. Carvalho et al. (2012) reported lower mean values for weight gain, specific growth and protein efficiency when animals (100 g) were fed with 150 g of the cocoa meal. Pezzato et al. (1996) observed no difference (p> 0.05) for weight gain in Nile Tilapia (7g) with the inclusion of up to 20% of cocoa meal in the diet.
Other species of fish, such as catfish (Clarias mossambicus), were investigated and it was concluded that the inclusion of the substance in the diets negatively affected the growth and efficiency of feed conversion (Christensen, 1981), as well as carp (Cyprinus carpio L) (Moreau et al., 2003) and tilapia (Oreochromis aureus) (Rojas and Verreth, 2002). Fagbenro (1992) showed a decline in growth rate by replacing maize with cocoa meal by up to 45%, however, without changes in carcass quality, digestibility and feed conversion ratio of Clarias isiheriensis.
In the present study, the survival variable did not present difference (p> 0.05). It is suggested that the use of black tarpaulin and shelters added in the animal enclosures contributed positively to their survival, and it is possible that the effect of stress mitigation had a positive influence on this variable. Merighe et al. (2004) also observed a decrease in stress in tilapias by means of the black staining of the enclosures. Although not significant, the same together with the others that were significant may also have been imposed, due to the situations of agonistic confrontations, which demand energy reserves, which may have been diverted from growth and focused on the metabolic demands imposed by such confrontations. However, all these factors, when associated with the possible metabolic and lipolytic action of caffeine, triggered different responses according to the dosages used.
The results described by Bayne et al. (1976) presented no difference (p> 0.05) for weight gain. Although, the mean values of all the variables analyzed were satisfactory until the inclusion of 1 g of caffeine, it is emphasized that the substance used in the present study was 99% pure, unlike that used by Bayne et al. (1976) where caffeine was from the brown pulp. According to Caielli (1984), the chemical compositions of the pulp and the coffee husk are very similar. According to Souza et al. (2001), fiber, tannins, crude protein, among others, are components that may interfere negatively with animal performance, as already observed in ruminants (Bernardino et al., 2009; Carvalho Júnior et al., 2010). Pimenta et al. (2011) reported a high digestibility coefficient and better performance of Nile Tilapia (3 g) fed with the ration containing coffee husk, molasses, whey and formic acid, with different drying methodologies. Costa et al. (2017) studying waste of coconut biscuit residues for Nile tilapia observed that, the inclusion of biscuit residue affected significantly, the variability that can replace maize bran by up to 11.62%, improving fish performance because it affects fish metabolism and performance.
It is important to note the marked behavioral change of the individuals who received caffeine in their diets, characterizing agonistic behaviors, such as confrontations, changes in disposition in the room and high excitability. These manifestations were also in the present study as high frequency of biting, especially in the treatments with higher inclusion rates. Just as excess caffeine consumption can cause behavioral disturbances (Santos and Sant'ana, 2014), excitement, euphoria and increased motor activity in humans (Valenzuela, 2004), the same was identified in this research using juveniles of Nile Tilapia.
Behavioral changes due to substance use have also been described in other animals, such as mares (Santana, 2009) and rats (Marin et al., 2011), with excitation and increased motor activity being observed. This can be explained by the stimulating effect of the central nervous system caused by caffeine (Mello et al., 2007), where inadequate dosages may trigger psychomotor changes in animals (Holtzman et al., 1991).
The effect of caffeine on aggressiveness may vary among species, according to Nehling et al. (1992). Ingestion of psychoactive substances, such as caffeine, according to Fernández-Serrano et al. (2010) led to functional and structural changes throughout the body, mainly through interference in neurotransmitters. Caffeine is able to bind to metabotropic receptors for adenosine, further blocking the gamma-amino-butyric acid neurotransmitter (GABAA) (Meyer and Quenzer, 2005), and adenosine receptors co-located with dopamine receptors (Garett and Griffiths, 1997).
These neurotransmitters are closely linked to the central nervous system, where the binding of caffeine to its active sites can trigger various behavioral actions, as well as act on mood and food intake (Terry et al., 1995), which may have influenced consumption of animals. Changes in dopamine through caffeine may also alter the level of stress, aggressiveness (Montoya et al., 2012) and competitiveness (Arias-Carrion et al., 2010). The neurotransmitter GABA, when in interference, can also cause disturbances in food intake (Tsuji and Bray, 1991). It is suggested that caffeine dosages greater than 1 g may have triggered the described situations and caused the negative effect of the substance as the variables analyzed.
The energy balance of the animals should be considered. Vazzoler (1996) reported that fish can distribute their energy through several alternatives, directing it towards reproduction and growth. Such patterns of allocation, according to Encina and Granado-Lorencio (1997) and Huntingford et al. (2001) may vary according to habitat and food conditions, leading to changes in their physiological state. The energetic balance of the organism may undergo modifications during the life cycle of the species, as well as express the proportion of energy consumed, directed to the vital processes (Phan et al., 1993).
According to Benedito-Cecilio et al. (2005), seasonal changes in the reproductive and fish growth cycle, cause changes in the caloric content of the tissues, showing a close relationship between physiological status and body composition. Folkvord and Ottera (1993), Neu et al. (2012) and Carvalho et al. (2012) reported that the stage of development of the animals is one of the main factors that determine their feeding frequency, since young fish have a higher metabolic activity and need a greater food supply than adult animals. In addition, the intense metabolic activity of younger animals triggers less accumulation of fat, unlike the individuals in the termination phase (Lima et al., 2012).
It is inferred that higher feed consumption estimated in the present study, when compared with others, may be derived from the life stage of the animals of the present research, juveniles, which according to Kubitza (1999), direct a large part of their energy to growth. The positive effect of caffeine with the inclusion of up to 1 g in the diet can be inferred by the direct action of the substance in the central nervous system, causing an increase in metabolism (Silva et al., 2014). Thus, making the individual to present greater energy expenditure and need to feed on the higher frequency and/or breed quantity, resulting in greater food consumption, weight gain and body growth.
However, it is evidenced that even in a phase of intense metabolism and need for greater food consumption, caffeine dosages above 1 g interfered negatively with the consumption of the same, consequently causing a decrease in weight and body growth, demonstrating that caffeine can act satisfactory
or not, according to the dosage used. In dosages higher than 1 g, it is assumed that the lipolytic effect of caffeine may have occurred negatively, reducing the weight gain of the animals and interfering with their body development. One of the biggest arguments involving caffeine would be its lipolytic potential. Mello et al. (2007) described that this substance triggers an increase in the mobilization of free fatty acids in the tissue with less oxidation of carbohydrates and greater oxidation of muscle fat, once, according to Altimari et al. (2006) acts on the enzyme lipase. In addition, caffeine increases the synthesis of catecholamines, the neurotransmitters that stimulate lipolysis (Saldanha, 2012).
In humans, it is known that dosages of 3 to 9 mg caffeine/kg stimulate lipolysis; above 9 mg/kg, it no longer produce such an effect and doses of 10-15 mg/kg has toxic effects on the body (Graham et al., 2001). Studies on pigs showed positive effects when added. Chorilli et al. (2005) found that caffeine applied in the dosage of 2 mL of a solution (2%) for piglets, caused the reduction of adipose tissue thickness of the hypoderm in 55.3%. Parra et al. (2008) using coffee residue in pig feed, observed a decrease in fat in the carcass, and it is possible that this result is related to the lipolytic action of caffeine.
For other animals, as well as fish, the lipolytic effect of caffeine and its adequate dosages have not yet been stipulated; such effect is achieved in a positive manner as the decrease of fat without causing negative changes in the productive performance of the animal. It is of great economic interest that such an effect be achieved once; and excess fat in carcasses and fillets is an undesirable feature and should be maintained at levels that do not adversely affect the organoleptic characteristics of the final product, fillet yield and commercial value.
However, Spriet (1995) has described that there is no single mechanism explaining the action of caffeine, once it is able to overcome the blood-brain barrier, and the cell membranes of all tissues, making it difficult to determine specifically its action. Therefore, more studies on their potentialities in all species including fish, are needed, so that adequate dosages can be stipulated and economic, production and mainly animal welfare improvement can be provided.
CONCLUSIONS
The inclusion of caffeine in feed at doses up to 1 g/3 kg of feed improves the productive performance of juveniles of Nile Tilapia (Oreochromis niloticus).
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Arias-Carrión O, Stamelou M, Murillo-Rodriguéz E, Menéndez-González M, Pöppel E (2010). Dopaminergic reward system: a short integrative review. International Archives of Medicine 6:3-24. |
|
|
Altimari LR, Moares AC, Tirapegui J, Moreau RLM (2006). Caffeine and performance in anaerobic exercise. Brazilian Journal of Pharmaceutical Sciences 42:17-27. |
|
|
Bayne DR, Dunseth D, Ramirios CG (1976). Supplemental feeds containing coffee pulp for rearing tilapia in Central America. Aquaculture 7:133-146. |
|
|
Benedito-Cecilio E, Pereira AL, Baleroni H, Faria A (2005). Effects of habitat on physiological indicators in Leporinusfriderici (Pisces, Anostomidae) in the influence area of the Corumbá Reservoir, Goiás, Brazil. Acta Limnology Brasil, 17:71-79. |
|
|
Bernardino FS, Garcia R, Tonucci RG, Rocha FC, Valadares Filho SC, Pereira OG (2009). Nutrient intake and digestibility of elephant grass silages with coffee husks, by sheep. Revista Brasileira de Saúde e Produção Animal 10:460-469. |
|
|
Braga LGT, Rodrigues FL, Azevedo RV, Carvalho JSO, Ramos APS (2010). Apparent digestibility of energy and nutrients from agroindustrial co-products for Nile tilapia. Revista Brasileira de Saúde e Produção Animal 11:1127-1136. Caielli EL (1984). Use of coffee husks in feed for ruminants. Informe Agropecuário 119:36-38. |
|
|
Carvalho Júnior JN, Pires AJV, Veloso CM, Silva FF, Reis RA, Carvalho GGP (2010). Apparent digestibility of diet with elephantgrass ensiled with different additives. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 62(4):889-897. |
|
|
Carvalho JSO, Azevedo RV, Ramos APS, Braga LGTR (2012). Agroindustrial byproducts in diets for Nile tilapia juveniles. Revista brasileira de Zootecnia 41(12):479-484. |
|
|
Cazarim MS, Ueta J (2014). Coffee: a drink rich in substances with important clinical effects, especially caffeine. Revista de Ciências Farmacêuticas Básica e Aplicada 35:363-370. |
|
|
Chorilli M, Carvalho LS, Pires-de-Campos MSM, Leonardi GR, Ribeiro MCAP, Polacow MLO (2005). Histological evaluation of the hypodermis of pigs submitted to mesotherapeutic treatment with tiratriol, caffeine and hyaluronidase. Acta Farm Bonaerense 24:14-18. |
|
|
Christensen MS (1981). Preliminary tests on the suitability of coffee pulp in the diets of common carp (Cyprinuscarpio L.) and catfish (Clariasmossambicus Peters). Aquaculture 25:235-242. |
|
|
Costa DS, Maciel CMR, Maciel Júnior A, Magalhães Júnior FO, Smith GCB, Santos MJM, Braga LGT (2017). Waste from the biscuit and noodle industries in Nile tilapia diets. Semina: Ciências Agrárias 38:2151-2162. |
|
|
Encina L, Granado-Lorencio C (1997). Seasonal changes in condition, nutrition, gonad maturation and energy content in barbel, Barbussclateri, inhabiting a fluctuating river. Environmental Biology of Fishes 50:75-84. |
|
|
Fagbenro OA (1992). Utilization of cocoa-pond husk in low-cost diets by the clariid catfish, Clariasisheriensis(Sydenham). Aquaculture and Fisheries Management 23:175-182. |
|
|
Fernández-Serrano MJ, Pérez-García M, Río-Valle JS, Verdejo-García A (2010). Neuropsychological consequences of alcohol and drug abuse on different components of executive functions. J. Psychopharmacology 24:1317-1332. |
|
|
Folkvord A, Ottera H (1993). Effects of initial size distribution, day length, and feeding frequency on growth, survival, and cannibalism in juvelile Atlantic cod (Gadusmorhua, L.). Aquaculture 114:243-260. |
|
|
Garett BE, Griffiths R (1997). The role of dopamine in the behavioral effects of caffeine in animals and humans. Pharmacology and Chemical Behavior 57:533-541. |
|
|
Gatlin DM, Barrows FT, Brown P, Dabrowski K, Gaylord TG, Hardy RW, Herman E, Hu G, Krogdahl Å, Nelson R, Overturf K, Rust M, Sealey W, Skonberg DJ, Souza E, Stone D, Wilson R, Wurtele E (2007). Expanding the utilization of sustainable plant products in aquafeeds: a review. Aquaculture Research 38:551-579. |
|
|
Graham TE, Helge JW, Maclean DA, Kiens B, Richter EA (2001). Caffeine ingestion does not alter carbohyfrate or fat metabolism in human skeletal muscle during exercise. Journal of Physiology 26:103-119. |
|
|
Holtzman SG, Mante S, Minneman KP (1991). Role of adenosine receptors in caffeine tolerance. Journal of Pharmacology and Experimental Therapeutics 256:62-68. |
|
|
Huntingford FA, Chellappa S, Taylor AC, Strang RHC (2001). Energy reserves and reproductive investment in male three-spined sticklebacks, Gasterosteusaculeatus. Ecology of Freshwater Fish 10:111-117. |
|
|
Kubitza F (1999). Nutrition and feeding of tilapia - part 1. Aquaculture overview 9:42-50. |
|
|
Lima MR, Ludke MCMM, Holanda MCR, Pinto BWC, Ludke JV, Santos EL (2012). Performance and digestibility of Nile tilapia fed with pineapple residue bran. Acta Scientiarum. Animal Sciences 34:41-47. |
|
|
Mahboob S (2014). Replacing fish meal with a blend of alternative plant proteins and its effect on the growth performance of Catla catla and Hypophthalmichthys molitrix. Pakistan Journal of Zoology 46:747-752. |
|
|
Marin MT, Zancheta R, Paro AH, Possi APM, Cruz FC, Planeta CS (2011). Comparison of caffeine-induced locomotor activity between adolescent and adult rats. European Journal of Pharmacology 25:363-367. |
|
|
Merighe GKF, Pereira-Da-Silva EM, Negrão JA, Ribeiro S (2004). Effect of the Color of the Environment on Social Stress in Nile Tilapia (Oreochromis niloticus). Revista Brasileira de Zootecnia 33(4):828-837. |
|
|
Meyer JS, Quenzer LF (2005). Nicotine and caffeine. In: Psychopharmacology: Drugs, the brain, and behavior pp. 303-324. |
|
|
Montoya ER, Terburg D, Bos PA, Van Honk J (2012).Testosterone, cortisol, and serotonin as key regulators of social aggression: A review and theoretical perspective. Motivation and Emotion 36:65-73. |
|
|
Moreau Y, Arredondo JL, Perraud-Gaime I, Roussos S (2003). Dietary utilization of the protein and energy of the pulp of fresh coffee and silage by the tilapia of the Nile (Oreochromis niloticus). Brazilian Archives of Biology and Technology 46:35-347. |
|
|
Mello D, Kunzler DK, Farah M (2007). Caffeine and its ergogenic effect. Revista Brasileira de Nutrição Esportiva 1:30-37. |
|
|
Lin S, Mai K, Tan B, Liu W (2010). Effects of Four Vegetable Protein supplementation on Growth, Digestive Enzyme Activities, and Liver Functions of Juvenile Tilapia, Oreochromisniloticus× Oreochromis aureus. Journal of the World Aquaculture Society 41:583-593. |
|
|
Nehling A, Daval JL, Debry G (1992). Caffeine and central nervous system: mechanisms of action, biochemical, metabolic and psychostimulant effects. Brain Research Reviews 17:139-170. |
|
|
Neu DH, Furuya WM, Tamashiro D, Bittencourt F, Moro EB, Fernandes DRA, Boscolo WR, Feiden A (2012). Glycerol in the diet of Nile Tilapia fingerlings (Oreochromis niloticus). Revista Agrarian 5:288-294. |
|
|
Ostia JAS, Moraes MAB, Carmob CF, Mercante CTJ (2018). Nitrogen and phosphorus flux from the production of Nile tilapia through the application of environmental indicators. Brazilian Journal of Biology 78:25-31. |
|
|
Parra ARP, Moreira I, Furlan AC, Painao D, Scherer C, Carvalho PLO (2008). Use of coffee husks in feed for pigs in the growing and finishing stages. Revista Brasileira de Zootecnia 37:433-442. |
|
|
Phan VN, Gomes V, Morais DM, Passos MJACR (1993). Bioenergetic studies of coastal marine animals Paralonchurus brasiliensis (Perciformes, Sciaenidae). Paralonchurus Brasiliensis 10:199-215. |
|
|
Pimenta CJ, Oliveira MM, Ferreira LO, Pimenta MESG, Logato PVR, Leal RS, Murgas LDS (2011). Utilization of coffee residue in Nile Tilapia feed. Archivos de Zootecnia 60:583-593. |
|
|
Pezzato LE, Oliveira ACB, Dias E, Barros MM, Pezzato AC (1996). Weight gain and anatomopathological changes of Nile tilapia fed with cocoa meal. Pesquisa Agropecuaria Brasileira 31:375-278. |
|
|
Radhakrishnan S, Bhavan PS, Seenivasan C, Muralissankar T (2016). Impact of fishmeal replacement with Arthrospira platensis on growth performance, body composition and digestive enzyme activities of the freshwater prawn, Macrobrachium rosenbergii. Aquaculture Research 3:35-44. |
|
|
Rojas UJB, Verreth JAJ (2002). Growth, feed utilization and nutrient digestibility in tilapia fingerlings (Oreochromisaureus Steindachner) fed diets containing bacteria-treated coffee pulp. Aquaculture 33:189-195. |
|
|
Salaro AL, Okano WY, Zuanon JAS, Lambertucci DM (2006). Water quality in fish farming. Cadernos Técnicos de Veterinária e Zootecnia 50:1-8. |
|
|
Santana KS (2009) Effects of caffeine on the memory of marmosets (Callithrix jacchus). 66f. Dissertation (Master in Psychobiology) - Federal University of Rio Grande do Norte, Natal. |
|
|
Saldanha LA (2012). Effects of caffeine, coffee (Coffea arabica) and mate tea (Ilex paraquariensis) on the lipolytic activity of adipose tissue and metabolic parameters in rats submitted to physical exercise. 50f. Thesis (Nutrition in Public Health) - Faculty of Public Health (USP), São Paulo. |
|
|
Salinas-Rios T, Sánchez-Torres-Esqueda MT, Hernándes-Bautista J, Díaz-Cruz A, Nava-Cuellar C, Ortega-Cerrilla ME, Cordero-Mora JL, Vaquera-Huerta H, Velasco JLF (2014). Carcass characteristics, physicochemical changes and oxidative stress indicators of meat from sheep fed diets with coffee pulp. Arquivo Brasileiro de Medicina Veterinária e Zootecnia 66:1901-1908. |
|
|
Santos RNS, Sant'ana DMG (2014). Relationship between the use of licit drugs and memory. MUDI Archives 18:43-54. |
|
|
Silva LA, Pereira RA, Túrmina JA, Kerppers II, Altimari LR, Malfatti CRM (2014). Acute caffeine intake lowers glycemia before and after acute physical exercise in diabetic rats. Revista de Nutrição 27:143-149. |
|
|
Silva ETL, Pedreira MM, Moura GS, Pereira DKA, Otoni CJ (2016). Larval culture of nile tilapia lineages at different storage densities. Revista Caatinga 29:709-715. |
|
|
Sistema para Análises Estatísticas (SAEG) (2007). SAEG: system for statistical analysis, version 9.1. Viçosa: UFV. |
|
|
Souza RC, Melo JFB, Nogueira Filho, RM, Campeche DFB, Figueiredo RACR (2013). Influence of mango flour on growth and body composition of Nile Tilapia. Archivos de Zootecnia 62:217-225. |
|
|
Sotelo A, Alvarez RG (1991) Chemical composition of wild theobroma species and their comparison to the cacao bean. Journal of Agriculture and Food Chemistry 39:1940-1943. |
|
|
Spriet LL (1995). Caffeine and performance. International Journal of Sport, Nutrition, Exercise and Metabolism 5:84-99. |
|
|
Terry P, Gilbert DB, Cooper SJ (1995). Dopamine receptor sub-type agonists and feeding behavior. Obesity Research 3:515-523. |
|
|
Tsuji S, Bray GA (1991). GABA-related feeding control in genetically obese rats. Brain Research 540:48-54. |
|
|
Valenzuela AB (2004). The consumption and health: characteristics and beneficial properties of this millenary drink. Revista Chilena de Nutrición 31:72-82. |
|
|
Veras GC, Soares LMO, Brabo MF, Paixão DJMR, Dias BCB, Alves AX, Murgas LDS, Campelo DAV (2016). Photoperiod and feeding frequency in acarelander larviculture Pterophyllum scalare. Archivos de Zootecnia 65:581-584. |
|
|
Vazzoler AEAM (1996). Reproductive biology of teleosteos fish: theory and practice. Maringá: Eduem; São Paulo: SBI. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


