Full Length Research Paper

ABSTRACT
Zika virus (ZIKV), a mosquito-borne member of the family Flaviviridae, is a human pathogen of global significance. Recently, ZIKV, has become a public health problem with increases in numbers of cases and a strong association between ZIKV outbreaks and the spread of cases of Guillain-Barré Syndrome and microcephaly. In this study, the extracts of the seaweed Osmundaria obtusiloba (O. obtusiloba) (native to the Brazilian coast) against ZIKV using Vero cells was evaluated. The seaweed extract tested inhibited ZIKV replication in a dose-dependent manner at low concentrations with EC50 values of 1.82 μg/mL and a selective index (SI) of 288. Other results showed that this extract had significant virucidal effects. In addition, when the extract and Ribavirin were used concomitantly there was a significant synergistic effect. Our promising results suggest that extracts of O. obtusiloba are excellent candidates for further studies, and that marine algae are potentially important sources for the development of novel anti-ZIKV agents.
Key words: ZIKA, seaweeds, antiviral activity, marine algae, Osmundaria obtusiloba.
INTRODUCTION
The antiviral potential of marine macroalgae is widely recorded. Several studies have demonstrated activity of a number of species of algae against HIV-1 (Cirne-Santos et al., 2008; Barros et al., 2016), HSV-1 (Macedo et al., 2012; Mendes et al., 2012; Soares et al., 2012; Barros et al., 2015), HSV-2 (Mendes et al., 2012; Soares et al., 2012) and dengue type 2 strains (Talarico et al., 2007). Thus, marine algae provide a potentially rich source for the discovery of antiviral drugs. In this study, the native algal extract of Osmundaria obtusiloba (C.Agardh) R.E.Norris, from the Brazilian coast, was tested for inhibiting the replication of the Zika virus (ZIKV) (Hayashi et al., 2007).
Arthropod-borne viruses, commonly called arboviruses, normally circulate in nature through biological transmission between susceptible vertebrate hosts and blood-feeding arthropods, such as mosquitoes. Studies show that the most important mosquitoes in this transmission are A. aegypti, although there is also strong evidence for the role of A. albopictus in this process too (Kraemer et al., 2015; Calvez et al., 2016).
The main arboviruses causing disease in humans include the alphaviruses (Togaviridae: Alphavirus), flaviviruses (Flaviviridae: Flaviviruses), Bunyaviruses (Bunyaviridae) and some members of other virus families (Rhabdoviridae and Reoviridae). Currently, of the 534 viruses listed in the International Catalogue of Arboviruses, 214 are known to be, or are probably associated with arthropods, 287 viruses are reported to be possible arboviruses, and 33 are probably or definitely not arboviruses (Gubler, 2001; Iranpour et al., 2016). In total, 134 of the 534 arboviruses have been reported to cause disease in humans and have a global distribution with the majority circulating in tropical areas, where climatic conditions favor transmission throughout the year (Gyawali et al., 2016; Tabachnick, 2016).
Zika virus (ZIKV) is a mosquito-borne and from the genus Flavivirus, family Flaviviridae and clusters with the Spondweni serocomplex (Vorou, 2016). Flaviviruses have a positive sense single-strand RNA genome of approximately 11,000 nucleotides in length. The genome contains a long open-reading frame (ORF) that encodes three structural proteins (capsid, precursor membrane and envelope) that form the viral particles and seven non-structural proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5). The non-structural proteins participate in viral replication, virion assembly, and evasion of the host immune response (Lindenbach et al., 2013). The infection in humans produces a self-limiting acute febrile illness with fever, headache, myalgia and rash, very similar to other arboviruses like Dengue Virus and Chikungunya. Therefore, in regions where more than one arbovirus is detected, ZIKV could be circulating but not be notified causing misleading and low incidences recorded (Tappe et al., 2015; Vorou, 2016).
After the ZIKV was first detected in 1947, during a yellow fever surveillance program in Uganda, few reports of the disease were recorded until 2007 when there was a large outbreak in Yap Island Federal Sate of Micronesia (Faye et al., 2014; Hamel et al., 2016). In October of 2013, a large outbreak occurred in French Polynesia where 396 laboratory confirmed cases were reported. Up to now, two main distinct ZIKV geographic lineages have been described (African and Asian) (Musso et al., 2014; Kucharski et al., 2016).
A substantial proportion of ZIKV infections are subclinical, but when clinical symptoms occur, the disease produced was considered moderate and self-limiting. Recent studies, however, have described a strong association between ZIKV outbreak and an increased number of cases of Guillain-Barré Syndrome (GBS) in French Polynesia, indicating a first complication resulting from a ZIKV infection (Cao-Lormeau et al., 2016; Teixeira et al., 2016). There is also strong evidence for the incidence of cases of microcephaly following ZIKV infection of women during pregnancy. These observations have been supported by evidence accrued during different outbreaks and resulted in great fear in pregnant women (Mlakar et al., 2016; Rodrigues, 2016). Other factors associated with ZIKV infection, such as hyperglycemia, among other malignancies are clear demonstrations of the a potent morbidity of this virus (Nielsen and Bygbjerg, 2016).
In May 15, 2015, the Ministry of Health of Brazil confirmed ZIKV circulation in the country after ZIKV identification in 16 samples (eight from Bahia and eight from Rio Grande do Norte) by the National Reference Laboratory. The symptoms that were described as are the most common include, arthralgia, edema of the extremities, slight fever, headache, retro-orbital pain, conjunctival hyperemia and maculopapular rashes, often spreading down the face to the limbs and often itchy, dizziness, myalgia and digestive disorders.(Junior et al., 2015; Heukelbach et al., 2016).
There is no vaccine or specific antiviral therapy for the prevention or treatment of infections by ZIKV (Barrows et al., 2016). A study identified the viral polymerase inhibitor, 7DMA, as an inhibitor of in vitro ZIKV replication, and, in virus-infected mice significantly reduced viremia and delayed virus-induced morbidity and mortality (Zmurko et al., 2016). Deng et al. (2016) also showed that an adenosine analog has in vitro and in vivo activity against ZIKV.
Previous studies have shown different biological activities for O. obtusiloba extract such as: In vivo tests using BALB/c mice infected with L. amazonenses in the control of the dissemination of this parasite (Lira et al., 2016); acute toxicity tests have demonstrated that O. obtusiloba extract does not produce significant toxic effects in BALB/c mice (de Souza Barros et al., 2018); and studies have shown that compounds derived from O. obtusiloba showed potent antiviral activity against HSV-1 and HSV-2 and had low toxicity to cell cultures (de Souza et al., 2012). In this way, these studies reinforced our perspectives for the accomplishment of this work.
Characterization of the antiviral activity of the crude extract of the algae O. obtusiloba against ZIKV was done in this work. It is shown that the extract inhibited ZIKV replication and thus our findings broaden the antiviral scope.
MATERIALS AND METHODS
Seaweed material and extraction
The seaweed O. obtusiloba, is a native species of Brazil and was collected by snorkeling at a depth 1-3 m in various coastal sites in Rasa Beach, Armação de Búzios, Rio de Janeiro State (lat. 22° 45’40”, long. 41° 54’ 32”). The seaweed was separated from sediments, epiphytes, and other associated organisms, washed with sea water and air-dried (approximate ca. temperature 28-30°C for 7-10 days) until the total evaporation of any water. Air-dried seaweed (approximately 100 g) was powdered and exhaustively extracted three times using ethanol for 72 h in the approximate temperature of 28 to 30ºC. The extract was evaporated under reduced pressure, yielding crude extract (15 to 20 mg), of which 2 to 5 mg was used in tests against the ZIKV. The ethanolic extract was chosen due to the efficiency of ethanol in extracting the phenolic compounds from O. obtusiloba (Carvalho et al., 2006)and also the low toxicity in vivo of this extract (de Souza Barros et al., 2018).
Cells and virus
Vero cells (African green monkey kidney) were grown in Dulbecco’s Modified Eagle Medium (DMEM; Invitrogen, cat. no 11960) supplemented with 5% Fetal Bovine Serum (FBS; Invitrogen), 2 mmol/L L-glutamine (Invitrogen, cat. no. 25030). Antibiotics were added at a final concentrations of 50 units/ml penicillin and streptomycin (Invitrogen, cat. no. 15070). ZIKV (ATCC® VR-1839™) was amplified in C6/36 mosquito cells line from A. albopictus, adapted to grow at 28°C, was cultured in L-15 Medium (Leibovitz) supplemented with 0.5% tryptose phosphate broth, 0.03% glutamine, 1% MEM non-essential amino acids solution and 5% FBS.
Cellular cytotoxicity assays
To evaluate the cytotoxic effect of the seaweed extract, VERO cells were cultured in 96-well plates to 90% confluence. The cells were then treated with increasing concentrations (25, 50, 100, 200, 400 and 800 μg/ml) of the crude extracts of algae and incubated for 2 to 3 days in DMEM culture medium with 5% FBS at 37°C in a 5% CO2 atmosphere. For assessment of cell viability, the MTT method was used as previously described by Mosmann (1983). In the 96-well plate previously treated with the extracts, the MTT reagent [3-(4,5-dimethylthiazole-2-yl)-2,5-diphenyltetrazolium bromide] Sigma-Aldrich) was added at a concentration of 5 mg/ml and incubated for 3 h at 37°C. The MTT medium was then removed and 100 μM dimethylsulfoxide (DMSO) was added to the plate and incubated for 15 min to dissolve the formazan crystals (Mosmann, 1983).. The plate was read in an ELISA reader at 550 nm absorbance. The percentage of metabolically active cells was compared to the control of extract untreated cells, to determine the cytotoxicity of the compounds. These assays were performed three times independently, each in triplicate.
Plaque reduction assay
VERO cells were cultured in DMEM medium and after confluence were incubated with the ZIKV (MOI of 0.1) for 2 h. Subsequently, the cells were washed with PBS to remove the residual virus and a 2% (w/v) mixture of CMC (Sigma Aldrich) was added with DMEM supplemented with 5% FBS, 5 mmol/L L-glutamine and 0.20% of sodium bicarbonate. Different concentrations of the seaweed extracts were added and incubated at 37°C in a 5% CO2 atmosphere and analyzed daily for plaque formation. Subsequently, the cells were fixed with 10% formaldehyde, then stained with 1% crystal violet and the plates were examined and plaque formation quantified. The assay was used for evaluation of antiviral activity and for viral titration. The infectious virus titer (PFU/mL) was determined using the following formula: plate count × dilution factor × (1 / inoculation volume).
Antiviral assay
Antiviral activity was evaluated using a virus plaque reduction assay. Vero cells were grown in 24-well plates under conditions described above and subsequently infected with ZIKV (MOI of 0.1) in the absence or presence of different concentrations of the crude seaweed extracts ranging from 1.25, 2.5, 5, 10, 15, 20, 25 or 50 μg/mL, respectively. After 1 h of adsorption at 37°C, residual inoculum was replaced by medium containing 2% methyl-cellulose and the corresponding dose of each extract. Plates were evaluated daily and counted between 5 to 10 days of incubation at 37°C in 5% CO2. The 50% inhibitory concentration (EC50) was calculated as the extract concentration required reducing the virus plaques by 50%. All experiments were performed twice and each in triplicate.
Viral kinetics and time-of-drug addition studies
Vero cells were cultured in 24 well plates, as above and after 90% confluence, was treated differentially. In some wells, the cells were pretreated with the crude extract at 5 μg/mL, from O. obtusiloba for 1, 2 or 3 h prior to infection. Subsequently, these cells were incubated with ZIKV (MOI of 0.1) while in other wells cells were incubated at time 0 (immediately after infection) or at 1, 2 and 3 h post-infection with the extract concentration of 5 μg/mL. Cells were then maintained under the conditions for the plaque assay production at 37°C in 5% in CO2. Inhibition of viral replication was evaluated in relation to the control cells, incubated without extract in 3 independent experiments in triplicate.
Virucidal effect
A suspension of ZIKV, containing the relative concentration which in culture corresponds to an MOI of 0.1, was incubated with the same volume of algae extract from O. obtusiloba at concentrations of 2.5, 5 or 10 μg/mL and incubated in microtiter plates for 2 h at 37°C. The pre-incubated samples were then added to Vero cells in 24-well plates for 2 h, washed and incubated under plate-assay conditions. The virucidal effect was defined by the ability of the compound to inactivate the particles not allowing infection and without generating cytopathic effect as observed in the virus-only control.
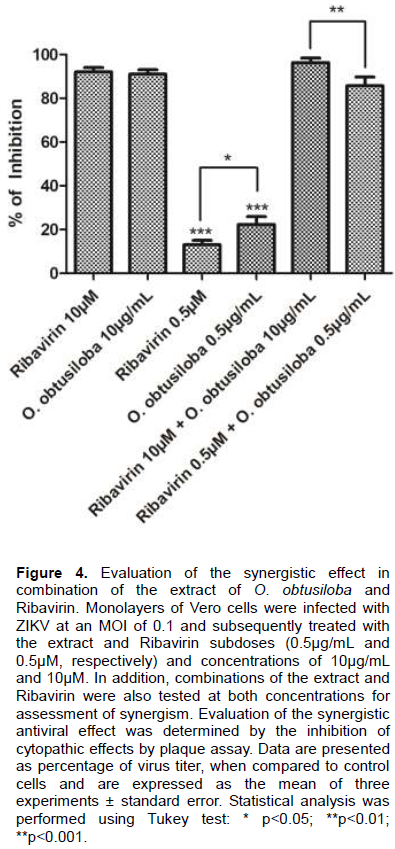
Synergistic effect test O. obtusiloba plus Ribavirin
For this analysis, Vero cells were cultured in 24-well plates subsequently infected with ZIKV MOI 0.1. Subsequently, the infected cells were treated with a high dose of 10 μg / ml extract and 10 μM Ribavirin capable of inhibiting 90% of viral replication. The inhibition with a minimum dose of both 0.5 μg / mL extract and 0.5 μM Ribavirin which are concentrations that inhibit replication below 20% was also evaluated. For the synergism evaluation, we combined the concentrations of the compounds and added the infected cells. After 72 h, inhibition of cytopathic effect was observed by inhibition of viral plaque formation.
Statistical analysis
The data were analyzed by one-way analysis of variance (ANOVA) followed by a Tukey test using the GraphPad Instat version 3 program. A p value of <0.05 was considered statistically significant. The values of p<0.05, p<0.01 and p<0.001 are shown in the figures.
RESULTS
Cytotoxicity and effect of the extract on the ZIKV replication in Vero cells
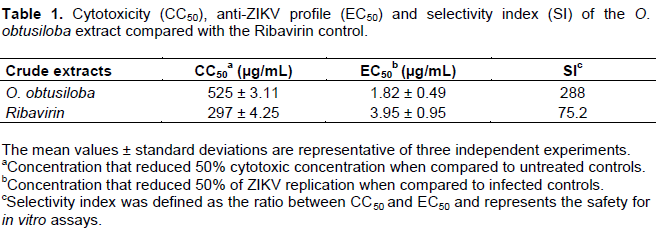
The cytotoxicity (CC50) of the extract from the algae was assessed by MTT (Sigma-Aldrich), as previously described (Mosmann, 1983)with some modifications. The results in Table 1 show that the O. obtusiloba extract produced the best CC50 with a value of 525 µg/mL. Subsequently, the antiviral activities of the extract were evaluated. For these analyzes, different concentrations of the extract was tested starting with a concentration having an inhibitory potential of 20 μg/mL and reducing the concentration progressively. The extract inhibited the replication of ZIKV in a dose-dependent manner (Figure 1). The results demonstrate that O. obtusiloba extract inhibited above 90% of ZIKV replication in the highest concentrations (20 μg/mL) with low EC50 values of 1.82 μg/mL. Based on these data, the Selectivity Index (SI), representing the degree of reliability of the extracts for possible future use, was derived from the relationship between the CC50 and EC50 levels. The values of EC50 and SI presented by O. obtusiloba extract, 1.82 µg/mL and 288 respectively, were better and gave significances and times significantly better than those obtained by ribavirin (EC50= 3.95 µg/mL; SI=75.2) used as a control (Table 1).
Virucidal effect
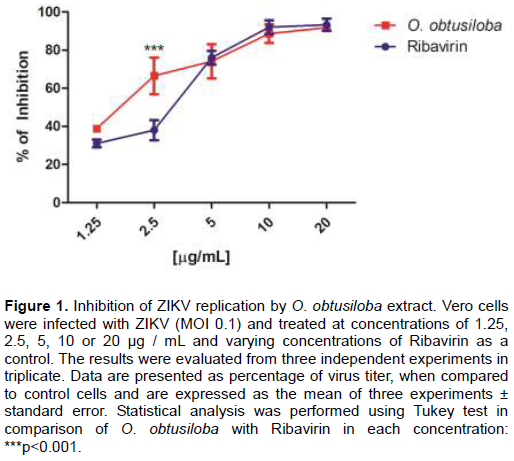
The virucidal activity of O. obtusiloba extract was evaluated against ZIKV. The viral suspension was maintained with different concentrations of the extracts (2.5, 5 and 10 µg/mL) for 2 h and then added to Vero cell cultures. The results showed that O. obtusiloba had a good inactivation capacity of the virus (virucidal effect). Figure 2 shows that the O. obtusiloba extract significantly inhibited ZIKV infectivity at higher levels than Ribavirin at all concentrations tested and at 10 μg/mL of this extract inhibited about 80% of ZIKV replication.
Time of drug addition
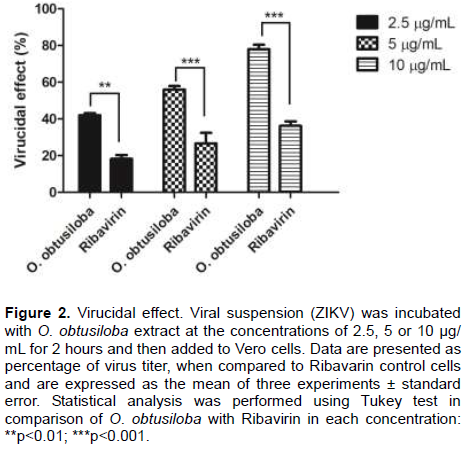
To identify the step at which viral replication might be inhibited, time of addition experiments were performed with the compounds administered 5.0 µg/mL at 3, 2, 1 h before infection. Subsequently, the virus was added at time 0, 1, 2, and 3 h after infection and Ribavirin was used as a control (5 µM). At time 0, O. obtusiloba extract inhibited over 80% of viral replication. O. obtusiloba and Ribavirin maintained an inhibitory effect at the other times but with a decline recorded but even at 3 h after virus infection at least 60% inhibition occurred in viral replication (Figure 3).
Synergism between the extract of Osmundaria obtusiloba and Ribavirin
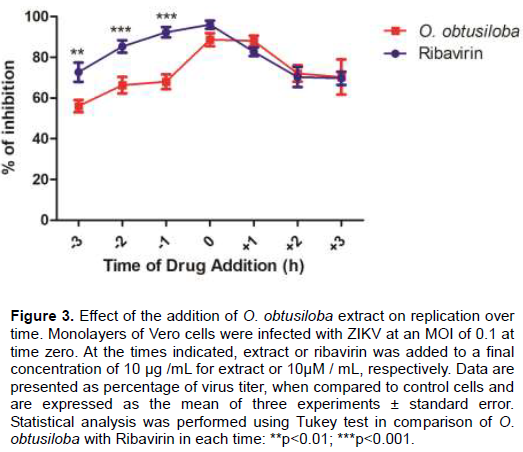
The results obtained clearly show the inhibitory efficiency of the O. obtusiloba extract on ZIKA was greatly increased with the addition of Ribavirin and can be attributed to a synergistic effect. As shown in Figure 4, the addition of subdoses of O. obtusiloba extract (0.5 μg / mL) resulted in only ca. 20% virus inhibition, and, similarly, 0.5 μM doses of Ribavirin recorded ca. 15% inhibition. However, when the seaweed extract and Ribavirin were given together and associated with the lowest concentrations, the inhibitory effect was potentiated and was inhibiting almost 3 times more than the effect of both added at 0.5 μM alone, generating an inhibition of the replication of the ZIKV of approximately 90%, and thus showing characterizing a strong synergistic effect.
DISCUSSION
In this study, the results demonstrate that O. obtusiloba extract inhibited viral replication significantly when cells were treated with various concentrations of the extract and that inhibition was dose-dependent generating an EC50 of 1.82 μg/mL. In addition, there was a low cytotoxicity of the extract on Vero cells resulting in a CC50 of 525 μg/mL. Interestingly, the extract showed a selectivity index (SI) of 288 which, as demonstrated in the literature, is described as good for SI compounds with values ​​greater than 100 (Silva et al., 2011; Zandi et al., 2011).
The literature records a considerable number of studies of antivirals for Dengue that has very significant results (Zandi et al., 2012). However, few studies have been undertaken for other arboviruses that have results as significant as those on Dengue. ZIKA was focused on, which has been associated with severe syndromes (Alvarado-Socarras et al., 2018; Barbi et al., 2018). Previously seaweed extracts have been reported with little cytotoxicity (Alencar et al., 2014)but with considerable antiviral activity, for example, in studies with HIV (Nogueira et al., 2016)and against Herpes (de Souza Barros et al., 2017).
In the present work, initially the seaweed extract was evaluated for virucidals activity and shown to inactivate the ZIKA particles. The studies demonstrated that the extract of O. obtusiloba inactivated the viral particles up to 80% in concentrations of up to 10 μg/mL. Thus, additional studies of this virucidal compound are necessary to develop new strategies for the preparation of preventive measures.
Looking for specific characteristics of the mechanism of action of the extract, studies such as the time of addition of the extracts (Time of Addition Experiment) showed that the O. obsutiloba used at different times both pre- and post-infection has great potential to inhibit the replication of ZIKV, around 60% after treating up to 3 h prior to infection. At time 0, however, the addition of the extract was concomitant with the infection of the cells, and resulted in 90% inhibition of the virus replication. Even if the extract was added up to 3 h after infection, there was still inhibition of around 80% of the virus. As far as infections are concerned, the signs and symptoms can initially be confusing and the follow-up may be delayed but a drug capable of treating late infections would be an important additional strategy. These data suggest the potential of O. obtusiloba as an algal extract candidate for future development. Similarly, Zmurko et al. (2016) performed time of drug addition studies against ZIKV with the 7 DMA viral polymerase inhibitor, but without pretreatment and with posttreatment at different times up to 24 h. They showed that the addition of the compound to the infected cells could be delayed up to ~10 h after infection without much loss of antiviral potency (Zmurko et al., 2016).
There is much interest in searching for combinations of drugs for the inhibition of virus replication as described for Dengue (Yeo et al., 2015), HSV-1 (Mancini et al., 2009), and also for Chikungunya (Mishra et al., 2016). These analyzes are performed in order to reduce the concentration of substances used and to optimize the treatments making them more effective and less toxic. The results herein demonstrated an important synergistic effect by the combination of Ribavirin and the O. obtusiloba extract (Figure 4), since the use of both combined in subdoses were able to inhibit viral replication three times more than both evaluated separately.
CONCLUSIONS
The findings showed that the crude O. obtusiloba seaweed extract tested had activity against ZIKV, demonstrating that marine algae are an interesting source for drug discovery and the development of novel anti-ZIKV agents. Extracts of O. obtusiloba all gave very promising results, and are candidates for further studies to isolate their active factors and better elucidate their mechanisms of action. In summary, O. obtusiloba extract has anti-ZIKV with a particularly significant virucidal effects and synergistic effect in combination with Ribavirin. Currently, there are no vaccines or specific drugs for prevention and treatment of ZIKV infection. The results demonstrate the importance of the marine environment in the search for antivirals drugs with activity against ZIKV.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
The authors are grateful to Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq) for the financial support and for Productivity Fellowships to ICNPP and VLT (443930/2014-7 and 304070/2014-9). ICNPP and VLT (E-26/201.442/2014) also thank Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ) for the Cientista do Nosso Estado Fellowship. CCCS thanks Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) for the Postdoc fellowship and CSB thanks FAPERJ for the Postdoc fellowship (E-26/201.344/2016) in the postgraduate program in sciences and biotechnology of UFF (PPBI-UFF).
REFERENCES
|
Alencar DB, Silva SR, Pires-Cavalcante KM, Lima RL, Pereira Junior FN, Sousa MB, Viana FA, Nagano CS, Nascimento KS, Cavada BS, Sampaio AH, Saker-Sampaio S (2014). Antioxidant potential and cytotoxic activity of two red seaweed species, amansia multifida and meristiella echinocarpa, from the coast of northeastern brazil. Anais da Academia Brasileira de Ciencias 86(1):251-263. |
|
|
Alvarado-Socarras JL, Idrovo AJ, Contreras-Garcia GA, Rodriguez-Morales AJ, Audcent TA, Mogollon-Mendoza AC, Paniz-Mondolfi A (2018). Congenital microcephaly: A diagnostic challenge during zika epidemics. Travel medicine and infectious disease. |
|
|
Barbi L, Coelho AVC, Alencar LCA, Crovella S (2018). Prevalence of guillain-barre syndrome among zika virus infected cases: A systematic review and meta-analysis. The Brazilian Journal of Infectious Diseases 22(2):137-141. |
|
|
Barros CdS, Cirne-Santos CC, Garrido V, Barcelos I, Stephens PRS, Giongo V, Teixeira VL, de Palmer Paixão ICN (2016). Anti-hiv-1 activity of compounds derived from marine alga canistrocarpus cervicornis. Journal of Applied Phycology 28(4):2523-2527. |
|
|
Barros CS, Teixeira VL, Paixão IC (2015). Seaweeds with anti-herpes simplex virus type 1 activity. Journal of Applied Phycology 27(4):1623-1637. |
|
|
Barrows NJ, Campos RK, Powell ST, Prasanth KR, Schott-Lerner G, Soto-Acosta R, Galarza-Mu-oz G, McGrath EL, Urrabaz-Garza R, Gao J (2016). A screen of fda-approved drugs for inhibitors of zika virus infection. Cell Host and Microbe 20(2):259-270. |
|
|
Calvez E, Guillaumot L, Millet L, Marie J, Bossin H, Rama V, Faamoe A, Kilama S, Teurlai M, Mathieu-Daudé F (2016). Genetic diversity and phylogeny of aedes aegypti, the main arbovirus vector in the pacific. PLoS Neglected Tropical Diseases 10(1):e0004374. |
|
|
Cao-Lormeau V-M, Blake A, Mons S, Lastère S, Roche C, Vanhomwegen J, Dub T, Baudouin L, Teissier A, Larre P (2016). Guillain-barré syndrome outbreak associated with zika virus infection in french polynesia: A case-control study. The Lancet 387(10027):1531-1539. |
|
|
Carvalho LRD, Guimarães SM, Roque NF (2006). Sulfated bromophenols from osmundaria obtusiloba (c. Agardh) re norris (rhodophyta, ceramiales). Brazilian Journal of Botany 29(3):453-459. |
|
|
Cirne-Santos CC, Souza TML, Teixeira VL, Fontes CFL, Rebello MA, Castello-Branco LRR, Abreu CM, Tanuri A, Frugulhetti IC, Bou-Habib DC (2008). The dolabellane diterpene dolabelladienetriol is a typical noncompetitive inhibitor of hiv-1 reverse transcriptase enzyme. Antiviral Research 77(1):64-71. |
|
|
de Souza Barros C, Garrido V, Melchiades V, Gomes R, Gomes MWL, Teixeira VL, de Palmer Paixão ICN (2017). Therapeutic efficacy in BALB/C mice of extract from marine alga canistrocarpus cervicornis (phaeophyceae) against herpes simplex virus type 1. Journal of Applied Phycology 29(2):769-773. |
|
|
de Souza Barros C, Gomes MWL, Gomes RdSP, Melchiades V, Nogueira CCR, Cirne-Santos CC, esar, Garrido V, Pinto CEC, Teixeira VL, de Palmer Paixao ICN (2018). Acute toxicity evaluation of ethanol extract of red algae, osmundaria obtusiloba, in balb/c mice. Journal of Medicinal Plants Research 12(17):217-221. |
|
|
de Souza LM, Sassaki GL, Romanos MTV, Barreto-Bergter E (2012). Structural characterization and anti-hsv-1 and hsv-2 activity of glycolipids from the marine algae osmundaria obtusiloba isolated from southeastern brazilian coast. Marine Drugs 10(4):918-931. |
|
|
Deng YQ, Zhang NN, Li CF, Tian M, Hao JN, Xie XP, Shi PY, Qin CF (2016). Adenosine analog nitd008 is a potent inhibitor of zika virus. In Open forum infectious diseases. Oxford University Press 3(4). |
|
|
Faye O, Freire CC, Iamarino A, Faye O, de Oliveira JVC, Diallo M, Zanotto PM (2014). Molecular evolution of zika virus during its emergence in the 20 th century. PLoS Neglected Tropical Diseases 8(1):e2636. |
|
|
Gubler DJ (2001). Human arbovirus infections worldwide. Annals of the New York Academy of Sciences 951: 13-24. |
|
|
Gyawali N, Bradbury RS, Taylor-Robinson AW (2016). Do neglected australian arboviruses pose a global epidemic threat? Australian and New Zealand Journal of Public Health 40(6):596-596. |
|
|
Hamel R, Liégeois F, Wichit S, Pompon J, Diop F, Talignani L, Thomas F, Desprès P, Yssel H, Missé D (2016). Zika virus: Epidemiology, clinical features and host-virus interactions. Microbes and Infection 18(7-8):441-449. |
|
|
Hayashi L, de Paula EJ, Chow F (2007). Growth rate and carrageenan analyses in four strains of kappaphycus alvarezii (rhodophyta, gigartinales) farmed in the subtropical waters of são paulo state, brazil. Journal of Applied Phycology 19(5): 393-399. |
|
|
Heukelbach J, Alencar CH, Kelvin AA, de Oliveira WK, de Góes Cavalcanti LP (2016). Zika virus outbreak in brazil. The Journal of Infection in Developing Countries 10(02):116-120. |
|
|
Iranpour M, Moghadam AR, Yazdi M, Ande SR, Alizadeh J, Wiechec E, Lindsay R, Drebot M, Coombs KM, Ghavami S (2016). Apoptosis, autophagy and unfolded protein response pathways in arbovirus replication and pathogenesis. Expert Reviews in Molecular Medicine 18 p. |
|
|
Junior VLP, Luz K, Parreira R, Ferrinho P (2015). Zika virus: A review to clinicians. Acta Medica Portuguesa 28(6):760-765. |
|
|
Kraemer MU, Sinka ME, Duda KA, Mylne AQ, Shearer FM, Barker CM, Moore CG, Carvalho RG, Coelho GE, Van Bortel W (2015). The global distribution of the arbovirus vectors aedes aegypti and Ae. Albopictus. Elife 4:e08347. |
|
|
Kucharski AJ, Funk S, Eggo RM, Mallet H-P, Edmunds WJ, Nilles EJ (2016). Transmission dynamics of zika virus in island populations: A modelling analysis of the 2013–14 french polynesia outbreak. PLoS Neglected Tropical Diseases 10(5):e0004726. |
|
|
Lindenbach B, Murray C, Thiel H, Rice C (2013). Flaviviridae. Fields Virology 1:712-746. |
|
|
Lira M-LF, Lopes R, Gomes AP, Barcellos G, Verícimo M, Osako K, Ortiz-Ramirez FA, Ramos CJB, Cavalcanti DN, Teixeira VL (2016). Anti-leishmanial activity of brazilian green, brown, and red algae. Journal of Applied Phycology 28(1):591-598. |
|
|
Macedo NRPV, Ribeiro MS, Villaça RC, Ferreira W, Pinto AM, Teixeira VL, Cirne-Santos C, Paixão IC, Giongo V (2012). Caulerpin as a potential antiviral drug against herpes simplex virus type 1. Revista Brasileira de Farmacognosia 22(4):861-867. |
|
|
Mancini DAP, Torres RP, Pinto JR, Mancini-Filho J (2009). Inhibition of DNA virus: Herpes-1 (hsv-1) in cellular culture replication, through an antioxidant treatment extracted from rosemary spice. Brazilian Journal of Pharmaceutical Sciences 45(1):127-133. |
|
|
Mendes GdS, Bravin IC, Yoneshigue-Valentin Y, Yokoya NS, Romanos MTV (2012). Anti-hsv activity of hypnea musciformis cultured with different phytohormones. Revista Brasileira de Farmacognosia 22(4):789-794. |
|
|
Mishra P, Kumar A, Mamidi P, Kumar S, Basantray I, Saswat T, Das I, Nayak TK, Chattopadhyay S, Subudhi BB (2016). Inhibition of chikungunya virus replication by 1-[(2-methylbenzimidazol-1-yl) methyl]-2-oxo-indolin-3-ylidene] amino] thiourea (mbzm-n-ibt). Scientific Reports 6:20122. |
|
|
Mlakar J, Korva M, Tul N, Popović M, Poljšak-Prijatelj M, Mraz J, Kolenc M, Resman Rus K, Vesnaver Vipotnik T, Fabjan Vodušek V (2016). Zika virus associated with microcephaly. New England Journal of Medicine 374(10):951-958. |
|
|
Mosmann T (1983). Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. Journal of Immunological Methods 65(1-2):55-63. |
|
|
Musso D, Nilles E, Cao-Lormeau V-M (2014). Rapid spread of emerging zika virus in the pacific area. Clinical Microbiology and Infection 20(10):O595-O596. |
|
|
Nielsen KK, Bygbjerg IC (2016). Zika virus and hyperglycaemia in pregnancy. The Lancet 387(10030):1812. |
|
|
Nogueira CCR, de Palmer Paixão ICN, Cirne-Santos CC, Stephens PRS, Villaça RC, de Souza Pereira H, Teixeira V, Laneuville E (2016). Anti-hiv-1 activity in human primary cells and anti-hiv-1 rt inhibitory activity of extracts from the red seaweed acanthophora spicifera. Journal of Medicinal Plants Research 10(35):621-625. |
|
|
Rodrigues LC (2016). Microcephaly and zika virus infection. The Lancet 387(10033):2070-2072. |
|
|
Silva A, Morais S, Marques M, Lima D, Santos S, Almeida R, Vieira I, Guedes M (2011). Antiviral activities of extracts and phenolic components of two spondias species against dengue virus. Journal of Venomous Animals and Toxins Including Tropical Diseases 17(4):406-413. |
|
|
Soares AR, Robaina M, Mendes GS, Silva TS, Gestinari L, Pamplona OS, Yoneshigue-Valentin Y, Kaiser CR, Romanos MTV (2012). Antiviral activity of extracts from brazilian seaweeds against herpes simplex virus. Revista Brasileira de Farmacognosia 22(4):714-723. |
|
|
Tabachnick WJ (2016). Climate change and the arboviruses: Lessons from the evolution of the dengue and yellow fever viruses. Annual Review of Virology 3:125-145. |
|
|
Talarico LB, Duarte ME, Zibetti RG, Noseda MD, Damonte EB (2007). An algal-derived dl-galactan hybrid is an efficient preventing agent for in vitro dengue virus infection. Planta Medica 73(14):1464-1468. |
|
|
Tappe D, Nachtigall S, Kapaun A, Schnitzler P, Gunther S, Schmidt-Chanasit J, (2015). Acute zika virus infection after travel to malaysian borneo, september 2014. Emerging infectious diseases 21(5):911-913. |
|
|
Teixeira MG, da Conceição N. Costa M, de Oliveira WK, Nunes ML, Rodrigues LC (2016). The epidemic of zika virus–related microcephaly in brazil: Detection, control, etiology, and future scenarios. American Journal of Public Health 106(4):601-605. |
|
|
Vorou R (2016). Zika virus, vectors, reservoirs, amplifying hosts, and their potential to spread worldwide: What we know and what we should investigate urgently. International Journal of Infectious Diseases 48:85-90. |
|
|
Yeo KL, Chen Y-L, Xu HY, Dong H, Wang Q-Y, Yokokawa F, Shi P-Y (2015). Synergistic suppression of dengue virus replication using a combination of nucleoside analogs and nucleoside synthesis inhibitors. Antimicrobial Agents and Chemotherapy 59(4):2086-2093. |
|
|
Zandi K, Teoh BT, Sam SS, Wong PF, Mustafa MR, Abubakar S (2012). Novel antiviral activity of baicalein against dengue virus. BMC Complementary and Alternative Medicine 12(1):214. |
|
|
Zandi K, Teoh BT, Sam SS, Wong PF, Mustafa MR, Abubakar S (2011). Antiviral activity of four types of bioflavonoid against dengue virus type-2. Virology Journal 8(1):560. |
|
|
Zmurko J, Marques RE, Schols D, Verbeken E, Kaptein SJ, Neyts J (2016). The viral polymerase inhibitor 7-deaza-2'-c-methyladenosine is a potent inhibitor of in vitro zika virus replication and delays disease progression in a robust mouse infection model. PLoS Neglected Tropical Diseases 10(5):e0004695. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


