ABSTRACT
The synthetic antimicrobial peptide SAAP-148 was found to be effective against a wide range of (antimicrobial-resistant) gram-positive and gram-negative bacteria, in vitro and in a superficial skin infection model in mice, but not a surgical wound infection model in rats. In addition, the efficacy of SAAP-148 in protein-rich environments such as blood plasma and eschar extract is considerably reduced as compared to PBS. The aim of the present study is to determine which wound-related factors reduce SAAP-148’s efficacy. As high dosages of SAAP-148 may be required in infected wounds we also assessed the in vitro and ex vivo cytotoxicity of SAAP-148. Preincubation of SAAP-148 in 50% (v/v) blood plasma or eschar extract for 24 h, 3% (v/v) bovine serum albumin or a collagen-elastin 3D matrix significantly reduced the bactericidal efficacy of the peptide. Also, the presence of protease inhibitors did not restore the bioactivity of SAAP-148, suggesting a reduced bioavailability of SAAP-148 due to protein binding, rather than degradation. Furthermore, relatively low concentrations of SAAP-148 (≥0.23 nmol/100 µl) induced cytotoxicity for human skin cells in 2D culture. Contrarily, SAAP-148 did not induce cytotoxicity for cells in ex vivo human skin. In addition, unlike silver sulfadiazine (SSD), SAAP-148 showed no statistically significant adverse effects on the re-epithelialization of ex vivo excision wounds and burn wounds. Hence, SAAP-148 potentially has a beneficial profile for antimicrobial treatment of infected wounds.
Key words: SAAP-148, proteolytic degradation, bioactivity, peptide interaction, cytotoxicity, skin and soft tissue infection.
Abbreviation:
AMPs, antimicrobial peptides; BrdU, 5-bromo-2- deoxyuridine; BSA, bovine serum albumin; CFU, colony forming units; DMEM, Dulbecco's Modified Eagle Medium; DNA, deoxyribonucleic acid; EDTA, ethylenediaminetetraacetic acid; EWM, excision wound model; FCS, fetal calf serum; LB, Luria Bertani; LDH, lactate dehydrogenase; MRSA, methicillin resistant Staphylococcus aureus; PBS, phosphate-buffered saline; P/S, penicillin/streptomycin; RPMI, Roswell Park Memorial Institute; SD, standard deviation; SPS, sodium polyanethol sulfonate; SSD, silver sulfadiazine.
Bacterial colonization of burn wounds is common because the skin, which normally acts as a physical barrier, is compromised by the injury (Glik et al., 2012). Opportunistic bacteria such as Staphylococcus aureus have the ability to evade the immune responses (Rooijakkers et al., 2005), which can lead to uncontrolled bacterial colonization in burns. To prevent and control bacterial colonization in burn wounds, topical treatments such as silver sulfadiazine (SSD), mupirocin and fusidic acid in ointments have been developed (Atiyeh et al., 2007; Gould and Villiger, 1986; Williamson et al., 2017). However, limited bactericidal efficacy, emergence of drug-resistance or cytotoxicity for the host cells induced by these topicals has increased the need for novel antimicrobial agents.
In the fight against antibiotic-resistant bacteria, antimicrobial peptides (AMPs) have been identified as promising therapeutic agents (Gordon et al., 2005). AMPs are usually small cationic molecules ranging from 12 to 50 amino acids with an amphiphilic tertiary structure. They are effective against a wide range of bacteria as their amphiphilic characteristics allow them to penetrate into the bacterial membrane (Beisswenger and Bals, 2005; Shai, 2002). Due to this anticipated mechanism of action, resistance against AMP is less likely to occur (Zasloff, 2002). The variety in antibacterial efficacy that has been observed among different naturally occurring AMPs is related to the broad diversity in the amino acid sequence and secondary structure (Kumar et al., 2018). These structural properties of AMPs also determine their stability in biological environments and potential toxicity for human cells (Yin et al., 2012).
Previously, a novel synthetic AMP, known as SAAP-148, was identified as a promising agent to combat (resistant) bacteria (de Breij et al., 2018). This peptide effectively eradicated methicillin resistant S. aureus (MRSA) in a skin infection mouse model (de Breij et al., 2018). However, SAAP-148 did not eradicate MRSA in a partial thickness wound model in rats (Dijksteel et al., 2019a). We hypothesize that this effect was mainly caused by the protein and protease-rich wound environment of the open wound. Hence, the aim of the present study is to determine the functional stability and bioactivity of SAAP-148 in conditions mimicking the wound environment. As relatively high SAAP-148 concentrations are needed to eliminate bacteria in biological relevant environments, we also determined the cytotoxicity of such SAAP-148 concentrations for human cells in 2D culture and in ex vivo human skin, and estimated the therapeutic index.
Ethylenediaminetetraacetic acid (EDTA) was purchased from Gibco-BRL (Life Technologies, N.Y. USA) and protease inhibitor mini tablets (EDTA-free) were purchased from Pierce Biotechnology (Rockford, UK). Novomaix™ is a collagen-elastin 3D scaffold, which was a kind gift from Matricel GmbH (Herzogenrath, Germany). SAAP-148 was obtained from the Leiden University Medical Center (Leiden, the Netherlands). For the synthesis, purification and identification of SAAP-148 see Nell et al. (2006). Lyophilized peptide was dissolved in phosphate-buffered saline (PBS; Gibco, Paisley, UK) and stored in aliquots at −20°C until use.
Human blood plasma and tissue
Human blood and tissues were obtained according to institutional guidelines and medical research “code of conduct for responsible use”, drafted by Federa (Foundation Federation of Dutch Medical Scientific Societies). Human blood plasma from three donors was obtained from Sanquin (Leiden, the Netherlands). The plasma was pooled, centrifuged at 2266 ´ g for 10 min, filtered using 0.2 µm filters to remove aggregates and stored in aliquots of 50 ml at -20°C. Eschar or burn wound tissues was obtained from the Burn Center, Red Cross Hospital (Beverwijk, the Netherlands). An extract was prepared from eschar as previously described (Dijksteel et al., 2019a).
Prior to the experiment, blood plasma or eschar extract were thawed and centrifuged at 2738 ´ g for 10 min. Proteins from blood plasma or eschar extract were precipitated using methanol at a final concentration of 90%. The protein suspension was centrifuged at 10000 ´ g for 10 min and the pellet was washed and dissolved in PBS.
Cell cultures
The epidermis of 0.3 mm split-thickness skin grafts was mechanically separated from the dermis after 30 min incubation with 0.25% (wt/v) dispase (Gibco) in PBS at 37°C and 5% CO2 using two forceps. Keratinocytes were isolated from the epidermis after 15-25 min incubation with 0.05% (wt/v) trypsin in 0.53 mM EDTA (Gibco), as previously described (Coolen et al., 2007). The dermis was cut into small pieces using a scissor. Fibroblasts were isolated from the dermal tissue after 2 h incubation with a 1:1 mixture of 0.25% (wt/v) collagenase and dispase, as previously described (Van den Bogaerdt et al., 2002). Keratinocytes were cultured in defined keratinocyte serum free medium (Gibco) supplemented with 1% (w/v) penicillin/streptomycin (P/S; Gibco); further referred to as keratinocyte medium. Fibroblasts were cultured in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% (v/v) fetal calf serum (FCS; Gibco), 1% (v/v) P/S and 1% Glutamax (Gibco); further referred to as fibroblast medium. Cells were seeded (12000 cells/ cm2) onto tissue culture plastic (fibroblasts) or tissue culture plastic coated with 1 µg/cm2 collagen type IV ((Merck KGaA) (keratinocytes) (Coolen et al., 2007). Cells were cultured at 37 °C and 5% CO2 until 80-90% confluency was reached.
Ex vivo human skin models
Human skin was obtained from patients undergoing elective abdominal dermo-lipectomy. Skin grafts with a thickness of 0.8 mm were prepared using a dermatome (Aesculap AG & Co. KG, Tuttlingen, Germany). Grafts were stored in Roswell Park Memorial Institute (RPMI) 1640 medium (Gibco) supplemented with 2% (v/v) P/S (Gibco) at 4°C for a maximum of two weeks. Prior to the experiment, the grafts were washed in RPMI 1640 medium at 4°C overnight, followed by PBS for approximately 2 h at room temperature to ensure the removal of P/S.
Ex vivo excision wound models (EWMs) were prepared from skin grafts using a dermatome (width 7 mm) to remove 0.3 mm of the upper part of the skin graft containing the epidermis. Subsequently, the grafts were cut into pieces of 1 ´ 1.5 cm using a scalpel (Dijksteel et al., 2019a). To prepare burn wound models (BWMs) the skin graft was cut into pieces of 1 ´ 1 cm using a scalpel and a burn was inflicted on the skin piece by placing a soldering iron (10 ´ 2 mm) set at 95°C on the epidermis for 10 s without exerting pressure.
Bacterial culture
A clinical isolate of MRSA (LUH14616; Haisma et al., 2014)was cultured in Luria Bertani (LB) medium at 37 °C and shaken at 200 rpm for approximately 4 h. The bacterial culture was centrifuged at 3600 ´ g for 5 min and the pellet was re-suspended in PBS to a desirable concentration, based on the optical density at 600 nm.
In vitro killing assay
MRSA or SAAP-148 were suspended in PBS or PBS containing 50% (v/v) blood plasma, 50% (v/v) eschar extract, 3% (v/v) bovine serum albumin (BSA; Thermo Fisher Scientific, Paisley, UK), 90% (v/v) DMEM (Gibco) or precipitated proteins from 50% (v/v) blood plasma or eschar extract. SAAP-148 was serially diluted 2-fold (0-30 nmol) and mixed with MRSA at a final concentration of 105 colony forming units (CFU)/ml. The final mixtures of 100 µl were briefly vortexed and incubated at 37°C. After 30 min of incubation, the samples were processed as previously described (Dijksteel et al., 2019b).
Ex vivo efficacy test
Ex vivo EWMs were inoculated with 10 µl of 107 CFU/mL MRSA for 1 h and topically exposed to 20 µL of PBS containing 0, 15, 30 and 60 nmol SAAP-148 for 1 h at 37 °C and 5% CO2. The models were transferred to polypropylene vials containing a 7-mm metal bead and 1 ml of 0.05% (wt/v) sodium polyanethol sulfonate (SPS; Merck KGaA, Darmstadt, Germany) in PBS for homogenization using the TissueLyser set at 50 Hz for 4 min. Ten-fold serial dilutions of the homogenates were cultured on LB agar plates to quantify the number of viable bacteria after an overnight incubation at 37°C and 5% CO2.
Cytotoxicity assay
In vitro
Human fibroblasts or keratinocytes were seeded (80,000/cm2) in 96-well plates at 37°C and 5% CO2. After 18 h, medium was discarded and cells were washed using PBS. Subsequently, 90 µl of DMEM followed by 10 µl of 2-fold serially diluted SAAP-148 in PBS (0-30 nmol) or 10% (v/v) Triton X-100 (Thermo Fisher Scientific) was added to the cells. After 1 h incubation at 37°C and 5% CO2, the level of lactate dehydrogenase (LDH) in the media was determined as a measure for cytotoxicity using the non-radioactive cytotoxicity assay kit (Promega Corporation, Madison, USA), according to the manufacturer’s instructions (Promega Corporation, n.d.). The absorbance of the samples was measured on a SpectraMax M2 (Molecular Devices, California, USA) at 490 nm. To determine the maximum LDH release, 10 µl of 10% (v/v) Triton X-100 was added to the surviving cells and the level of LDH was measured. The level of cytotoxicity was calculated as follows: [(experimental LDH release – spontaneous LDH release)/ (maximum LDH release – spontaneous LDH release)] ´ 100%.
Ex vivo
EWMs were exposed to 20 µL of PBS containing 0, 15, 30 and 60 nmol SAAP-148 or as negative control to 20 µl of 1% (v/v) Triton X- 100 for 1 h at 37°C and 5% CO2. Subsequently, the models were washed in PBS and fixed in 1% (wt/v) paraformaldehyde in PBS for 2 h at 4°C, followed by an overnight incubation in 20% (wt/v) sucrose solution at 4°C while rotating. The samples were embedded in Tissue Tek O.C.T. compound (Sakura Finetek, Torrance, USA) to prepare 10 µm sections for LDH staining as previously described (Karim et al., 2019). LDH-stained cells were microscopically identified as a measurement for viable cells. To determine the level of cytotoxicity, we measured the area in the wound without LDH-stained cells using NIS Elements (Nikon Instruments Europe B.V., Amstelveen, the Netherlands).
Ex vivo wound healing
Ex vivo wound models were placed dermis down on stainless-steel grids for air-exposed culture at 37°C and 5% CO2, as previously described (Coolen et al., 2008). The models were exposed to 20 µl of PBS containing 0, 15, 30 and 60 nmol SAAP-148, or 1% (wt/wt) SSD in cetomacrogol cream (Pharmacy of the Medical Center Alkmaar, Alkmaar, the Netherlands), twice a week. EWMs were cultured for two weeks and BWMs were cultured for three weeks to determine the effect of the peptide on re-epithelialization. Prior to exposure to SAAP-148, SSD or PBS, the culture medium was refreshed. Twenty-four hours prior to fixation of the ex vivo models in kryofix (50% ethanol and 3% PEG300), 20 µM 5-bromo-2-deoxyuridine (BrdU) was added to the culture medium.
Immunohistochemistry
Samples were processed for paraffin embedding and 5 µm sections were deparaffinized and rehydrated for HE and BrdU (B5002; Merck KGaA) staining, as previously described (Dijksteel et al., 2020). The newly formed epidermis (outgrowth) and the number of BrdU-positive cells/mm in the wounded area were microscopically determined using NIS Elements.
Statistical analysis
Statistical analysis was performed using SPSS version 24. For differences between groups the Kruskal-Wallis test followed by the Mann-Whitney-U test was used. To compare two related groups, the Wilcoxon signed rank sum test was used. Values of p<0.05 are considered significantly different.
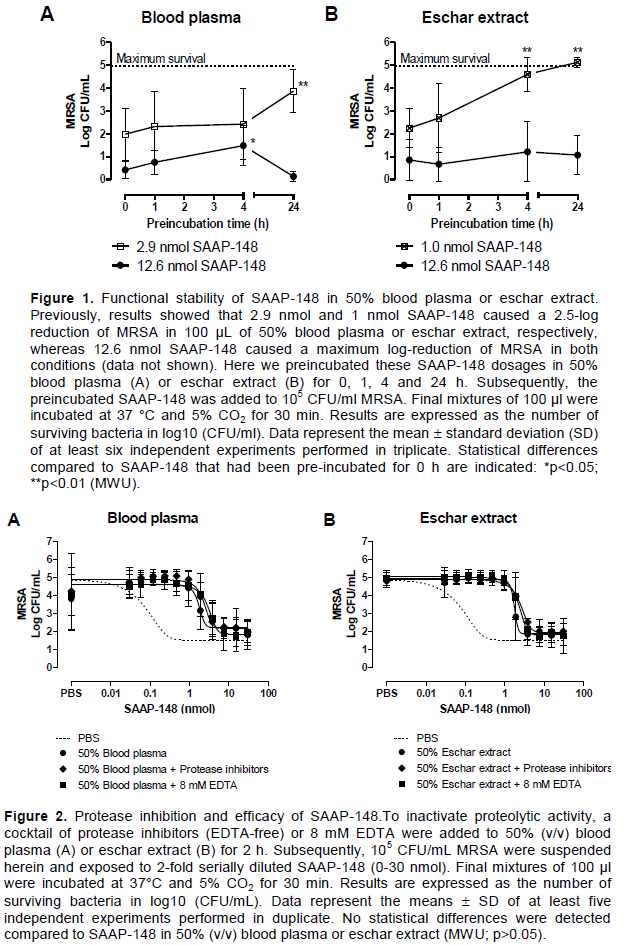
Functional stability of SAAP-148 in 50% (v/v) blood plasma and eschar extract
Approximately 2.9 and 1 nmol SAAP-148 caused a 2.5-log reduction of MRSA in 100 µl of 50% (v/v) blood plasma and eschar extract, respectively, whereas 0.2 nmol SAAP-148 was sufficient to reach a 2.5-log reduction of MRSA in PBS environment (data not shown). To maximally reduce a bacterial count of 105 CFU/ml in 50% blood plasma and eschar extract, approximately 12.6 nmol SAAP-148 was necessary (data not shown). To determine if the bactericidal activity of SAAP-148 was dependent on the time that it resided in the biological environment, SAAP-148 was preincubated in 50% (v/v) blood plasma or eschar extract at a high (12.6 nmol) and low dosage (2.9 or 1 nmol). A 24 h preincubation of SAAP-148 at low dosages, capable to yield a 2.5-log reduction of MRSA, in 100 µl of 50% (v/v) blood plasma or eschar extract reduced the bactericidal efficacy of SAAP-148 considerably (Figure 1). In contrast, a 24 h preincubation period of 12.6 nmol SAAP-148 in 50% (v/v) blood plasma or eschar extract did not affect the bactericidal efficacy.
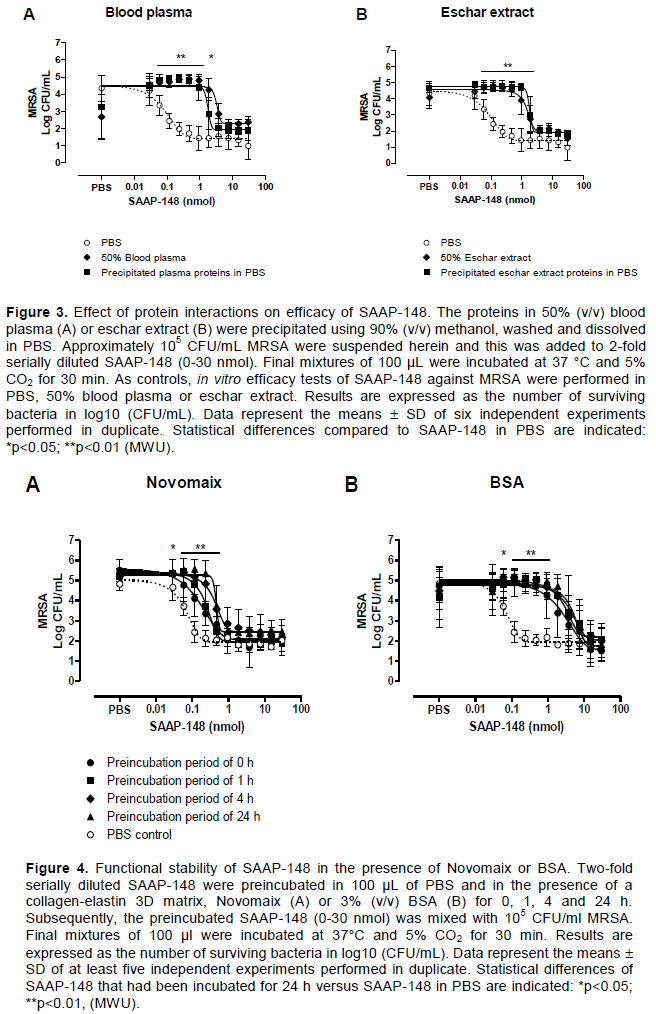
Effect of protease inhibition on SAAP-148’s stability
The necessity for higher concentrations of SAAP-148 to reach equivalent bactericidal activity in biologically relevant environments as compared to PBS might be explained by proteolytic degradation of SAAP-148. Therefore, in vitro efficacy tests of SAAP-148 against MRSA were performed using 50% (v/v) blood plasma or eschar extract that had been incubated for 2 h with a cocktail of protease inhibitors or 8 mM EDTA to inhibit metalloproteases. Inhibition of proteolytic activity did not affect the bactericidal efficacy of SAAP-148 against MRSA in blood plasma and eschar extract (Figure 2).
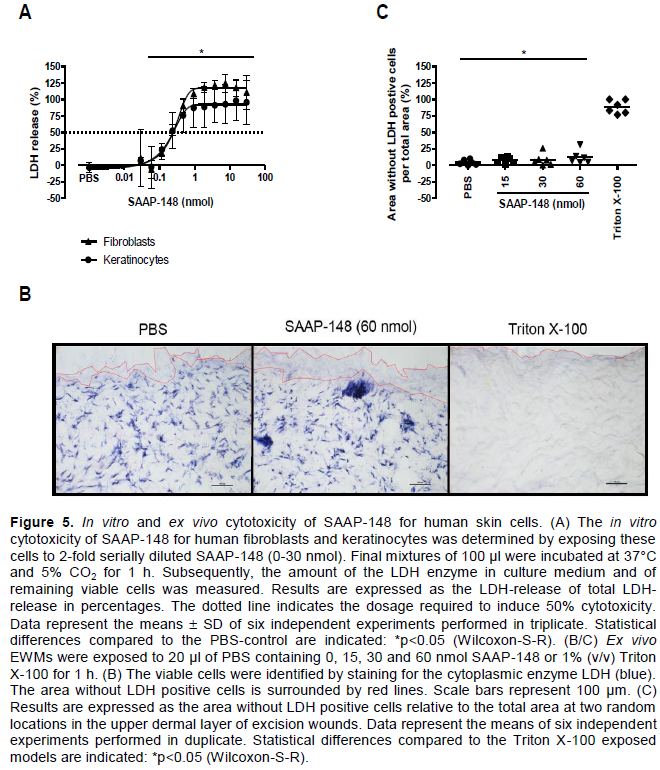
Bioactivity of SAAP-148 in relevant environments for wound healing
Interactions of SAAP-148 with proteins present in 50% (v/v) blood plasma or eschar extract might reduce the bioavailability and thus affect the bactericidal efficacy of SAAP-148. Therefore, we precipitated the proteins in 50% (v/v) blood plasma or eschar extract and performed in vitro efficacy tests of SAAP-148 with the precipitated proteins dissolved in PBS. To achieve a 2-log reduction of MRSA in 100 µl of suspension, >20-fold higher dosages of SAAP-148 were required in the presence of the precipitated proteins as compared to PBS only (Figure 3). This effect is similar to the reduction found for 50% (v/v) blood plasma or eschar extract.
To assess the types of proteins that could be involved, we preincubated SAAP-148 with a collagen-elastin 3D matrix, Novomaix or 3% (v/v) BSA for 0, 1, 4 and 24 h. Subsequently, 105 CFU/mL MRSA was exposed to the preincubated SAAP-148 for 30 min. Figure 4A shows that the required dosage of SAAP-148 to induce bacterial killing in 100 µl of suspension gradually increased as the preincubation period of SAAP-148 with Novomaix increased from 0 to 24 h. An even stronger effect was seen as SAAP-148 was preincubated with 3% (v/v) BSA. This effect occurred instantly and remained until the 24 h preincubation period (Figure 4B).
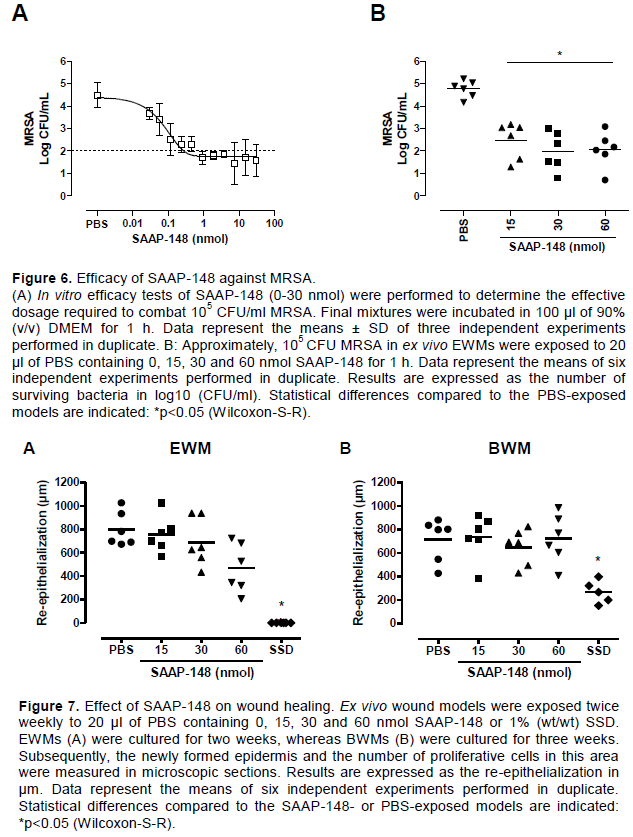
Cytotoxicity of SAAP-148 for human fibroblasts and keratinocytes
Human fibroblasts and keratinocytes in 90 µl of medium were exposed to 10 µl of PBS containing increasing dosages of SAAP-148 for 1 h. Subsequently, we measured the amount of LDH, a cytosolic enzyme present in nearly all living cells, which is released into the culture medium as a result of cell damage. Figure 5A shows that SAAP-148 induced the release of cytosolic LDH for both human fibroblasts and keratinocytes in a dose-dependent manner. Notably, a SAAP-148 dosage of only 0.23 nmol in 100 µl of 90% (v/v) DMEM resulted in 50% LDH release. To determine the level of cytotoxicity of SAAP-148 for human skin cells in their natural biological environment, ex vivo EWMs were exposed to 20 µl of PBS containing various dosages of SAAP-148 and – as positive control - Triton X-100 for 1 h. Subsequently, the viable cells were identified by staining tissue sections for LDH. SAAP-148 did not penetrate through the whole thickness of the skin sample (data not shown) and therefore the level of cytotoxicity was determined using the area in the sample without LDH-stained cells. Unlike Triton X-100, SAAP-148 dosages up to 60 nmol/model did not eliminate the LDH containing cells in the excision wounds (Figure 5B and C).
To determine the therapeutic index of SAAP-148, the bactericidal efficacy of this peptide was determined at the same conditions as the cytotoxicity assays. SAAP-148 eradicated MRSA in a dose-dependent manner in vitro and approximately 0.23 nmol SAAP-148 in 100 µl of medium resulted in a 3-log reduction of MRSA (Figure 6A). In ex vivo EWMs, SAAP-148 was equally potent against MRSA at 15, 30 and 60 nmol/model (Figure 6B). Approximately 15 nmol of SAAP-148 was sufficient to achieve a 3-log reduction of MRSA. Hence, SAAP-148 was effective against MRSA at the same dosage that induced cytotoxicity in 2D culture, resulting in a therapeutic index of 1. In contrast, the lowest tested dosages of 15 nmol SAAP-148 effectively eradicated MRSA in ex vivo excision wound models, whereas the highest tested dosage of 60 nmol SAAP-148 was not sufficient to induce 50% cytotoxicity ex vivo. This indicates a therapeutic index of at least 4.
Effect of SAAP-148 on wound healing
To determine the effect of SAAP-148 on wound healing, we exposed ex vivo EWMs and BWMs to 20 µL of PBS containing 0, 15, 30 and 60 nmol SAAP-148 or 1% (wt/wt) SSD. Subsequently, we measured the newly formed epidermis (re-epithelialization) and the number of proliferative cells in that area. A SAAP-148 dose of 60 nmol/model showed no statistically significant negative effects on the re-epithelialization of EWMs (Figure 7A) and BWMs (Figure 7B). In addition, SAAP-148 did not affect the number of proliferating cells in the EWMs and BWMs (data not shown). Contrarily, exposure to SSD resulted in little to no re-epithelialization and proliferating cells in both types of wound models.
In the current study, we tried to elucidate why higher concentrations of SAAP-148 in 50% (v/v) blood plasma or eschar extract are necessary to reach a similar bactericidal effect as in PBS (Dijksteel et al., 2019a). We have shown that preincubation of SAAP-148 in these biological relevant environments significantly reduced the bactericidal efficacy of this peptide against MRSA. This suggested a reduced bioavailability of SAAP-148 that could have been caused by proteolytic degradation and/or binding of SAAP-148. The bactericidal efficacy of several AMPs including LL-37 is reduced by proteolytic activity in the presence of blood plasma (McCrudden et al., 2014; Sieprawska-Lupa et al., 2004; Thwaite et al., 2006). Previously, naturally occurring and synthetic AMPs were found to be susceptible to protease activity (Moncla et al., 2011), whereas other AMPs displayed a reduced bioactivity due to interactions with albumin or NaCl (Björn et al., 2016; Svenson et al., 2007). Similar to some synthetic AMPs (Shin et al., 2010), SAAP-148 was not degraded by proteases present in blood plasma or eschar extract.
The reduced bioactivity of SAAP-148 seemed related to protein interactions, resulting in a reduced bioavailability of SAAP-148 to combat MRSA.
Our findings are in agreement with those reported by de Breij et al. (2018) demonstrating a concentration dependent reduction in the bactericidal efficacy of SAAP-148 in the presence of high density lipoproteins or human serum albumin.
It is striking that the presence of BSA instantly reduced the bactericidal efficacy of SAAP-148 a 100-fold, whereas a 24 h incubation period of SAAP-148 with the collagen-elastin 3D matrix, Novomaix, was required to significantly reduce the bactericidal efficacy of SAAP-148. This suggests that the presence of blood-derived rather than skin-derived proteins within the eschar extract was predominantly responsible for the reduced bactericidal efficacy of SAAP-148. In addition, the discrepancy between instant albumin response and the slower plasma response on the reduction of the bactericidal effect might be explained by the presence of other proteins in blood plasma. Hence, blood protein interactions with SAAP-148 may have reduced the bioavailability of the peptide and this remains a challenge for the development of SAAP-148 as a topical antimicrobial agent for burns.
The last decade, the bactericidal efficacy of AMPs against pathogens has improved but their effects on mammalian cells have not been fully investigated. Therefore, we included cytotoxicity studies of SAAP-148 for human skin cells. SAAP-148 did display some cytotoxic effects on human fibroblasts and keratinocytes in in vitro cultures but not in ex vivo skin. This suggests that in 3D skin, cells are protected by the extracellular matrix. For a set of different AMPs, the in vitro therapeutic index was determined, whereby the highest therapeutic index was found for cecropins and the smallest for melittin (Bacalum and Radu, 2015). Based on our findings, the in vitro therapeutic index of SAAP-148 was lower than for melittin. Nonetheless, the therapeutic index of SAAP-148 for ex vivo human skin models was at least 4-times higher than for in vitro skin cells. Hence, the difference between the in vitro and ex vivo findings highlights the importance to evaluate the bactericidal efficacy and safety of AMPs such as SAAP-148 in models that better simulate the clinical situation.
Several AMPs such as LL-37, OA-GL12 and GE33 have the ability to promote wound healing (Huang et al., 2014; Ramos et al., 2011; Song et al., 2019; Steinstraesser et al., 2008). Therefore, we determined the effect of SAAP-148 on wound healing. SAAP-148 showed no negative effects on the re-epithelialization of ex vivo excision wounds and burn wounds. The ‘gold standard’ for treatment of burn wound infections, SSD, induced cellular toxicity, which resulted in a delayed wound closure. Since SAAP-148 had no effects on wound healing while it was effective against resistant bacteria, SAAP-148 could be a potential alternative for SSD as a topical antimicrobial strategy for burns.
To improve the bioavailability of AMPs, different approaches can be used such as the introduction of D-amino acids into the AMPs’ amino acid sequence, cyclization, amidation or acetylation of the thermal regions (Chu et al., 2013; Gentilucci et al., 2010; Strömstedt et al., 2009). These approaches, however, are designed to improve the stability of AMPs against proteolytic degradation. Chemical modifications, such as tryptophan end-tagging, the introduction of the non-natural amino acid b-naphtlylalanine or the replacement of tryptophans and histidines in Trp- and His-rich AMPs could result in an improved salt and serum stability (Chu et al., 2013; Pfalzgraff et al., 2018). Also, amino acid substitution to decrease the net positive charge of the peptide within a certain range could result in less cytotoxicity without effecting the antimicrobial efficacy (Jiang et al., 2008; Xu et al., 2020).
Alternatively, SAAP-148 might be protected against unwanted protein and cell binding using drug delivery systems, such as hydrogels, polylactic-co-glycolic acid nanoparticles or lipid-based encapsulation systems (Meikle et al., 2017; Xie et al., 2015). Formulation of SAAP-148 might not only result in an improved bioavailability of the peptide (Haisma et al., 2016), but also in an improved delivery of the AMPs to deeper layers of the skin.
In the presence of blood plasma, blood proteins or skin proteins, higher concentrations of SAAP-148 were required to reach the same bactericidal effects as in PBS. The reduced bioavailable concentrations of SAAP-148 were not caused by proteolytic degradation but presumably by protein binding. Furthermore, in contrast to isolated human skin cell cultures, relatively high concentrations of SAAP-148 did not display cytotoxic effects in ex vivo excision wounds and burn wounds and allowed undisturbed epithelialization while retaining bactericidal activity.
The authors have not declared any conflict of interests.
AMPs, antimicrobial peptides; BrdU, 5-bromo-2- deoxyuridine; BSA, bovine serum albumin; CFU, colony forming units; DMEM, Dulbecco's Modified Eagle Medium; DNA, deoxyribonucleic acid; EDTA, ethylenediaminetetraacetic acid; EWM, excision wound model; FCS, fetal calf serum; LB, Luria Bertani; LDH, lactate dehydrogenase; MRSA, methicillin resistant Staphylococcus aureus; PBS, phosphate-buffered saline; P/S, penicillin/streptomycin; RPMI, Roswell Park Memorial Institute; SD, standard deviation; SPS, sodium polyanethol sulfonate; SSD, silver sulfadiazine.
The authors appreciate Nourhan Shehata and Esmée Alibux for assisting in the experiments during an internship at the Association of Dutch Burn Centers, (Beverwijk, the Netherlands). This collaborative project was funded by the Ministry of Economic Affairs through two public–private partnership (PPP) allowances, made available by Health-Holland and Top Sector life Sciences and Health. One of the PPP allowances was co-funded by the Dutch Burns Foundation, Madam Therapeutics B.V., Avivia B.V., Leiden University Medical Center, Amsterdam University Medical Center and the Association of Dutch Burn Centers (LSHM17078-SGF) and the other PPP allowance was co-funded by the Dutch Burns Foundation, Madam Therapeutics B.V., Mölnlycke Health Care AB, Leiden University Medical Center, Amsterdam University Medical Center and the Association of Dutch Burn Centers (LSH-TKI40-43100-98-017).
REFERENCES
|
Atiyeh BS, Costagliola M, Hayek SN, Dibo SA (2007). Effect of silver on burn wound infection control and healing: Review of the Literature Burns 33(2):139-148.
Crossref
|
|
|
|
Bacalum M, Radu M (2015). Cationic antimicrobial peptides cytotoxicity on mammalian cells: An analysis using therapeutic index integrative concept. International Journal of Peptide Research and Therapeutics 21(1):47-55.
Crossref
|
|
|
|
|
Beisswenger C, Bals R (2005). Functions of antimicrobial peptides in host defense and immunity. Current protein & peptide science 6(3):255-264.
Crossref
|
|
|
|
|
Björn C, Mahlapuu M, Mattsby-Baltzer I, Håkansson J (2016). Anti-infective efficacy of the lactoferrin-derived antimicrobial peptide HLR1r. Peptides 81:21-28.
Crossref
|
|
|
|
|
Van den Bogaerdt AJ, Van Zuijlen PPM, Van Galen M, Lamme EN, Middelkoop E (2002). The suitability of cells from different tissues for use in tissue-engineered skin substitutes. Archives of Dermatological Research 294(3):135-142.
Crossref
|
|
|
|
|
de Breij A, Riool M, Cordfunke RA, Malanovic N, de Boer L, Koning RI, Ravensbergen E, Franken M, van der Heijde T, Boekema BK, Kwakman PHS, Kamp N, El Ghalbzouri A, Lohner K, Zaat SAJ, Drijfhout JW, Nibbering PH (2018). The antimicrobial peptide SAAP-148 combats drug-resistant bacteria and biofilms. Science Translational Medicine 10(423):eaan4044.
|
|
|
|
|
Chu HL, Yu HY, Yip BS, Chih YH, Liang CW, Cheng HT, Cheng JW (2013). Boosting salt resistance of short antimicrobial peptides. Antimicrobial Agents and Chemotherapy 57(8):4050-4052.
Crossref
|
|
|
|
|
Coolen NA, Verkerk M, Reijnen L, Vlig M, van den Bogaerdt AJ, Breetveld M, Gibbs S, Middelkoop E, Ulrich MMW (2007). Culture of keratinocytes for transplantation without the need of feeder layer cells. Cell transplantation 16(6):649-661.
Crossref
|
|
|
|
|
Coolen NA, Vlig M, Van Den Bogaerdt AJ, Middelkoop E, Ulrich MMW (2008). Development of an in vitro burn wound model. Wound Repair and Regeneration 14(4):559-567.
Crossref
|
|
|
|
|
Dijksteel GS, Ulrich MMW, Vlig M, Nibbering PH, Cordfunke RA, Drijfhout JW, Middelkoop E, Boekema BKHL (2019a). Potential factors contributing to the poor antimicrobial efficacy of SAAP-148 in a rat wound infection model. Annals of Clinical Microbiology and Antimicrobials 18(1):38.
Crossref
|
|
|
|
|
Dijksteel GS, Nibbering PH, Ulrich MMW, Middelkoop E, Boekema BKHL (2019b). SPS-neutralization in tissue samples for efficacy testing of antimicrobial peptides. BMC Infectious Diseases 19(1):1093.
Crossref
|
|
|
|
|
Dijksteel GS, Ulrich MMW, Vlig M, Sobota A, Middelkoop E, Boekema BKHL (2020). Safety and bactericidal efficacy of cold atmospheric plasma generated by a flexible surface Dielectric Barrier Discharge device against Pseudomonas aeruginosa in vitro and in vivo. Annals of Clinical Microbiology and Antimicrobials 19(1):37.
Crossref
|
|
|
|
|
Gentilucci L, De Marco R, Cerisoli L (2010). Chemical modifications designed to improve peptide stability: Incorporation of non-natural amino acids, pseudo-peptide bonds, and cyclization. Current Pharmaceutical Design 16(28):3185-3203.
Crossref
|
|
|
|
|
Glik J, Kawecki M, Gaździk T, Nowak M (2012). The impact of the types of microorganisms isolated from blood and wounds on the results of treatment in burn patients with sepsis. Polish Journal of Surgery 84(1):6-16.
Crossref
|
|
|
|
|
Gordon YJ, Romanowski EG, McDermott AM (2005). A review of antimicrobial peptides and their therapeutic potential as anti-infective drugs. Current Eye Research 30(7):505-515.
Crossref
|
|
|
|
|
Gould PW, Villiger JW (1986). Clinical and bacteriological efficacy of mupirocin (Bactroban): a new topical antibiotic. The New Zealand Medical Journal 99(805):516.
|
|
|
|
|
Haisma EM, De Breij A, Chan H, Van Dissel JT, Drijfhout JW, Hiemstra PS, El Ghalbzouri A, Nibbering PH (2014). LL-37-derived peptides eradicate multidrug-resistant Staphylococcus aureus from thermally wounded human skin equivalents. Antimicrobial Agents and Chemotherapy 58(8):4411-4419.
Crossref
|
|
|
|
|
Haisma EM, Göblyös A, Ravensbergen B, Adriaans AE, Cordfunke RA, Schrumpf J, Limpens, RWAL, Schimmel KJM, den Hartigh J, Hiemstra PS, Drijfhout JW, El Ghalbzouri A, Nibbering PH (2016). Antimicrobial peptide P60.4Ac-containing creams and gel for eradication of methicillin-resistant Staphylococcus aureus from cultured skin and airway epithelial surfaces. Antimicrobial Agents and Chemotherapy 60(7):4063-4072.
Crossref
|
|
|
|
|
Huang H N, Pan CY, Chan YL, Chen JY, Wu CJ (2014). Use of the antimicrobial peptide pardaxin (GE33) to protect against methicillin-resistant Staphylococcus aureus infection in mice with skin injuries. Antimicrobial Agents and Chemotherapy 58(3):1538-1545.
Crossref
|
|
|
|
|
Jiang Z, Vasil AI, Hale JD, Hancock REW, Vasil ML, Hodges RS (2008). Effects of net charge and the number of positively charged residues on the biological activity of amphipathic α-helical cationic antimicrobial peptides. Biopolymers - Peptide Science Section 90(3):369-383.
Crossref
|
|
|
|
|
Karim AS, Yan A, Ocotl E, Bennett DD, Wang Z, Kendziorski C, Gibson ALF (2019). Discordance between histologic and visual assessment of tissue viability in excised burn wound tissue. Wound Repair and Regeneration 27(2):150-161.
Crossref
|
|
|
|
|
Kumar P, Kizhakkedathu JN, Straus SK (2018). Antimicrobial peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 8(1):4.
Crossref
|
|
|
|
|
McCrudden MTC, McLean DTF, Zhou M, Shaw J, Linden GJ, Irwin CR, Lundy FT (2014). The host defence peptide LL-37 is susceptible to proteolytic degradation by wound fluid isolated from foot ulcers of diabetic patients. International Journal of Peptide Research and Therapeutics 20(4):457-464.
Crossref
|
|
|
|
|
Meikle TG, Zabara A, Waddington LJ, Separovic F, Drummond CJ, Conn CE (2017). Incorporation of antimicrobial peptides in nanostructured lipid membrane mimetic bilayer cubosomes. Colloids and Surfaces B: Biointerfaces 152:143-151.
Crossref
|
|
|
|
|
Moncla BJ, Pryke K, Rohan LC, Graebing PW (2011). Degradation of naturally occurring and engineered antimicrobial peptides by proteases. Advances in Bioscience and Biotechnology 2(6):404-408.
Crossref
|
|
|
|
|
Nell MJ, Tjabringa GS, Wafelman AR, Verrijk R, Hiemstra PS, Drijfhout JW, Grote JJ (2006). Development of novel LL-37 derived antimicrobial peptides with LPS and LTA neutralizing and antimicrobial activities for therapeutic application. Peptides 27(4):649-660.
Crossref
|
|
|
|
|
Pfalzgraff A, Brandenburg K, Weindl G (2018). Antimicrobial peptides and their therapeutic potential for bacterial skin infections and wounds. Frontiers in Pharmacology 9:281.
Crossref
|
|
|
|
|
Promega Corporation USA (n.d.). CytoTox 96® Non-radioactive cytotoxicity assay instructions for use of product G1780.
View [Accessed 6 June 2019].
|
|
|
|
|
Ramos R, Silva JP, Rodrigues AC, Costa R, Guardão L, Schmitt F, Soares R, Vilanova M, Domingues L, Gama M (2011). Wound healing activity of the human antimicrobial peptide LL37. Peptides 32(7):1469-1476.
Crossref
|
|
|
|
|
Rooijakkers SHM, van Kessel KPM, van Strijp JAG (2005). Staphylococcal innate immune evasion. Trends in Microbiology 13(12):596-601.
Crossref
|
|
|
|
|
Shai Y (2002). Mode of action of membrane active antimicrobial peptides. Biopolymers 66(4):236-248.
Crossref
|
|
|
|
|
Shin YP, Park HJ, Shin SH, Lee YS, Park S, Jo S, Lee YH, Lee IH (2010). Antimicrobial activity of a halocidin-derived peptide resistant to attacks by proteases. Antimicrobial Agents and Chemotherapy 54(7):2855-2866.
Crossref
|
|
|
|
|
Sieprawska-Lupa M, Mydel P, Krawczyk K, Wójcik K, Puklo M, Lupa B, Suder P, Silberring J, Reed M, Pohl J, Shafer W, McAleese F, Foster T, Travis J, Potempa J (2004). Degradation of human antimicrobial peptide LL-37 by Staphylococcus aureus-derived proteinases. Antimicrobial Agents and Chemotherapy 48(12):4673-4679.
Crossref
|
|
|
|
|
Song Y, Wu C, Zhang X, Bian W, Liu N, Yin S, Yang MF, Luo M, Tang J, Yang X (2019). A short peptide potentially promotes the healing of skin wound. Bioscience Reports 39(3):BSR20181734.
Crossref
|
|
|
|
|
Steinstraesser L, Koehler T, Jacobsen F, Daigeler A, Goertz O, Langer S, Kesting M, Steinau H, Eriksson E, Hirsch T (2008). Host defense peptides in wound healing. Molecular Medicine 14(7-8):528-537.
Crossref
|
|
|
|
|
Strömstedt AA, Pasupuleti M, Schmidtchen A, Malmsten M (2009). Evaluation of strategies for improving proteolytic resistance of antimicrobial peptides by using variants of EFK17, an internal segment of LL-37. Antimicrobial Agents and Chemotherapy 53(2):593-602.
Crossref
|
|
|
|
|
Svenson J, Brandsdal BO, Stensen W, Svendsen JS (2007). Albumin binding of short cationic antimicrobial micropeptides and its influence on the in vitro bactericidal effect. Journal of Medicinal Chemistry 50(14):3334-3339.
Crossref
|
|
|
|
|
Thwaite JE, Hibbs S, Titball RW, Atkins TP (2006). Proteolytic degradation of human antimicrobial peptide LL-37 by Bacillus anthracis may contribute to virulence. Antimicrobial Agents and Chemotherapy 50(7):2316-2322.
Crossref
|
|
|
|
|
Williamson DA, Carter GP, Howden BP (2017). Current and emerging topical antibacterials and antiseptics: Agents, action, and resistance patterns. Clinical Microbiology Reviews 30(3):827-860.
Crossref
|
|
|
|
|
Xie Z, Aphale NV, Kadapure TD, Wadajkar AS, Orr S, Gyawali D, Qian G, Nguyen KT, Yang J (2015). Design of antimicrobial peptides conjugated biodegradable citric acid derived hydrogels for wound healing. Journal of Biomedical Materials Research 103(12):3907-3918.
Crossref
|
|
|
|
|
Xu L, Shao C, Li G, Shan A, Chou S, Wang J, Ma Q, Dong Na (2020). Conversion of broad-spectrum antimicrobial peptides into species-specific antimicrobials capable of precisely targeting pathogenic bacteria. Scientific Reports 10(1):944.
Crossref
|
|
|
|
|
Yin LM, Edwards MA, Li J, Yip CM, Deber CM (2012). Roles of hydrophobicity and charge distribution of cationic antimicrobial peptides in peptide-membrane interactions. Journal of Biological Chemistry 287(10):7738-7745.
Crossref
|
|
|
|
|
Zasloff M (2002). Antimicrobial peptides of multicellular organisms. Nature 415(6870):389-395.
Crossref
|
|