Full Length Research Paper

ABSTRACT
The study was conducted to assess the influences of edaphic and topographic variables on plant diversity on the slopes of Gra-kahsu forest area. Collection of vegetation data was made using systematic sampling methods, laying 19 transects and 62 quadrats, each with 20 m × 20 m for trees and 5 m × 5 m for shrubs. In each quadrat, heights (1.5 m), diameters (2.5cm) and numbers of woody species, level of grazing intensity, level of human disturbance and topographic variables were recorded. Analysis of one way using R-software was used to analyze the mean of plant diversity across the edaphic and topographic variables. The highest species richness and diversity appeared in the upper altitude gradients and the variation was significant (p<0.05). The nil and slightly grazed sites had significantly higher woody plant species diversity, density and basal area compared to the heavy grazed sites (p<0.001). Grazing pressure and human disturbance had an effect on the density and number of woody species as well as on the vegetation structure. This research concluded that, changes in woody plant vegetation, density and regeneration status are caused by interactions edaphic and topographic variables and these interactions determine the ecological of plant diversity.
Key words: Altitudinal pattern, disturbance, plant species diversity, slope.
INTRODUCTION
Patterns of plant species diversity are influenced by latitudinal, altitudinal and soil gradients (Huston, 1994). Understanding vegetation and species diversity patterns is fundamental for conservation of natural areas; these patterns have frequently been the focus of ecological studies (Loreau et al., 2001; Fetene et al., 2006; Muhumuza and Byarugaba, 2009). Several different patterns of plant species diversity have been noted in response to altitudinal gradients, with plant species diversity being reported to: (i) decline with higher altitude; (ii) increase with higher altitude; (iii) bulge at mid-altitude; (iv) dip at mid-altitude; or (v) have no clear relationship with altitude (Chang et al., 2005). Vegetation patterns are determined by environmental factors that exhibit heterogeneity over space and time, such as climate and topography as well as human disturbances (Alexander and Millington, 2000). Topography has climatic influence with respect to prevailing sunshine, air currents and water bodies. Altitude has a marked influence on the kind, nature and productivity of rangeland and it creates ecologically diverse vegetation (Herlocker, 1999).
In Southern Tigray, Northern Ethiopia, there are some of the studies conducted earlier (Tefera et al., 2005; Wolde et al., 2011a, b, c; Yayneshet, 2011) were specifically tried to estimate the biodiversity without considering environmental gradients like altitude and slope and edaphic variables. Some of them were only tried in comparing the natural vegetation with that of other land uses like communal grazing. However, environmental factors such as altitude and slope play significant roles in influencing species diversity (Acharya et al., 2011). Therefore, there is high demand on information about biodiversity of natural forests considering edaphic and topographic variables. Such baseline information in return will help for proper land use planning of large watershed area, considering agro ecology, topography and altitude. To minimize loss of forest fragments, understanding the ecological and anthropogenic factors involved in the process are highly important. Studying species richness patterns at different scales is very important both for ecological explanations and for effective conservation design (Feyera et al., 2014). Similarly, the forest of the study area has been exploited by surrounding communities for agricultural land expansion, timber harvesting, ï¬rewood collection, woodcutting for construction and other purposes (Leul et al., 2010).
Even though there are many natural vegetation areas in southern Tigray including forests in different agro-ecology and land use, their potential regarding species diversity have not been studied and documented. Especially regarding edaphic and topographic variation of species were not being previously assessed. These data will enable local communities and Forestry Administration to design appropriate intervention strategies for managing the remnant forest of the study area (Leul et al., 2015). Gra-Kahsu forest presents an ideal location to test the hypotheses of altitudinal patterns of plant species diversity because it integrates several distinct vertical vegetation zones and the status of the vegetation and the plant species habitats are natural. There have been a few studies on the altitudinal pattern of plant diversity on Gra-Kahsu forest (Leul et al., 2010), but a more detailed investigation on this subject had not been reported. Therefore, l investigated the effect of edaphic and topographic variables on the plant diversity distribution in Gra-Kahsu forest. The research identifies (1) the significant difference in tree species richness, structure and regeneration status between associated edaphic and topographic variables; (2) the key associated edaphic and topographic variables influencing plant species diversity.
MATERIALS AND METHODS
Description of the study Area
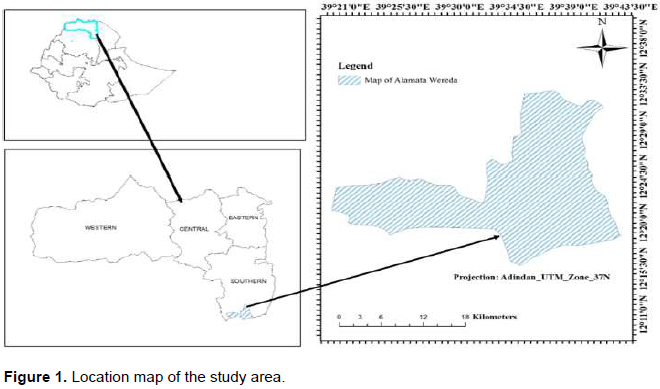
Gra-Khasu forest is located in the southern zone of Tigray at about 600 km north of Addis Ababa or some 160 km south of Mekele, the capital of Tigray Regional State. It is located between 120 22’ and 120 42’N latitude, 390 28’ and 390 40’ E longitude (Figure1) at an altitudinal distribution from 1560 m to 2688 m. The total area of the forest is more than 3500 ha. The study area, Gra-Kahsu forest was designed to conserve long lasted unique natural features, historical interests and other natural values with legal and administrative supports on the upper part of Alamata town and is endowed with different natural resources such as wildlife and other biodiversity, which contribute great potent source as important pillars for future local development. The natural vegetation coverage of the district is very small. It is the Gra-Kahsu forest, which accounts good cover of the district’s forest cover. The lowland parts of the district are dominated by Acacia species (WAOARD, 2017).
Data collection
Systematic sampling was employed for vegetation. The results were based on a complete inventory of temporary plots, stratified at altitude intervals from 1620 to 2298 m, were set up, and 2–4 quadrats were established around each sampling point in September 2017 (When most plants were in flowering stage) and species data were recorded. The quadrat size was 20 × 20 m for trees, within which 5 × 5 m quadrats were used to record shrubs. The plant height was measured using a height meter, and diameter at breast height was measured using a caliper. In each major plot, subplots (1 m2) were established at the center and corner for seedlings and saplings data. The number of seedlings (height < 1.3 m) and saplings (height > 1.3 m, diameter at breast height < 5 cm) were counted and trees (height >1.3 m, diameter at breast height ≥ 5 cm) were measured at breast height (1.3 m) by a diameter tape in each quadrat.
Altitude, slope, disturbance and grazing intensity were also recorded for each quadrat. The altitude in each quadrat was measured using a global positioning system, the slope were measured from the center of the plot using a compass meter. The state of human interference at each quadrat was estimated following Hadera (2000). A 1 to 4 subjective scale was taken into consideration to record the presence or absence of stumps, logs and signs of fuel wood collection. Therefore, the magnitude of the impact was quantified as follows:1  =  no obvious disturbance, 2  =  low disturbance, 3  =  medium disturbance, and 4  =  heavy disturbance). Grazing intensity was estimated following Kebrom et al., (1997): 0= nil; 1= slight; 2= moderate and 3= heavy. Three altitudinal gradients: lower altitude (1620-1699 masl.); middle altitude (1700-1891 masl) and upper altitude (³1892– 2298 masl). The ranges given are the minimum and maximum elevation of samples (Bruun et al., 2006). Slope location: 1, 2 and 3 for lower (0-25%), middle (26-45%) and upper (³46-65%) position at a slope respectively.
Data analysis
The following formula was used to determine density and basal area of wood species in the study sites (Mueller-Dombois and Ellenberg, 1974).
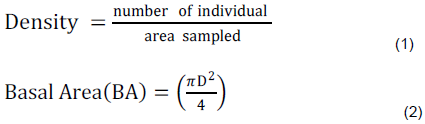
Where ''D" is diameter at breast-height in meter and π =3.14
Species diversity indices were used to calculate diversity values as follows (Shannon and Wiener, 1949):

Where; H’= Shannon diversity indices, S= the number of species, Pi=proportion of individual species and InPi=log proportion of individual species. The following formula was used to determine species evenness of woody species in the study sites (Magurran, 2004).
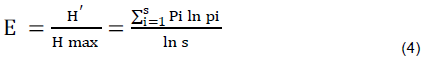
Where; H’= Shannon diversity indices, S= the number of species and Hmax = is the maximum level of diversity possible within a given population.
Regression analyses were performed to establish the relationships between species diversity and environmental variables. All statistical analyses were performed in the statistical software package R (R Development Core Team, 2015).
RESULTS AND DISCUSSION
Species diversity and evenness
The three-altitudinal gradients had significantly different value of Shannon and Weaner diversity index and species richness. Upper altitude had significantly higher Shannon and Weaner diversity index value (2.2) than the other two-altitudinal gradients (Table 1). Thus, such result with higher diversity in the upper altitude might be due to relatively favourable climatic, environmental condition and soil characteristics specially soil texture, soil organic matter (Desalegn and Carl, 2010) and slope (De Lafontaine and Houle, 2007). Altitude is a factor that determines the distribution of climatic factors and land suitability; this influences the natural vegetation types and their species diversity (Alemayehu, 2003) and influencing much of other abiotic and biotic component like temperature, soil, topography and vegetation (Otypkova et al., 2011; Tibebu and Teshome, 2015). Studies on the variations in species richness along altitudinal gradients indicate a variety of results under different biotic and abiotic condition. Most of the studies suggests that species richness decline with increasing altitude (Sharma et al., 2009). However, in most cases findings stress that species richness decrease with altitude (Bruun et al., 2006). Nevertheless, several studies have also documented a non-monotonic pattern of species richness (Vetaas, 2002). The most commonly observed pattern of diversity is a mid-altitudinal bulge (Rahbek, 2005; Zhao et al., 2005). This may arise as the upper altitudes have typical and difficult climatic condition than the lower ones, so these conditions may restrict the spread of species in the upper limit of altitude. Therefore, this will create favorable condition to sustain their fitness in the upper limit of altitude (Ole et al., 2002). Some have argued that whether species diversity will increase or decrease with increasing altitude or will peak at an intermediate altitude depends largely on specific patterns of interactions among plant communities, species, and environmental factors (Brown, 2001; Körner, 2007).
Altitude gradients are one of the most commonly determinative factors in shaping the spatial patterns of species richness (Acharya et al., 2011; Ru and Zhang, 2012; Zhang et al., 2013). The reason of high species richness, Shannon diversity index in the upper altitude might be its climatic conditions that allow many species to coexist and due to the topographical nature where upper altitude is almost steep slope made itself away from human disturbance. On the other hand, lower altitude is more prone to arable land due to gentle slope nature made to coexist less plant species. Yohannes et al. (2015) conclude that, mountain forest mostly affects by environmental variables due to change in species structure and composition.Species evenness did not show a strong response to slope gradient as there is no significant difference in its value in all the three slope gradients (P>0.05). This indicates that the distribution of species in the natural vegetation was not affected by slope. Even though it had slight variation among the three slope gradients, noticeably the upper and middle had slightly highest species richness value with 13.6 as compared with lower (11.24) (Table 1). However, there is a significant difference (P<0.001) in species richness and Shannon and Weaner diversity index between the four human disturbance and grazing intensity (Table 2). This indicates human disturbance and animal grazing intensity significantly influence species richness as well as Shannon and Weaner diversity index of an area. This idea was also in line with other similar report (Jin et al., 2013). Similarly, species richness, diversity, and evenness decreased with increasing disturbance intensity, reaching a maximum at no obvious disturbance.
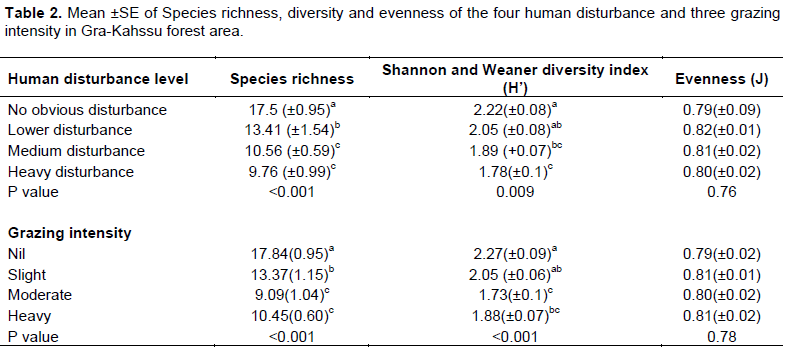
The mean of Shannon and Weaner diversity index for no obvious disturbance, low and moderate human disturbance were 2.22±0.08, 2.05±0.08 and 1.78±0.1, respectively. However, areas having heavy and moderate grazing intensity had significantly lower species richness coverage than the other two grazing intensity followed by lower grazing intensity. However, nil grazing intensity had significantly higher species richness coverage (Table 2). This result could be related to the decreasing of animal grazing pressures and human activities on plant damaged with increased distance from settlements (Brinkmann, 2009). These differences are primarily a function of differences in site productivity, habitat heterogeneity and/or disturbance factors (Maestre, 2004). For example, the low species richness in the heavy and moderate disturbance are due to anthropogenic disturbances such as burning, grazing, and wood collection, which have significantly reduced species richness (Maestre, 2004; Feyera et al., 2014). The low values of the Shannon diversity index of heavy and moderate grazing intensity also support the hypothesis of dominances of few early successional species and/or few species due to selective cutting of other species (Bone et al., 1997). Generally, the difference in the values of Shannon diversity among the studied human and grazing intensity are an indicator of disturbance level difference. Also, Gunnar and Ove (2001), Oba (2011), Tessema et al. (2011), and Angassa (2014) conclude that grazing is a key process for maintaining biodiversity.
Species richness, Shannon–Wiener index and evenness showed a quadratic decreasing pattern along the disturbance gradient, that is, the highest diversity appearing at the lowest disturbance intensity (Xiang and Zhang, 2009). Some researchers have argued that the highest diversity appeared at medium disturbance (grazing) (Pavlu et al., 2003; Smet and Ward, 2005). The maximum diversity appeared under the weakest disturbance (Jin-Tun et al., 2016). Similarly, Amsalu (2000) suggested that heavy grazing might cause reduction of plant species composition and diversity over time. Grazing is also considered essential to maintain the structure, functionality and diversity of plant communities (Hoshino et al. 2009). Similarly, mean species richness in undisturbed treatments was significantly higher than disturbed treatments (Leul et al., 2015). A parallel result was observed by Aynekulu et al. (2009) in Northern Ethiopia, demonstrating the impact of disturbance on species richness. Main factors responsible for destruction of natural forests are combination of agricultural expansion, commercial harvest, free livestock grazing, unsustainable firewood collection and charcoal production regimes (Alemayehu, 2007). Adane (2011) correspondingly observed that both mean number of species and species richness decrease along disturbance gradient.
There is a significant difference in basal area between the three altitudinal gradients (P=0.001) (Table 3). This indicates altitude significantly influence basal area of an area. This finding was also in line with other similar report (Markos and Simon, 2015). The mean basal area for lower altitude, middle altitude and upper altitude were 6.40 m2/ha, 6.48 m2/ha and 10.15 m2/ha, respectively. Therefore, upper altitude had significantly higher basal area coverage than the other two gradients followed by middle altitude. Based on the report the basal area of the lower altitude and lower slope, indicating the woody species were thin. The greater difference in basal area between the environmental variables could be due to the high number of multi-stemmed trees in the upper altitude and slope, leading to bigger diameters. This directly relational trend indicated that the value of basal area starts to grow higher as altitude increase. This might be due to the presence of relatively higher proportion of larger and aged trees in higher altitudinal classes, owing to little intervention by human activities like tree cutting, farming, grazing, etc. In addition, Markos and Simon (2015) suggest that altitude affects basal area due to its influence on light radiation, temperature, moisture, runoff and infiltration. Similarly, Kumelachew and Tamrat (2002) and Teshome et al., (2004) confirmed the results. The mean basal area increases with altitude but does show a significant trend (Carpenter, 2005).
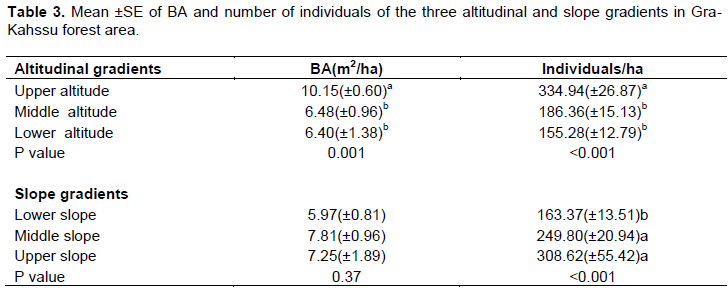
The mean basal area of trees and shrubs per hectare and number of individuals in the four human disturbance as well as grazing intensity difference was highly significant (P<0.001). Areas with low human disturbance and nil grazing intensity had higher number of individual per hectare (371.14+26.57 and 382.30+26.04), respectively (Table 4). The stands sampled in this community are located in an area with low human interference in the form of firewood collection and selective cutting and cattle is encountered. This might be due to being far from the farmer’s settlement area. However, areas with heavy human disturbance and grazing intensity had lower mean of basal area, this may be due to highly influenced by people collecting firewood, charcoal making and grazing animals. This is due to its being nearby to community and having species of plants suitable for charcoal making and firewood. This finding is dissimilarity with what was reported by Althof (2005) and Rembold (2011) where basal area is inversely related to disturbance. Higher basal area means there are large number of individuals in bigger diameter classes. However, when a given forest is heavily disturbed most of the trees with bigger diameter are removed for various reasons and result in low basal area (Adane, 2011).

Regeneration status of plants across edaphic and topographic variables
Human disturbance and grazing intensity of Gra-kahsu forest were significantly affected the regeneration status of species. Of all the edaphic factors studied, human and grazing intensity were determined the distribution of the density of mature trees, sapling and seedling identified in the study. For instance, the highest of number of mature trees, seedlings and saplings/ha appeared in no obvious human disturbance; this suggests that human disturbance was important factor for regeneration status of plant species (Table 5). Lower number of seedlings from trees might attributed to poor seeding of matured trees due to age, livestock grazing (can destroy seedlings by trampling and browsing), seed and/or fruit predation, etc. or a combination of two or more of these factors (Adane, 2011).
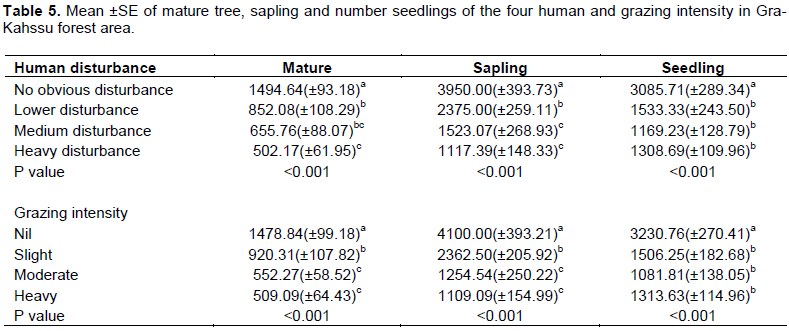
Disturbances such as intensive removal of trees for timber, construction and forest grazing have placed signiï¬cant pressure on forest regeneration (Leul et al., 2010). Leul et al. (2015) also conclude that, human intervention is high in the disturbed sites accompanied with uncontrolled grazing that affected regeneration success of the species. High density of seedlings/ saplings in the undisturbed sites is a sign of successful seed germination and establishment compared to disturbed sites (Ganesan and Siddappa, 2004). The influence of slope and altitude affects regeneration status of the vegetation. It is also similar to McDonald et al. (2010) idea, which states that the sensitivity of plants to moisture availability renders the regeneration of the dry forests. The regeneration of the forest is affected not only by environmental factors but also by anthropogenic activities (Markos and Simon, 2015). Adane (2011) conclude that the main causes of regeneration of the forest are mainly due to the geographical location of the forests and/or altitudinal difference type of disturbance (grazing and cuttings) as well as the intensity of disturbance or combination these factors. Several types of disturbances such as logging, landslides, herbivores, etc. can affect the potential regenerative status of species composing the natural vegetation stand spatially and temporally (Guarino and Scariot, 2012). Disturbance intensity directly affects plant growth, development and regeneration by trampling, bending shoots, cutting stems and grazing (Austrheim, 2002; Zhang et al., 2013; Sproull et al., 2015).
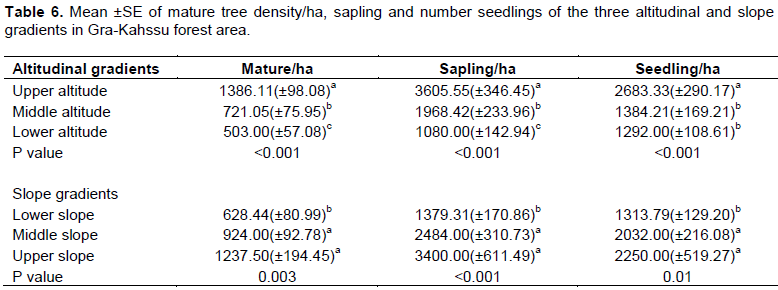
In the upper altitude, far distance was significantly higher in number of mature tree, sapling as well as seedling than lower and middle altitude (Table 6). This was happened as relatively favorable biotic and abiotic condition of the upper altitude for natural growth and reproduction of a variety of most plant species including the one found in the other altitudinal gradients (Melese and Wendawek, 2016). The lowest density for sapling seedling and matured plants were detected in lower gradient might arise due to illegal grazing and tree cutting in lower altitude. The main reason for this difference could be low seed production due to heavy logging for charcoal and timber production and physical damage on seedlings and sapling during cuttings (Adane, 2011). In addition, in the upper and middle slope; the number of mature tree, sapling and seedling were recorded significantly higher than middle lower slope gradients. This is because, the quadrats in this community were might not be suitable for grazing and browsing by animals of the community due its topography. Besides, Soromessa et al. (2004) ensured effect of slope on runoff, drainage, and moisture. The influence of slope and altitude of vegetation on moisture availability affects regeneration status of the vegetation. The anthropogenic disturbances in the study area are higher at the lower altitude (for example, logging and wood collection) and lower at the higher altitude. This is could be the main reason for the increasing trend of regeneration status along altitudinal gradient. The regeneration of the forest is affected not only by environmental factors but also by anthropogenic activities (cutting of trees for charcoal production, constructing wood and fence (Deribe, 2006).
Population structure of plants across edaphic and topographic variables
Population structure is one of the forest characteristics that can be easily influenced by disturbance. Different disturbance factors also affect population structure differently (Adane, 2011). Plant height and diameter at breast height was significant differences (p<0.001) along human disturbance and grazing intensity. It was observed that the higher plant height and diameter at breast height was recorded in no obvious disturbance with mean of 5.74± 0.14 cm and 9.94 ± 0.32 cm, respectively. On the other hand, the lower plant height and diameter at breast height was recorded in the heavy disturbance with the mean of 4.07 ± 0.12 and 6.18±0.21 cm and the higher plant height and diameter at breast height was computed in nil and slight grazing intensity with significant differences (p<0.001) along grazing intensity (Table 7). The reason greater plant height recorded in the no obvious disturbance and nil grazing could be due to the presence of higher species density, less human and livestock interference than in the others. The possible reason for decreasing the mean value of diameter at breast height in the heavy disturbance and heavy grazing site might be due to illegal cutting system used by the local people for construction materials and fuel wood consumption. This study is in line with the study made by Getaneh (2007). Tefera et al. (2015), who showed that the local people for construction and charcoal preparation harvested woody species with diameter at breast height > 30cm. It is also clear those cuttings (be it for charcoal or timber) affect basal area as it targets bigger diameter trees (Adane, 2011). Disturbance regimes may play a considerable role in structuring the pattern of diversity distribution (Jentsch et al., 2002).
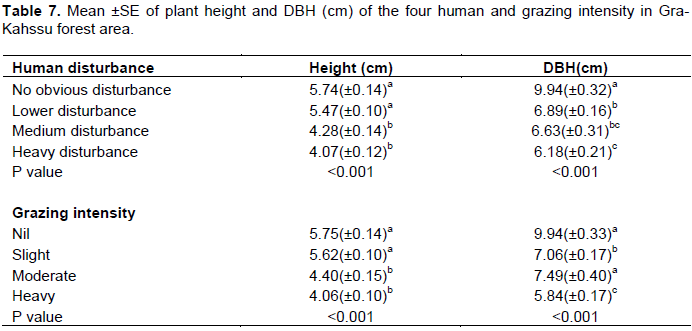
Due to the altitudinal gradient, the values of plant height and diameter at breast height varied (Table 8). The upper altitudinal class had the highest plant height of 5.68m, whereas the lower altitudinal class had the lowest plant height with the recorded value of 4.31m. In addition, the diameter at breast height was significantly lower in the lower altitudinal class compared to the other altitudinal class (p<0.001) (Table 8). The diameter at breast height was significantly larger in the upper slope class compared to the others slope class (p<0.001).The average value of diameter at breast height at the lower slope class was 6.34cm. Similarly, the mean plant height was varied in classes of lower, middle and higher slope with mean value of 4.49±0.08, 5.13±0.12 and 5.76±0.12cm, respectively (Table 8). Generally, the present study shown distinct pattern of variation of plant height and diameter at breast height in altitudinal and slope class although the variation has significant difference.
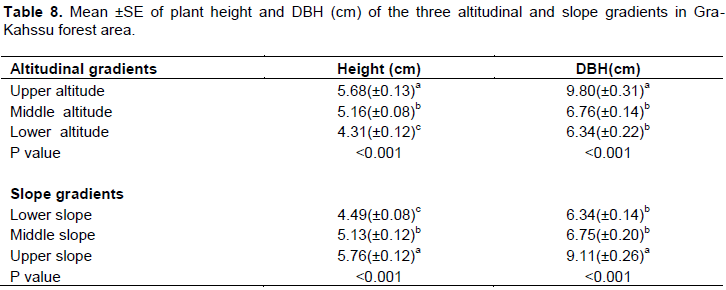
Relationship between vegetation patterns and environmental factors
Vegetation patterns are determined by environmental factors that exhibit heterogeneity over space and time, such as topography as well as human disturbances (Alexander and Millington, 2000). In the present study, the variation of plant diversity was closely related to environmental factors, including altitude, disturbance intensity and slope among which altitude and disturbance were the most important. Species richness and diversity all showed significant relationships with altitude and disturbance intensity (Jin et al., 2013). Species richness was negatively correlated with grazing intensity (r = −0.59, p < 0.001), and number of individuals/ha was positively correlated with slope (r = 0.46, p < 0.001). Disturbance intensity was significantly correlated with number of individuals/ha, species richness and Shannon diversity index (r = −0.67,-0.4 and -0.57) respectively (Table 9). Species richness, Shannon diversity index and evenness were significantly correlated with altitude and disturbance. Altitude is a key variable affecting species diversity in mountains (Zhang et al., 2006; Muhumuza and Byarugaba, 2009; Chawla et al., 2008). The effects of slope on vegetation were also significant, confirming the results of many other studies (Lovett et al. 2006; Zhang and Zhang, 2007 and Jin-Tun et al., 2016). Changes in slope may lead to changes in humidity and temperature, all of which affect species diversity (Virtanen et al., 2010). Basal area was negatively correlated with grazing intensity and human disturbance (r = −0.3, p =0.01), and positively correlated with altitude (r = 0.44, p < 0.001) (Table 9). Altitude and slope have an equally strong significant (p< 0.0001) effect on basal area of species (Markos and Simon, 2015). Although the linear trend explains a significant amount of the variability in basal area on the altitude gradient (Carpenter, 2005).
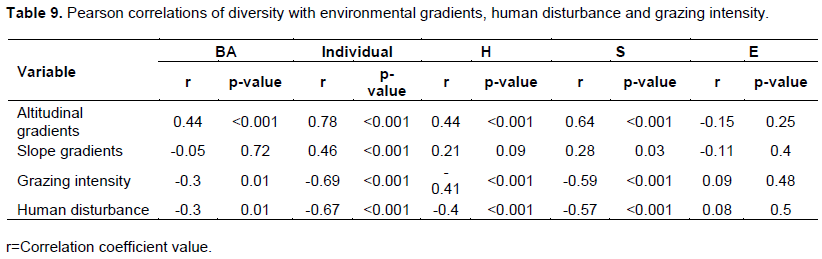
Altitude have strong correlation with species richness and Shannon diversity index of plant and have great impact especially on woody vegetation distribution (Zewde, 2014). Forest diversity was most signiï¬cantly correlated with altitude and disturbance (Jin-Tun et al., 2016). Species richness, Shannon diversity index and evenness all signiï¬cantly correlated with elevation and disturbance gradients. The study of Wana and Carl (2010) in the southwest Ethiopian highlands found that the correlation coefï¬cients for woody species richness and altitude were relatively lower (0.41). Forest composition, diversity and distribution pattern generally are signiï¬cantly correlated with environmental gradients that exhibit heterogeneity over space and time, such as topography and disturbances (Korner, 2007; Brinkmann et al., 2009; Zhang et al., 2015). This was proven true in the Gra-Kahsu forest. The variation of forests was closely correlated with environmental variables such as altitude, grazing, disturbance and slope.
CONCLUSION
The decline in species richness and Shannon and Weaner diversity index of woody vegetation was an outcome of the edaphic and topographic variables. Therefore, this study indicated that different edaphic and topographic variables has great role for the variation of plant species diversity in the forest. Altitudinal gradients, human disturbance and grazing intensity played key roles in plant species diversity. This is because, with altitudinal gradients, human disturbance and grazing intensity play major roles and composition tends varies; and this affects the diversity. A reduction in woody plant species diversity parameters was observed as a direct effect of human disturbance, grazing pressure and altitudes in the Gra-Kahsu forest and this consequently affected the growth, diversity, density, regeneration and population structure of woody plant species. Measures such as controlling grazing intensity and human disturbance, monitoring forest diversity change, and effective management should be taken serious in this forest.
CONFLICT OF INTERESTS
The author has not declared any conflict of interests.
REFERENCES
|
Acharya BK, Chettri B, Vijayan L (2011). Distribution pattern of trees along an elevation gradient of Eastern Himalaya, India. Oecology 37:329-336. |
|
|
Adane G (2011). Plant Communities, Species Diversity, Seedling Bank and Resprouting in Nandi Forests, Kenya. Doctoral dissertation, Universitätsbibliothek Koblenz. |
|
|
Alemayehu M (2003). Country Pasture/Forage Resource Profiles. Addis Ababa, Ethiopia: Addis Ababa University, Faculty of Science, Biology Department. |
|
|
Alemayehu WE (2007). Ethiopian Church Forests: opportunities and challenges for restoration, PhD thesis, Wageningen Univeristy, Wageningen, The Netherlands. |
|
|
Alexander R, Millington A (2000). Vegetation Mapping: From Patch to Planet. Chichester, United Kingdom: Wiley. |
|
|
Althof AJ (2005). Human Impact on Flora and Vegetation of Kakamega Forest, Kenya Structure, distribution and disturbance of plant communities in an East African rainforest. PhD Dissertation University of Koblenz-Landau. |
|
|
Amsalu S (2000). Herbaceous species composition, dry matter and condition of the major grazing areas in the middle rift valley, Ethiopia. MSc Thesis. Hramaya University, Dire Dewa, Ethiopia 159 p. |
|
|
Angassa A (2014). Effects of grazing intensity and bush encroachment on herbaceous specie and rangeland condition in southern Ethiopia. Land degradation and Development 25:438-451. |
|
|
Austrheim G (2002). Plant diversity patterns in semi-natural grasslands along an elevational gradient in southern Norway. Plant Ecology 161(2):193-205. |
|
|
Aynekulu E, Manfred D, Tsegaye D (2009). Regeneration response of Juniperus procera and Olea europaea subsp cuspidata to exclosure in a dry Afromontane Forest in Northern Ethiopia. Mountain Research and Development 29:143-152. |
|
|
Bone L, Lawrence M, Magombo Z (1997). The effect of a Eucalyptus camaldulensis (Dehn) plantation on native woodland recovery on Ulumba Mountains, Southern Malawi. Forest Ecology and Management 99:83-99. |
|
|
Brinkmann K, Patzelt A, Dickhoefer U, Schlecht E, Buerkert A (2009). Vegetation patterns and diversity along an altitudinal and a grazing gradient in the Jabal al Akhdar Mountain range of northern Oman. Arid Environment 73:1035-1045. |
|
|
Brown JH (2001). Mammals on mountainsides: Elevational patterns of diversity. Global Ecology and Biogeography 10:101-109. |
|
|
Bruun HH, Moen J, Virtanen R, Grytnes JA, Oksanen L, Angerbjorn A (2006). Effects of altitude and topography on species richness of vascular plants, bryophytes and lichens in alpine communities. Journal of Vegetation Science 17:37-46. |
|
|
Carpenter C (2005). The environmental control of plant species density on a Himalayan elevation gradient. Journal of Biogeography 32(6):999-1018. |
|
|
Chang-Ming Zhao, Wei-Lie Chen, Zi-Qiang Tian, Zong-Qiang XIE (2005). Altitudinal Pattern of Plant Species Diversity in Shennongjia Mountains, Central China. Journal of Integrative Plant Biology 47(12):1431-1449. |
|
|
Chawla A, Rajkumar S, Singh K, Lal B, Singh R, Thukral A (2008). Plant species diversity along an altitudinal gradient of Bhabha Valley in western Himalaya. Journal of Mountain Science 5:157-177. |
|
|
De Lafontaine G, Houle G (2007). Species richness along a production gradient: a multivariate approach. American Journal of botany 94(1):79-88. |
|
|
Deribe G (2006). Humbo community-managed natural regeneration project final report. Report, Humbo Regeneration Project, Addis Ababa, Ethiopia. |
|
|
Desalegn W, Beierkuhnlein C (2010). Plant species and growth form richness along altitudinal gradients in the southwest Ethiopian highlands. Journal of vegetation science 21(4):617-626. |
|
|
Fetene M, Assefa Y, Gashaw M, Woldu Z, Beck E (2006). Diversity of afroalpine vegetation and ecology of tree line species in the Bale Mountains, Ethiopia, and the influence of fire. In: Spehn EM, Liberman M and Körner C, editors. Land Use Change and Mountain Biodiversity. New York, NY: CRC, pp. 25-38. |
|
|
Feyera S, Christine S, Tadese W, Hans J, Manfred D (2014). Plant diversity, vegetation structure and relationship between Plant communities and environmental variables in The Afromontane forests of Ethiopia, SINET: Ethiopian Journal of Science 37(2):113-130. |
|
|
Ganesan R, Siddappa R (2004). Regeneration of Amla, an important non-timber forest product from southern India. Conservation and Society 2:365-375. |
|
|
Getaneh B (2007). Floristic composition and structure in Beschillo and Abay (Blue Nile) riverine vegetation. Doctoral Dissertation. Addis Ababa University, Addis Ababa, Ethiopia. |
|
|
Guarino ESG, Scariot AO (2012). Tree seedling survival and growth in logged and undisturbed seasonal deciduous forest fragments in central Brazil. Journal of Forest Research 17:193-201. |
|
|
Gunnar A, Ove E (2001). Plant Species Diversity and Grazing in the Scandinavian Mountains: Patterns and Processes at Different Spatial Scales, Ecography 24(6):683-695. |
|
|
Hadera G (2000). A study on the ecology and management of the Dessa forest in the northeastern escarpment of Ethiopia. M.Sc. Thesis A.A.U. |
|
|
Herlocker DJ (1999). Rangeland ecology and resource development in eastern Africa. GTZ, German Technical Cooperation. |
|
|
Hoshino A, Yoshihara Y, Sasaki T, Okayasu T, Jamsran U, Okuro T, Takeuchi K (2009). Comparison of vegetation changes along grazing gradients with different numbers of livestock. Journal of Arid Environments 73:687-690. |
|
|
Huston MA (1994). Biological diversity: the coexistence of species. Cambridge University Press. |
|
|
Jentsch A, Beierkuhnlein C, White PS (2002). Scale, the dynamic stability of forest ecosystems, and the persistence of biodiversity. Silva Fennica 36:393-400. |
|
|
Jin TZ, Bin X, Min L (2013). Vegetation patterns and species diversity along elevational and disturbance gradients in the Baihua Mountain Reserve, Beijing, China. Mountain Research and Development, 33(2):170-178. |
|
|
Jin-Tun Z, Mi Z, Rui M (2016). Effects of altitude and disturbance gradients on forest diversity in the Wulingshan Nature Reserve, North China. Environmental Earth Sciences 75:904. |
|
|
Kebrom T, Backeus I, Skuglund J, Zerihun W (1997). Vegetation on hillslopes of Wello, Ethiopia: Degradation and regeneration. Nordic Journal of Botany 17(5):483-493. |
|
|
Körner C (2007). The use of altitude in ecological research. Trends in Ecology and Evolution 22:569-574. |
|
|
Kumelachew Y, Tamrat B (2002). Plant community analysis and ecology of afromontane and transitional rainforest vegetation of Southwestern Ethiopia, SINET: Ethiopian Journal of Science 25(2):155-175. |
|
|
Leul K, Sileshi N, Zerihun W (2015). The effects of disturbance on the population structure and regeneration potential of five dominant wood species in Hugumburda Gratkhassu National Forest Priority Area, Northeastern Ethiopia. African Journal of Ecology 54(1):20-28. |
|
|
Leul K, Tamrat B, Sileshi N (2010). Vegetation Composition in Hugumbirda-Gratkhassu National Forest Priority Area, South Tigray. Momona Ethiopian Journal of Science 2(2):27-48. |
|
|
Loreau M, Naeem S, Inchausti P (2001). Biodiversity and ecosystem functioning: Current knowledge and future challenges. Science 294:804-808. |
|
|
Lovett JC, Marshall AR, Carr J (2006). Changes in tropical forest vegetation along an altitudinal gradient in the Udzungwa Mountains National Park, Tanzania. African Journal of Ecology 44:478-490. |
|
|
Maestre FT (2004). On the importance of patch attributes, environmental factors and past human impacts as determinants of perennial plant species richness and diversity in Mediterranean Semiarid Steppes. Diversity and Distributions 10:21-29. |
|
|
Magurran AE (2004). Measuring Biological Diversity. Blackwell publishing. Australia. |
|
|
Markos K, Simon S (2015). Floristic composition, vegetation structure, and regeneration status of woody plant species of Oda forest of Humbo Carbon Project, Wolaita, Ethiopia. Journal of Botany. ID 963816, 9p. |
|
|
Melese B, Wendawek A (2016). Floristic Composition and Vegetation Structure of Woody Species in Lammo Natural Forest in Tembaro Woreda, Kambata-Tambaro Zone, Southern Ethiopia. American Journal of Agriculture and Forestry 4(2):49-55. |
|
|
Mueller-Dombois D, Ellenberg H (1974). Aims and Methods of Vegetation Ecology. John Wiley and Sons, London. |
|
|
Muhumuza M, Byarugaba D (2009). Impact of land use on the ecology of uncultivated plant species in the Rwenzori mountain range, mid-western Uganda. African Journal of Ecology 47:614-621. |
|
|
Oba G, Veetas OR, Stenseth NC (2001). Relationships between biomass and plant species richness in arid zone grazing lands. Journal of Applied Ecology 38:836-845. |
|
|
Ole R Vetaas, John-Arvid G (2002). Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal. Global Ecology and Biogeography 11:291-30. |
|
|
Otypkova Z, Chytry M, Tichy L, Pechanec V, Jongepier JW, Hajek O (2011). Floristic diversity patterns in the White Carpathians biosphere reserve, Czech Republic. Biologia, 66:266-274. |
|
|
Pavlu V, Hejcman M, Pavlu L, Gaisler J (2003). Effect of rotational and continuous grazing on vegetation of an upland grassland in the Jizerske Hory Mts., Czech Republic. Folia Geobot 38(1):21-34. |
|
|
R Development Core Team (2015). R: a language and environment for statistical computing. R Foundation for Statistical Computing |
|
|
Rahbek C. 2005. The role of spatial scale and the perception of large-scale species richness patterns. Ecology Letters 8:224-239. |
|
|
Rembold K (2011). Conservation status of the vascular plants in East African rain forests. PhD Dissertation, University of Koblenz-Landau. |
|
|
Ru W, Zhang JT (2012). Ecological study of forests dominated by endangered species, Taxus chinensis var. mairei, in Shanxi of China. Applied Ecology and Environmental Research 10(4):457-470. |
|
|
Shannon CE, Weaner W (1949). The Mathematical Theory of Communication: Unknown Distance Function. Urbana, IL: Illinois Press. |
|
|
Sharma CM, Suyal S, Gairola S, Ghildiyal SK (2009). Species richness and diversity along an altitudinal gradient in moist temperate forest of Garhwal Himalaya. Journal American Science 5(5):119-128. |
|
|
Smet M, Ward D (2005). A comparison of the effects of different rangeland management systems on plant species composition, diversity and vegetation structure in a semi-arid savanna. African Journal of Range and Forage Science 22(1):59-71. |
|
|
Soromessa T, Teketay D, Demissew S (2004). Ecological study of the vegetation in Gamo Gofa zone, southern Ethiopia. Tropical Ecology 45(2):209-222. |
|
|
Sproull GJ, Quigley MF, Sher A, Gonzalez E (2015). Long-term changes in composition, diversity and distribution patterns in four herbaceous plant communities along an elevational gradient. Journal of Vegetation Science 26:552-563 |
|
|
Tefera J, Minyishaw E, Kebenu F, Nurhusen A, Gufran M (2015). Woody species diversity, management and contribution of local community in Protected Forest of Kuneba District, Afar Region Ethiopia. Journal of Environmental Science and Technology 8(4):172-179. |
|
|
Tefera M, Teketay D, Hulten H, Yemshaw Y (2005). The role of enclosure in the recovery of woody vegetation in degraded dry land hillsides of central and northern Ethiopia. Journal of Arid Environments 60(2):259-281. |
|
|
Teshome S. Demel T, Sebsebe D (2004). Ecological study of the vegetation in Gamo Gofa zone, Southern Ethiopia, Journal of Tropical Ecology 45(2):209-221. |
|
|
Tessema ZK, De Boer WF, Baars RMT, Prins HHT (2011). Changes in vegetation structure, herbaceous biomass and soil nutrients in response to grazing in semiâ€arid savannas in Ethiopia. Journal of Arid Environment 75:662-670. |
|
|
Tibebu Y, Teshome S (2015). Carbon Stock Variations along Altitudinal and Slope Gradient in the Forest Belt of Simen Mountains National Park, Ethiopia. American Journal of Environmental Protection 4(4):199-201. |
|
|
Vetaas OR, Grytnes JA (2002). Distribution of vascular plant species richness and endemic richness along the Himalayan elevation gradient in Nepal. Global Ecology and Biogeography 11:291-301. |
|
|
Virtanen R, Luoto M, Rama T, Mikkola K, Hjort J, Grytnes JA, Birks HJB (2010). Recent vegetation changes at the high-latitude tree line ecotone are controlled by geomorphological disturbance, productivity and diversity. Global Ecology and Biogeography 19:810-821. |
|
|
Wana De, Carl B (2010). Plant species and growth form richness along altitudinal gradients in the southwest Ethiopian highlands, Journal of Vegetation Science 21:617-626. |
|
|
Wereda Alamata Office of Agricultural and Rural Development (WAOARD) (2017). Natural Resources Core Process, annual report. Alamata Wereda Office of Agriculture and rural Development, Unpublished, Tigrigna version. |
|
|
Wolde M, Veldkamp E, Corre MD (2011b). Restoration of ecosystem carbon stocks following enclosure establishment in communal grazing lands in Tigray, Ethiopia. Soil Science Society of America Journal 75(1):246-256. |
|
|
Wolde M, Veldkamp E, Mesfin T, Olschewski R (2011c). Economic valuation of land restoration: The case of exclosures established on communal grazing lands in Tigray, Ethiopia. Land degradation and development 22:334-344. |
|
|
Wolde M, Aynekulu E (2011a). Enclosure land management for restoration of the soils in degraded communal grazing lands in northern Ethiopia. Land Degradation and Development 24:528-538. |
|
|
Xiang CL, Zhang JT (2009). Changes in species diversity and contributing factors in subalpine meadows in Dongling Mountain. Journal of Beijing Normal University (Natural Science) 45(3):275-278. |
|
|
Yohannes H, Teshome S, Mekuria A (2015). Carbon stock analysis along altitudinal gradient in gedo forest: Implications for forest management and climate change mitigation. American journal of environmental protection 4(5):237-244. |
|
|
Zewde A (2014). Distribution of the Woody Vegetation along the Altitudinal range from Abay (Blue Nile) Gorge to Choke Mountain, East Gojjam Zone, Amhara National Regional State, Northwest Ethiopia. MSc. Thesis, Department of Plant Biology and Biodiversity Management, Addis Ababa University, Ethiopia. |
|
|
Zhang JT, Zhang F (2007). Diversity and composition of plant functional groups in mountain forests of the Lishan Nature Reserve, North China. Botanical Studies 48:339-348. |
|
|
Zhang JT, Ru WM, Li B (2006). Relationships between vegetation and climate on the Loess Plateau in China. Folia Geobotanica, 41:151-163. |
|
|
Zhang JT, Xu B, Li M (2013). Vegetation patterns and species diversity along elevational and disturbance gradients in the Baihua Mountain Reserve, Beijing, China. Mountain Research and Development 33(2):170-178. |
|
|
Zhang JT, Zhang B, Qian ZY (2015). Functional diversity of Cercidiphyllum japonicum, communities in the Shennongjia Reserve, central China. Journal of Forest Research 26(1):171-177. |
|
|
Zhao CM, Chen WL, Tian ZQ, Xie ZQ (2005). Altitudinal pattern of plant species diversity in Shennongjia Mountains, Central China. Journal of Integrative Plant Biology 47:143-449. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


