Full Length Research Paper

ABSTRACT
Tamarind is native to tropical parts of Africa and Asia. It shows considerable phenotypic variation in morphological and horticultural traits that can be utilized in its genetic improvement. In Kenya, there exists a wide range of tamarind germplasm that has not been characterized. Initial characterization is based on morphological descriptors. The objective of this study is to evaluate morphological diversity of tamarind germplasm from Eastern parts of Kenya. Tamarind germplasms were collected from Kitui, Mwingi, Masinga, Embu and Kibwezi and then characterized using morphological descriptors based on seed, fruit and stem. Morphological characters were recorded and data from eighty-nine accessions were submitted to principal component and hierarchic ascendant analysis (HAC) and Euclidian average distance. Accessions from Kibwezi, Embu and Kitui showed the greatest diversity while accessions from Masinga and Mwingi had the least diversity. Trunk diameter at ground, pod weight, number of seeds/pod, height to the first branch and pod width showed greatest variation in principal component analysis. High morphological diversity obtained in these regions can be used to initiate new breeding and conservation programmes in tamarind for improved fruit and tree crop.
Key words: Tamarind, morphology, diversity, accessions, principle component.
INTRODUCTION
Tamarind (Tamarindus indica L) belongs to the family Leguminoseae (Khanzada et al., 2008). It is an evergreen tree that is native to tropical and subtropical regions of Africa and South Asia (Doughari, 2006). It is primarily used for it fruits that are either processed into juices, eaten directly, used in preparation of recipe and preservatives (Gullipalli and Kasiviswanatham, 2013). The leaves, bark, and pulp have extensively been used in ethnobotany (Gupta et al., 2014). The tree is widely used as an ornamental tree due to its availably and wide use (Doughari, 2006). It has significant importance in the cosmetics, paints and varnishes industries (Santos et al., 2012).
Morphological descriptors have been used as basic character in identification of plants, in breeding, commercialization, conservation of plant resources, cluster analysis understanding genetic similarities and dissimilarities (Santos et al., 2012; Cervantes and Diego, 2010). Components of fruits such as fruit size, shape, color and general appearance are important in plant description (Nasution and Chinawat, 2017). Morphological descriptors have limitations in distinguishing sub families and tribes as the traits are similar (Swenson and Anderberg, 2005).
T. indica trees are morphologically different in terms of fruit, crown, foliage, trunk, seed and flower characteristics (Nadine et al., 2011). In Thailand Nasution and Chinawat (2017) clustered sweet tamarind based on fruit characters. In a study Algabal et al. (2010) reported differences in pulp color used to distinguish the cultivars. Gunasena et al. (2007)reported that reproductive and fruit traits differ among different population of tamarind and they are influenced either by the environment or genetic make-up.
Morphological traits have been used to study Asian tamarind populations and the results revealed that there existed both morphological and genetic differences (Fandohan et al., 2010). These descriptors enabled them to choose superior cultivars for the market in terms of taste, pulp and thickness (El-Siddig et al., 2006). In West Africa most studies focused on biochemical compounds of tamarind (Adeola and Aworh, 2012). In Eastern parts of Kenya no studies have been carried out to compare morphological differences among tamarind populations in different places. The objective of this study is to evaluate morphological differences among tamarind accessions from the Eastern parts of Kenya. The results from morphological evaluation and clustering of tamarind will be useful in cultivar selection and improvement of breeding programmes for tamarind
MATERIALS AND METHODS
Collection of tamarind germplasm
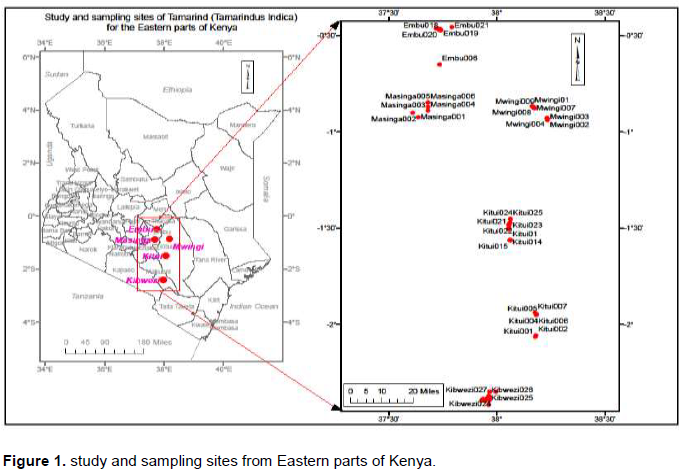
Sampling was done in December 2015 to August 2016. Study sites included: Kitui, Mwingi, Masinga, Embu and Kibwezi (Figure 1). Tamarind farms were identified using key informants and random sampling was done randomly in the farms.
Morphological characterization
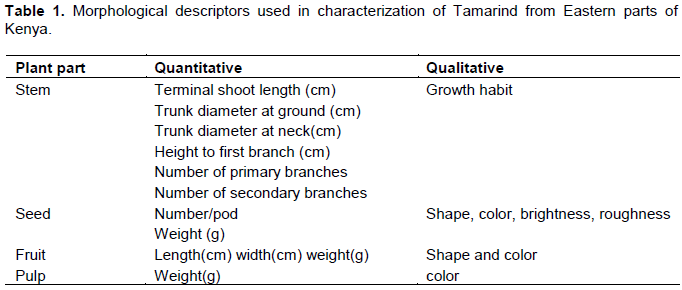
Characterization was done according to International Union of Plant Protection of New Vegetal Variants (UPOV 1987), International Committee of Genetic Resources of Plant for the description of tropical plants (IPGRI, 1991; Fandohan et al., 2010). Twenty descriptors were used for characterization (Table 1).
Data collection
Data collected from tamarind germplasm surveys included trunk diameter at ground, trunk diameter at the neck, height to the first branch. Pod length was determined as an average of five pods from pole to pole. Pod width was determined as mean of five pods from the equator of the cross section of the fruit and pod weight was determined as a mean of five pods of the same tree. Seed weight was determined as an average of seeds in an entire pod. Pulp weight was determined as the average of pulp in 5 pods. Seed number was determined as an average of seeds in 5 pods. Growth habit was either orthotropic or plagiotropic
Data analysis
Quantitative data were submitted to principal component analysis (PCA), using the R software statistical package. Cluster analysis was carried out on the principal components with SS loadings 0f 0.98 to 2.93 using the hierarchic ascendant analysis (HAC).
RESULTS
Morphological diversity of tamarind accessions from Eastern parts of Kenya
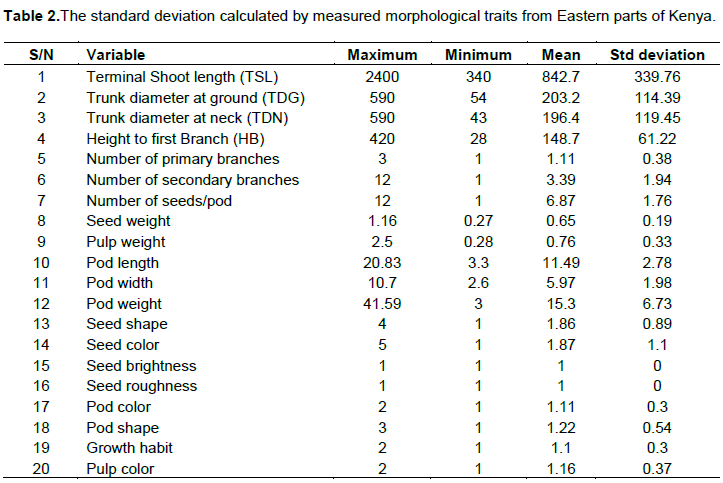
Tamarind from Eastern parts of Kenya showed a wide variation in tree characteristics including seed color, seed shape, seed number per pod, seed weight, pod shape, pod color, pod length, pod width, pod weight, pulp color, pulp weight, tree habit, terminal shoot length, trunk diameter at ground, trunk diameter at neck, height to the first branch, number of primary branches, number of secondary braches (Plate 1 and Table 2).

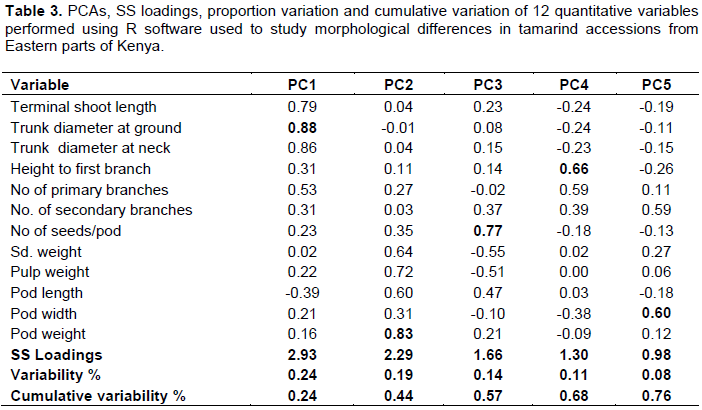
Principal component analysis
The first five components of principal components in quantitative analysis explained 76% of total variations (Table 3). Eleven traits contributed to PC1 with trunk diameter at the ground contributing more positively. In PC2 eleven traits contributed positively to the component with pod weight having a significant positive contribution to the PC. In PC3 eight traits contributed positively to the component with number of seeds per pod having a significant positive contribution. PC4 had six traits that contributed positively with height to the first branch having the highest positive contribution. In PC5, six traits had positive contribution with pod width having the greatest contribution.
Correlation among characters showed three clusters. In the first cluster, trunk diameter at the neck (TDN), trunk diameter at ground (TDG), number of secondary branches and terminal shoot length are highly correlated. In the second cluster, number of seeds per pod, pod length, pod weight (PPWT), seed weight and pulp weight were highly correlated. In the third cluster height to the first branch (HB) and number of primary branches were highly correlated (Figure 2).
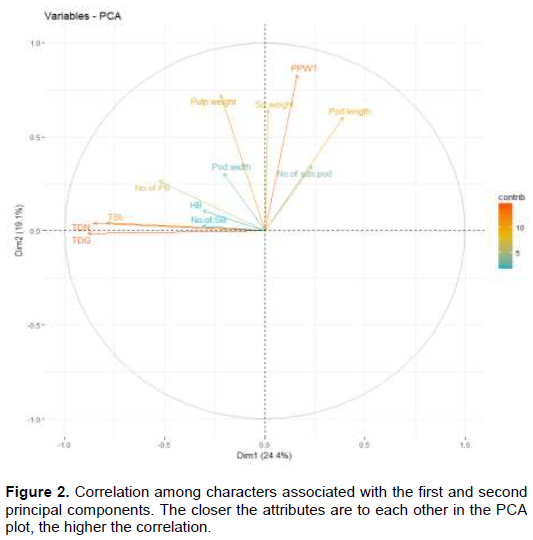
Cluster analysis
HAC distinguished two major clusters when truncated at 1000.Cluster 1 consisted of 50 accessions while cluster 2 had 39 accessions. Each cluster had two sub-clusters. Most diversity was observed from accessions in Kibwezi, Embu and Kitui while least diversity was observed in Mwingi and Masinga. The accessions were distributed across the clusters (Figure 3)
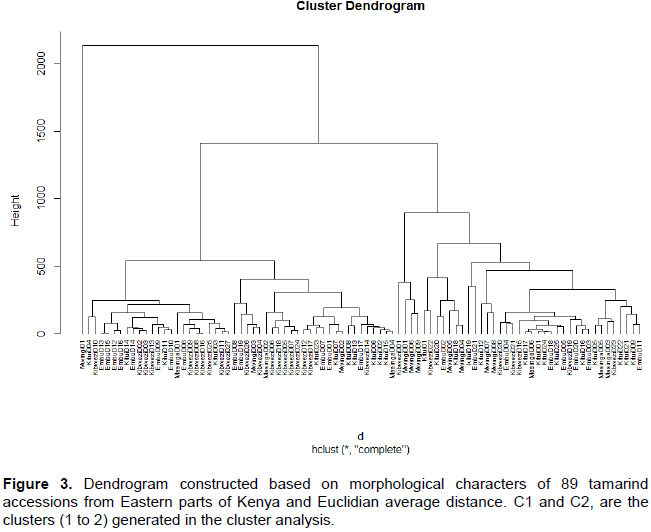
.
DISCUSSION
Morphological descriptors have been used in initial identification of organism (Piyasundura et al., 2008). In this study high morphological diversity was found among accessions collected from Eastern parts of Kenya. This variation is similar to reports by Nyadoi et al. (2014)who reported there was a great diversity among the tamarind populations collected in Maasai region in Kenya. Fandohan et al. (2011)reported 3-8 number of primary branches and 30-60 number of secondary branches but this study revealed that the number of primary braches ranged from 1-2 and secondary branches from 2-12. In this study trunk diameter varied greatly with the trees from intercrop farm having shorter trees than those that grew widely. This is in agreement with reports by Nyadoi et al. (2014)who reported that the diameter greatly varied with the type of vegetation in the habitat. This is different as the study sites were farm lands, savanna and forests and this was more on forests and farm land. Growth habit of orthotropic and plagiotropic was similar to findings by Ali et al. (2010)He studied tamarind from Southern India indicating that the growth habit is not influenced by changes in environment and cultural practices.
Three pod shapes observed (curved, semi curved and straight) were similar to the findings by Algabal et al. (2011) but Fandohan et al. (2010) had only two pod shapes (curved and the straight). The shapes are affected by the seed number and seed shapes which are influenced by its genetics. Pod color was either velvety brown or rusty brown that coincides with the findings of Ayala-Silva et al. (2016). Variations in pod color are highly influenced by the age of the pod and environmental changes. Pulp color varied from light brown to dark brown which slightly varies from the findings by Ayala- Silva et al. (2016) who reported colors of reddish brown and brown. The pulp color is highly influenced by genetic make-up of the plant. Highest diversity was observed in seed color; Fandohan et al. (2011) only recorded three seed colors of black, brown and dark brown but this study revealed other colors of dark brown at the center and brown outside, light brown. the colors reported in this study were also evident in the reports by Fawzi (2011.)This trait is inherited and affected by the environment, and in different environment different colors were observed. Fandohan et al. (2011) also recorded seed shape of quadrant, bowl shape and irregular while Fawzi (2011)reported more of oblong, asymmetrical, ovate and rhomboid but from the studies seed shapes of ovate and D shape were observed. The shape is inherited and also affected by the environment
In this study, the pod weight was 3-31.4 g, while Prerak et al. (2013)reported pod weight of 5.49- 24.55 g. This is directly correlated with pulp weight and seed number. Pulp weight ranged from 0.28-1.92 g while Van den et al. (2014)reported pulp weight of 1.96-4.65 g. Pulp weight is a factor of management practices given to the tree. Van den et al. (2014)also reported that the number of seeds per pod ranged from 5-7 while this study depicted seed range of 1-12 per pod. This is highly influenced by nutrition available for the plant and the management practices that also influence directly the length of the pod
Diversity was not observed in fibre color, seed roughness, seed brilliance and pulp taste. Fibre color observed was yellow brown, all seeds were rough, were non brilliant and pulp was sour. These factors were not altered by different environments. Fandohan et al. (2011) reported both brilliant and non-brilliant seed and rough and polished seeds and this could be affected by different environmental factors. HAC clustering grouped the accessions into two major clusters and two sub clusters. The samples were from across the regions indicating that the diversity was not based on the origin. This is also confirmed by reports of Iddi Garba et al. (2015).
According to Chatfield and Collins (1980) components with eigenvalues less than one should be eliminated; so those with eigenvalues of one and above are used for they are considered to be more significant. The eigenvalues decreased in this from PC1 to PC5 showing a decrease in variation. The first five components of principal components in quantitative analysis explained 76% of total variations among the accessions (Table 3). PCA identified eleven traits namely; trunk diameter at the ground, trunk diameter at the neck, height to first branch, number of primary branches, number of secondary branches, pulp weight, pod weight, seed weight and terminal shoot length that contributed positively to PC1. However, trunk diameter at the ground contributed more positively than the rest of the traits.
PC2 identified eleven traits that contributed positively to the component with pod weight having a significant positive contribution to the PC. PC3 identified eight traits that contributed positively to the PC; number of seeds per pod had significant positive contribution to PC3. PC4 had eight traits that contributed positively, with height to the first branch having the highest contribution. In PC5, eight traits had positive contribution with pod width having the greatest contribution. This research reveals morphological diversity and factors of pod weight, pod width and pulp weight that significant and directly correlated to the fruit should be considered for conservation and future improvement
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENT
The study is funded by Deutscher Akademischer Austausch Dienst (DAAD) in a country programme from 2015-2018.
REFERENCES
|
Adeola AA, Aworh OC (2012). A comparative evaluation of the chemical properties of wild tamarind (Tamarindus indica L.) fruits in Nigeria. Food 6 (1):49-57. |
|
|
Algabal AQAY, Narayanaswamy P, Luke S (2011). "Amplified Fragment Length Polymorphism Marker-Based Genetic Diversity in Tamarind (Tamarindus Indica)." International Journal of Fruit Science 11(1):1-16. |
|
|
Ali QAYA, Narayanaswamy P, Luke S (2010). "Estimation of Genetic Variability in Tamarind (Tamarindus Indica) Using RAPD Markers" 5(1):10-16. |
|
|
Cervantes E, Diego JG (2010). "Morphological Description of Plants: New Perspectives in Development and Evolution 4(1):67-71. |
|
|
Chatfield C, Collins AJ (1980). Introduction to Multivariate Analysis. Springer, Boston, MA. |
|
|
Doughari JH (2006). "Antimicrobial Activity of Tamarindus Indica Linn" Tropical Journal of Pharmaceutical Research 5(2):597-603. |
|
|
El-Siddig K, Gunasena HPM, Prasad BA, Pushpakumara DKNP, Ramana KVR, Vijayanand P, Williams JT (2006). "Southampton Centre for Underutilised Crops, Southampton, UK | Feedipedia." View |
|
|
Fandohan AB, Achille EA, Romain LGK, Brice S, Patrick VD (2010). "Impact of Habitat Type on the Conservation Status of Tamarind (<span Class="italic">Tamarindus Indica</span> L.) Populations in the W National Park of Benin." Fruits 65(1):11-19. |
|
|
Fandohan B, Achille EA, Romain GK, Tina K, Brice S (2011). "Quantitative Morphological Descriptors Confirm Traditionally Classified Morphotypes of Tamarindus Indica L. Fruits." Genetic Resources and Crop Evolution 58(2):299-309. |
|
|
Gullipalli SR, Kasiviswanatham V (2013). "Extraction of Tartaric Acid from Tamarind Pulp and Analysis of the Acid Composition in Leaves" International Journal of Students' Research in Technology and Management 1(5):478-88. |
|
|
Gunasena HPM, Prasad BA, Pushpakumara DKNG, Ramana KVR, Vijayanand P (2007). "Tamarind (Tamarindus Indica L.) | World Agroforestry Centre." 2007. |
|
|
Gupta C, Prakash D, Gupta S (2014). "Studies on the Antimicrobial Activity of Tamarind (Tamarindus Indica) and Its Potential as Food Bio-Preservative" International Food Research Journal 21(6):2437-2441. |
|
|
Iddi Garba NM, Yacoubou B, Mainassara Z, Sanoussi A, Maârouhi IM, Moutari A, Falalou H, Idi SS, Ali M, Mahamane S (2015). "Evaluation of agro-morphological diversity of groundnut (Arachis hypogaea L.) in |
|
|
Khanzada SK, Shaikh W, Sofia S, Kazi,TG, Usmanghani K, Kabir A, Sheerazi TH (2008). Chemical constituents of Tamarindus indica L. medicinal plant in Sindh. Pakistan Journal of Botany 40(6):2553-2559. |
|
|
Nasution F, Chinawat Y (2017). "Clustering of Five Sweet Tamarind Based on Fruit Characteristic" AGRIVITA, Journal of Agricultural Science 39(1):38-44. |
|
|
Niger." African Journal of Agricultural Research 10(5):334-344.2015. "Evaluation of Agro-Morphological Diversity of Groundnut (Arachis Hypogaea L.) in Niger" 10(5):334-44. |
|
|
Nyadoi PP, Okori JBL, Okullo JO, Fluch S, Burg K, Jamnadass R (2014). "Tamarindus indica L. patterns of diversity from the genetic to the niche-species level in East Africa." Tropical Ecology 55(1). |
|
|
Piyasundura JHN, Gunasekara MTK, Wickramasinghe IP (2008). "Identification of Descriminating Morphological Descriptors for Characterization of Tea (Camelia Sinensis L) Germplasm in Sri Lanka" 20:193-199. |
|
|
Prerak B, Meena CB, Singh U, Jain MC, Bhim S, Dashora LK (2013). "Exploring the Unexplored Tamarind in Rajasthan" Environment and Ecology 31(4):1739-42. |
|
|
Santos RC, Pires JL, Correa RX (2012). "Morphological Characterization of Leaf, flower, Fruit and Seed Traits among Brazilian Theobroma L. Species" Genetic Resources and Crop Evolution 59(3):327-345. |
|
|
Swenson U, Anderberg AA (2005). "Phylogeny, Character Evolution, and Classiï¬cation of Sapotaceae (Ericales)" Cladistics 21:101-30. |
|
|
UPOV (1987). International union of Plant protection of new Vegetal Variants. |
|
|
Van den B, Alaerts NK, Ghaffaripour S, David S, Roeland S (2014). Physico-Chemical Properties of Tamarind (Tamarindus Indica L.) Fruits from Mali: Selection of Elite Trees for Domestication. P. 61. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


