Full Length Research Paper

ABSTRACT
To broaden the genetic base of the allotetraploid peanut (Arachis hypogaea L.), pre-breeding programs have produced interspecific synthetic allotetraploids resulting from the chromosome duplication of hybrids between peanut related diploid species. These allotetraploids were highly cross-fertile with peanut making it possible to access the extensive genetic variability harbored by the wild species. This study aims to evaluate the impact of polyploidization and hybridization in resveratrol content in Arachis hybrids. Resveratrol is a potent antioxidant that has been shown to be useful in the treatment of many human diseases. For that, resveratrol was characterized in five synthetic allotetraploids of wild Arachis, six diploid wild species, three cultivars of A. hypogaea and three backcross (BC) hybrids between synthetic allotetraploids and A. hypogaea. Leaves from these genotypes were ultraviolet (UV) light irradiated for 2 h 30 min and their resveratrol contents were determined by high performance liquid chromatograph (HPLC). Resveratrol was found in all genotypes, but at variable concentrations. Synthetic allotetraploids and peanut did not differ and diploid species had the lowest resveratrol content. The highest concentrations were observed in hybrids between allotetraploids and cultivars of A. hypogaea that were probably the most heterozygous among the genotypes analyzed since their chromosome sets came from different species. This study data suggest a positive effect of polyploidy and hybridization in resveratrol content.
Key words: Peanut, wild relatives, polyploidy, pre-breeding.
INTRODUCTION
The cultivated peanut Arachis hypogaea L. is an allotetraploid (AABB) that originated from a single crossing event between the diploid wild species Arachis duranensis and Arachis ipaënsis (Kochert et al., 1996; Bertioli et al., 2016). These two species belong to section Arachis, which also comprises the cultivated and its most close relatives (Krapovickas and Gregory, 1994). All the other species of section Arachis are diploids with the single exception of Arachis monticola, which is also a tetraploid (Smartt et al., 1978; Fernández and Krapovickas, 1994; Peñaloza and Valls, 2005; Stalker, 2017).
Section Arachis species constitute the secondary gene pool of the cultivated peanut (Stalker and Moss, 1987) and because of that, many accessions of those species have been characterized and evaluated for several agronomic traits, including resistance to biotic (Stalker, 1984; Pande and Rao, 2001; Michelotto et al., 2015) and abiotic stresses (Nautiyal et al., 2008; Leal-Bertioli et al., 2012).
The reproductive barrier between cultivated and wild Arachis due to ploidy level difference has been overcomed using interspecific synthetic allotetraploids. Sterile diploid hybrids obtained by crossing A and B genome Arachis species have been turned fertile after their tetraplodization with colchicine, which allowed their crossing with peanut and the introgression of alleles from the wild species into the cultivated (Simpson , 1991).
Peanuts are among the few plant species that produce resveratrol (Lanz et al., 1990; Sobolev and Cole, 1999; Arora and Japlan, 2018). This phenolic compound is a potent antioxidant (Frankel et al., 1993) whose healing and preventive potential for many human diseases were described in some recent reviews (Colica et al., 2018; Galiniak et al., 2019). Resveratrol is also a phytoalexin that has been associated with resistance to major peanut diseases (Sobolev et al., 2007). Moreover, ten species of section Arachis also synthesizes resveratrol and three of them had levels higher than those found in cultivar Caiapó of A. hypogaea (Lopes et al., 2013).
The effect of polyploidization and hybrization on different traits in Arachis interspecific synthetic allotetraploids have been studied (Burow et al., 2001; Fávero et al., 2009, 2015; Leal-Bertioli et al., 2012, 2017; Michelotto et al., 2015, 2016, 2017). The characterization of resveratrol content in Arachis allotetraploids could add new value to these genotypes, which have been developed to be used in peanut pre-breeding programs (Bertioli et al, 2011). In this context, the objective of the present study was to evaluate the impact of polyploidization and hybridization in the resveratrol content analyzing synthetic allotetraploids, their respective diploid wild parentals and hybrids between two synthetic allotetraploids and three A. hypogaea cultivars.
METHODOLOGY
Plant material
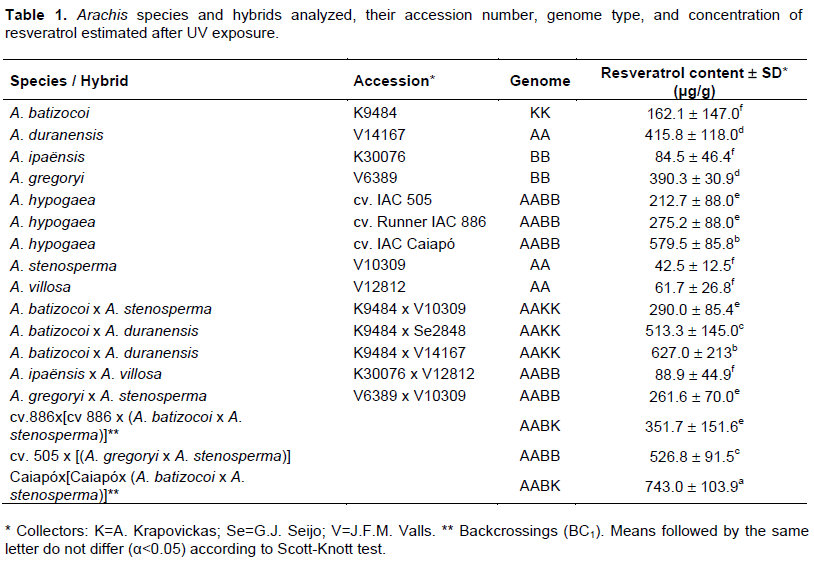
Seventeen Arachis genotypes were analyzed for resveratrol content being six wild diploid species, three tetraploid peanut (A. hypogaea) cultivars and eight tetraploid hybrids that comprised five interspecific synthetic allotetraploids, and three hybrids resulting from crosses between peanut and synthetic allotetraploids (Table 1). The Arachis wild species analyzed harbor different types of genome: A (villosa, stenosperma, A. duranensis), B (A. ipaënsis and Arachis gregoryi) and K (batizocoi). The cultivars of peanut analyzed were ‘IAC Caiapó’, ‘Runner IAC 886’ and ‘IAC 505’. All synthetic allotetraploids and hybrids analyzed were developed by Santos (2013). The plants were grown in greenhouses at Embrapa Genetic Resources and Biotechnology, Brasília, Brazil.
Induction of resveratrol synthesis using UV
The experiments were performed using detached leaves collected from six-month-old Arachis plants in greenhouse conditions, as previously described (Lopes et al., 2013). In short, detached leaves were exposed to an ultraviolet light for 2 h 30 min and maintained in the dark for additional 15 h at room temperature. UV-treated and non-treated control leaves of each genotype were divided into three aliquots of 1 g and stored at -80°C.
Resveratrol extraction and sample preparation
The resveratrol extraction protocol was based on Potrebko and Resurreccion (2009). Prior to high performance liquid chromatograph (HPLC) injection, the dried residue was reconstituted in 6.8 ml of 15% (v/v) ethanol. The samples were vortexed for 1 min and left in ultrasonic bath for four minutes. The procedure was repeated twice to ensure the complete recovery of the extract. The samples were then transferred to 2.0 ml tubes and centrifuged for 15 min at 25°C at 13,400 rpm. The supernatant of the centrifuged material was conditioned in a 2-ml tube and then used for injection in a HPLC (CLAE, Varian®) with ternary pump, automatic dial and coupled photodiode array detector (PDA Varian® PS- 240 / PS-410 / PS-335 / Galaxie Software 1.9).
HPLC analysis
The column used in HPLC was Zorbax XDB Agilent (250 x 4.6 mm, 5 μm), without guard column. A gradient of acetonitrile and a 0.02% aqueous phosphoric acid (J. T. Baker) were used as mobile phase. The conditions were: acetonitrile for 0 min at 13%; 6 to 9 min at 15%; 17 min at 17%; 28 to 33 min at 28%; 40 min at 50%; 45 min at 60%; 46 to 48 min at 80%; 49 to 54 min at 13%; flow rate of 1.0 ml/min. The UV absorption was monitored at 308 nm, 280 nm and also at the maximum absorption wavelength of each eluent (PDA). The injection volume of each sample was 10 μl.
The peak of resveratrol was identified by comparison with the retention time of the commercial standard solutions of resveratrol (> 99%, 230-240 μg/ml, Sigma-Aldrich) and phenolphthalein (> 98%, 2927-2835 μg/ml, Sigma-Aldrich) that were injected daily for area verification. Additional procedures for resveratrol identification were analysis of the spectrum provided by the diode array detector, and the quantification by co-elution with the resveratrol pattern and further comparison of the chromatograms of the induced and control samples. The final concentration of resveratrol per gram of leaf was calculated according to Potrebko and Resurreccion (2009).
Data analysis
The means of the resveratrol production were compared using the Scott and Knott test at 5% probability, considering the groups of plants over time (3 blocks) as covariate, aiming at filtering the variability observed due to these repetitions. The analysis was developed in the statistical language program R, free for download at the site http://www.r-project.org/.
RESULTS AND DISCUSSION
All genotypes analyzed were able to produce resveratrol in response to UV induction (Table 1). Traces of resveratrol (below 0.1 μg) were detected in the samples not exposed to UV (data not shown).
The resveratrol content varied greatly among the six wild diploid species analyzed going from 42.53±12.5 μg/g in A. stenosperma to 415.8±118.0 μg/g in A. duranensis (Table 1). Lopes et al. (2013) detected 370.0 μg/g of resveratrol in UV-treated plants of A. gregoryi (accession V6389) which was very similar to the value found in the present study (390.3±30.9 μg/g). Conversely, for A. batizocoi (accession K9484) and A. ipaënsis (accession K30076), Lopes et al. (2013) detected higher contents (524.5 and 314.0 μg/g, respectively) than those found here (162.1±147.0 μg/g and 84.5±46.4 μg/g, respectively). Also, Carvalho et al. (2017) detected resveratrol in UV-treated leaves of A. duranensis (accession V14167) in concentration (371.97 μg/g) similar to that observed here (415.8±118.0 μg/g), whereas in A. stenosperma, (accession V10309) resveratrol concentration was at least 13-times higher (512.6 μg/g) than in our study (42.49±12.5 μg/g). The differences in the resveratrol content of a same accession observed in these studies may be due to different factors, such as the intrinsic nature of resveratrol as a secondary metabolite, whose production is prone to changes according to the environment temperature (Wang and Zheng, 2001), plant age (Chung et al., 2001), water availability in the soil (Esteban et al., 2001), and cultivation season (Chen et al., 2002). Genetic, ontogenic, morphogenetic, and environmental factors that could cause variation on plant secondary metabolite content in different species were reviewed by Yang et al. (2018).
Concerning the three peanut cultivars evaluated, ‘IAC Caiapó’ presented the highest resveratrol concentration (579.5 ± 85.8b μg/g), followed by ‘Runner IAC 886’ (275.2 ± 88.0e μg/g) and ‘IAC 505’ (212.7 ± 88.0e μg/g) that showed similar concentrations to each other. Over the years, many studies have shown a variable resveratrol content among A. hypogaea varieties/cultivars, with differences due to the plant organ studied, crop location, annual season and pathogens infestation levels. Sanders et al. (2000) found differences among resveratrol content (from 0.03 to 0.147 µg/g) in seeds without coat of three peanut market types (Virginia, Runner, and Spanish) produced in different areas and without any specific induction of resveratrol, as UV used in this study. Significant variations in resveratrol content (from 0.125 to 1.626 μg/g) was also found when seeds of 20 germplasm accessions of A. hypogaea harvested from the same field were analyzed using HPLC (Wang and Pittman, 2009). Variation on resveratrol content was also found in roots of three peanut cultivars grown in 2000 fall and 2001 spring being the content of fall crops much higher than those of spring (Chen et al., 2002). Peanut cultivar ‘IAC Caiapó’ higher resveratrol content (ranging from 300 to 600 μg/kg) when compared to the cultivar ‘IAC 886’ (Zorzete et al., 2011). This last cultivar is less susceptible to thrips Enneothrips flavens infection (Moraes et al., 2005). Considering that resveratrol is a phytoalexin, peanut cultivars with higher concentrations of this metabolite are likely more resistant against pathogen attack. Resveratrol content in peanut seeds was negatively correlated with aflatoxin production and in vitro trials demonstrated that resveratrol could inhibit aflatoxin production (HouMiao et al., 2012). An association between total phytoalexin production and genotype resistance to major peanut diseases was observed being trans-resveratrol was one of the main compounds found in stress-resistant genotypes (Sobolev et al., 2007).
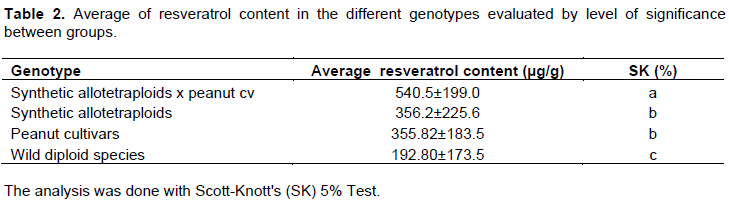
The grouping of the genotypes according to their ploidy level (Table 2), helped to observe that the tetraploids genotypes (peanut cultivars and hybrids) showed significantly higher resveratrol contents than the wild diploid species. The effect of polyploidization has been studied in Arachis comparing synthetic allotetraploids, their corresponding diploid parental species and peanut cultivars. The comparison among A. duranensis (V14167), A. ipaënsis (KG30076), a synthetic allotetraploid (A. duranensis V14167 × A. ipaënsis K 30076)4× and A. hypogaea subsp. hypogaea var. hypogaea ‘Runner IAC 886’ showed some diploid traits such as chlorophyll meter readings are maintained through hybridization and polyploidization and most characters are substantially modified (Leal-Bertioli et al., 2012). An increase in resistance to the foliar diseases rust (Puccinia arachidis) and late leaf spot (Cercosporidium personatum) was observed in the synthetic allotetraploids compared to their diploid parental species (Kumari et al., 2014). More recently, it was also demonstrated that Arachis allotetraploids have some general phenotypic trends that are common, regardless of the combination of their wild parental diploid suggesting that nucleotypic effect is more important than new allelic combination (Leal-Bertioli et al., 2017). The effect of polyploidization on the increase of bioactive compounds has also been studied in some other species. The concentrations of some phytoconstituents, such as emodin, physcion, piceatannol, resveratrol and rutin were determined by LC–MS in three species of Rumex and a positive correlation could be detected with the increasing ploidy status in different chromosomal races (Jeelania et al., 2017).
Our data suggested that the polyploidization could be one of the causes of the increase in the resveratrol content observed in the polyploidy compared to diploid samples analyzed.
The resveratrol content among the synthetic allotetraploids (Table 1) ranged from 88.9 ± 44.9f μg/g for A. ipaënsis x A. villosa to 627.0 ± 213.2b μg/g for A. batizocoi x A. duranensis V14167. Interestingly, the parental diploids of A. ipaënsis X A. villosa that had the lowest resveratrol content (88.9 ± 44.9f μg/g for) among the synthetic allotetraploids displayed the second and third lowest concentrations of resveratrol among the wild diploids (61.7 ± 26.8f and 84.5 ± 46.4f μg/g for A. villosa and A. ipaënsis, respectively). Likewise, the synthetic allotetraploid with the highest resveratrol content (513.3 ± 145.0c μg/g for A. batizocoi X A. duranensis V14167) had at least one parental diploids with high content (162.1 ± 147.0f and 415.8 ± 118.0d μg/g for A. batizocoi and A. duranensis, respectively). Overall, we observed that the hybrids that produced high quantities of resveratrol resulted from crosses between parents with the highest levels of resveratrol. Increase on ginsenoside content was also obtained using a interspecific Panax F1 hybrids (Kim et al., 2016). The use of hybridization to increase flavonoids using wild relatives in many cultivated species was recently reviewed (D’Amelia et al., 2018). Thus, our results suggest that, besides the polyploidization, the allelic composition of the allotetraploids might also be positively related to the production of resveratrol in Arachis.
The three hybrids resulting from the crosses between peanut cultivars and synthetic allotetraploids presented significant differences compared to the other genotypes analyzed, showing the highest resveratrol content averages (Table 2). Those hybrids were the most heterozygous among genotypes analyzed in this study since each of their four chromosomes sets came from peanut and two of the wild species used in the synthetic allotetraploids synthesis. This suggested that increase in heterozygosity might also have contributed to increase of resveratrol content. Besides, those hybrid chromosomes were the only ones among the material evaluated that had their chromosomes resultin from the recombination between A and B genomes from the cultivated with A and B or K genomes from the wild species. The other genotypes (diploid and synthetic polyploidy) were most probably homozygous since wild species are most autogamous and because of that recombination would not result in any new allelic combination as it happened in BC1 hybrids.
On average, the three hybrids between synthetic allotetraploids and peanut cultivars had the highest resveratrol content. Previous study showed that hybrids of peanut with interspecific synthetic allotetraploids showed an increased concentration of flavonoids than their parental that resulted in an increased larval mortality of Spodoptera litura (Mallikarjuna et al., 2004).
Variation on resveratrol content was found among the three BCs hybrids analyzed (Table 1). The hybrid [‘cv 886’ X [‘cv 886’ X (A. batizocoi X A. stenosperma)] that had ‘cv 886’ (275.2 ± 88.0e μg/g) as a parental showed lower resveratrol content concentration (351.7 ± 151.6e μg/g) than the one that had Caiapó’ (579.5 ± 85.8b μg/g) as parental [‘Caiapó’ X [‘Caiapó’ X (A. batizocoi X A stenosperma)] that had 743.0 ± 103.9a μg/g). This result suggested that peanut cultivar used as the parental in these crosses highly influences the resveratrol content in the resulting hybrids.
CONCLUSION
This study data suggest a positive effect of polyploidy and hybridization in resveratrol content in Arachis hybrids. Resveratrol can be synthesized by a few species and the major dietary natural sources include grapes, wine, peanuts, and soybeans (Burns et al., 2002). Our data opens the possibility to create and provide new sources of natural resveratrol by the used of interspecific synthetic Arachis hybrids analyzed, mainly the BCs genotypes, which displayed higher resveratrol contents than wild and the cultivated species.
ACKNOWLEDGMENTS
The authors thank Embrapa (Empresa Brasileira de Pesquisa Agropecuária) and CNPq (Conselho Nacional de Desenvolvimento Científico e Tecnológico) for the financial support (Embrapa grant number 03.09.01.014.00.00 and CNPq grant number 482562/2011-0) along with CAPES (Coordenação de Aperfeiçoamento de Pessoal de Nível Superior) for Paula Vasconcelos´ scholarship.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Arora D, Jaglan S (2018). Therapeutic applications of resveratrol nanoformulations. Environmental Chemistry Letters 16:35-41. |
|
|
Bertioli D, Seijo G, Freitas F, Valls J, Leal-Bertioli S, Moretzsohn M (2011). An overview of peanut and its wild relatives. Plant Genetic Resources 9:134-149. |
|
|
Bertioli DJ, Cannon SB, Froenicke L, Huang G, Farmer AD, Cannon EKS, Liu X, Gao D, Clevenger J, Dash S, Ren L, Moretzsohn MC, Shirasawa K, Huang W, Vidigal B, Abernathy B, Chu Y, Niederhuth CE, Umale P, Araújo ACG, Kozik A, Kim KD, Burow MD, Varshney RK, Wang X, Zhang X, Barkley N, Guimarães PM, Isobe S, Guo B, Liao B, Stalker HT, Schmitz RJ, Scheffler BE, Leal-Bertioli SCM, Xun X, Jackson SA, Michelmore R, Ozias-Akins P (2016). The genome sequences of Arachis duranensis and Arachis ipaënsis, the diploid ancestors of cultivated peanut. Nature Genetics 48:438-446. |
|
|
Burns J, Yokota T, Ashihara H, Lean ME, Crozier A (2002). Plant foods and herbal sources of resveratrol. Journal of Agricultural and Food Chemistry 50:3337-3340. |
|
|
Burow MD, Simpson CE, Starr JL, Paterson AH (2001). Transmission genetics of chromatin from a synthetic amphidiploid to cultivated peanut (Arachis hypogaea L.): Broadening the gene pool of a monophyletic polyploid species. Genetics 159: 823-837. |
|
|
Carvalho PASV, Brasileiro AC, Leal-Bertioli S, Bertioli DJ, Silva JP, Agostini-Costa TS, Gimenes MA (2017). Coupled transcript and metabolite identification: insights on induction and synthesis of resveratrol in peanut, wild relatives and synthetic allotetraploid. Genetics and Molecular Research 16:3. |
|
|
Chen RS, Wu PL, Chiou RYY (2002). Peanut roots as a source of resveratrol. Journal of Agricultural and Food Chemistry 50:1665-1667. |
|
|
Chung I-M, Park MR, Rehman S, Yun SJ (2001). Tissue specific and inducible expression of resveratrol synthase gene in peanut plants. Molecules and Cells 12:353-359. |
|
|
Colica C, Milanovićb M, Milićb N, Aielloc V, De Lorenzo A, Abenavolie AA (2018). Systematic review on natural antioxidant properties of resveratrol. Natural Product Communications 13:1195-1203. |
|
|
D'Amelia V, Aversano R, Chiaiese P, Carputo D (2018). The antioxidant properties of plant flavonoids: their exploitation by molecular plant breeding. Phytochemistry Reviews 17: 611. |
|
|
Esteban MA, Villanueva MJ, Lissarrague JR (2001). Effect of irrigation on changes in the anthocyanin composition of the skin of cv Tempranillo (Vitis vinifera L) grape berries during ripening. Journal of the Science of Food and Agriculture 81:409-420. |
|
|
Fávero AP, Moraes SAD, Garcia AAF, Valls JFM, Vello NA (2009). Characterization of rust, early and late leaf spot resistance in wild and cultivated peanut germplasm. Scientia Agricola 66: 110-117. |
|
|
Fávero AP, Pádua JG, Costa TS, Gimenes MA, Godoy IJ, Moretzsohn MC, Michelotto MD (2015). New hybrids from peanut (Arachis hypogaea L.) and synthetic amphidiploid crosses show promise in increasing pest and disease tolerance. Genetics and Molecular Research 14:16694-16703. |
|
|
Fernández A, Krapovickas A (1994). Cromosomas y evolucion en Arachis (Leguminosae). Bonplandia 8:187-220. |
|
|
Frankel EN, German JB, Kinsella JE, Parks E, Kanner J (1993). Inhibition of oxidation of human low-density lipoprotein by phenolic substances in red wine. The Lancet 341:454-457. |
|
|
Galiniak S, Aebisher D, Bartusik-Aebishe D (2019). Health benefits of resveratrol administration. Acta Biochimica Polonica 66:13-21. |
|
|
HouMiao W, Huang J, Lei Y, Yan L, Wang S, Jiang H, Ren X, Lou Q, Liao B (2012). Relationship of resveratrol content and resistance to aflatoxin accumulation caused by Aspergillus flavus in peanut seeds. Acta Agronomica Sinica 38:1875-1883. |
|
|
Jeelania SM, Farooqb U, Guptac AP, Lattooa SK (2017). Phytochemical evaluation of major bioactive compounds in different cytotypes of five species of Rumex L. Industrial Crops and Products 109: 897-904. |
|
|
Kochert G, Stalker H, Gimenes M, Galgaro L, Lopes C, Moore K (1996). RFLP and cytogenetic evidence on the origin and evolution of allotetraploid domesticated peanut Arachis hypogaea (Leguminosae). American Journal of Botany 83:1282-1291. |
|
|
Krapovickas A, Gregory WC (1994). Taxonomia del genero Arachis (Leguminosae). Bonplandia 8:1-186. |
|
|
Kim YJ, Silva J, Zhang D, Shi J, Joo SC, Jang M-G , Kwon W-S, Yan D-C (2016). Development of interspecies hybrids to increase ginseng biomass and ginsenoside yield. Plant Cell Report 35:779-790. |
|
|
Kumari V, Gowda MVC, Tasiwal V, Pandey MK, Bhat RS, Mallikarjuna N, Upadhyaya HD, Varshney RK (2014). Diversification of primary gene pool through introgression of resistance to foliar diseases from synthetic amphidiploids to cultivated groundnut (Arachis hypogaea L.). The Crop Journal 2:110-119. |
|
|
Lanz T, Schröder G, Schröder J (1990). Differential regulation of genes for resveratrol synthase in cell cultures of Arachis hypogaea L. Planta 181:169-175. |
|
|
Leal-Bertioli SC, Bertioli DJ, Guimarães PM, Pereira TD, Galhardo I, Silva JP, Brasileiro ACM, Oliveira RS, Silva PIT, Vadez V, Araujo ACG (2012). The effect of tetraploidization of wild Arachis on leaf morphology and other drought-related traits. Environmental and Experimental Botany 84:17-24. |
|
|
Leal-Bertioli SC, Moretzsohn MC, Santos SP, Brasileiro AC, Guimarães PM, Bertioli DJ, Araujo ACG (2017). Phenotypic effects of allotetraploidization of wild Arachis and their implications for peanut domestication. American Journal of Botany 104:379-388. |
|
|
Lopes RM, Silveira D, Gimenes MA, Vasconcelos PAS, Rosa de Belem NA, Silva JP, da Silveira Agostini-Costa T (2013). Characterization of resveratrol content in ten wild species of section Arachis, genus Arachis. Genetic Resources and Crop Evolution 60:2219-2226. |
|
|
Mallikarjuna N, Kranthi KR, Jadhav DR, Kranthi S, Chandra S (2004). Influence of foliar chemical compounds on the development of Spodoptera litura (Fab.) in interspecific derivatives of groundnut. Journal of Applied Entomology 128: 321-328. |
|
|
Michelotto MD, Barioni Jr W, de Resende MDV, de Godoy IJ, Leonardecz E, Fávero AP (2015). Identification of fungus resistant wild accessions and interspecific hybrids of the genus Arachis. PloS One 10:0128811. |
|
|
Michelotto MD, de Godoy IJ, dos Santos JF, Martins ALM, Leonardecz E, Fávero AP (2016). Identifying amphidiploids resistant to foliar fungal diseases. Crop Science 56:1792-1798. |
|
|
Michelotto MD, de Godoy IJ, Pirotta MZ, dos Santos JF, Finoto EL, Fávero AP (2017). Resistance to thrips (Enneothrips flavens) in wild and amphidiploid Arachis species. PloS One 12: 0176811. |
|
|
Moraes ARAD, Lourenção AL, Godoy IJD, Teixeira GDC (2005). Infestation by Enneothrips flavens Moulton and yield of peanut cultivars. Scientia Agricola 62:69-472. |
|
|
Nautiyal PC, Rajgopal K, Zala PV, Pujari DS, Basu M, Dhadhal BA, Nandre BM (2008). Evaluation of wild Arachis species for abiotic stress tolerance: I. Thermal stress and leaf water relations. Euphytica 159:43-57. |
|
|
Pande S, Rao JN (2001). Resistance of wild Arachis species to late leaf spot and rust in greenhouse trials. Plant Disease 85:851-855. |
|
|
Peñaloza APS, Valls JFM (2005). Chromosome number and satellite chromosome morphology of eleven species of Arachis (Leguminosae). Bonplandia 14:65-72. |
|
|
Potrebko I, Resurreccion AVA (2009). Effect of ultraviolet doses in combined ultraviolet−ultrasound treatments on trans-resveratrol and trans-piceid contents in sliced peanut kernels. Journal of Agricultural and Food Chemistry 57:7750-7756. |
|
|
Sanders TH, McMichael RW, Hendrix KW (2000). Occurrence of resveratrol in edible peanuts. Journal of Agricultural and Food Chemistry 48:1243-1246. |
|
|
Santos SPD (2013). Produção e caracterização de alotetraploides sintéticos entre espécies silvestres do gênero Arachis. Dissertação de mestrado Universidade de Brasília 89. |
|
|
Simpson CE (1991). Pathways for introgression of pest resistance into Arachis hypogaea L. Peanut Science 18:22-26. |
|
|
Smartt J, Gregory WC, Gregory MP (1978). The genomes of Arachis hypogaea. Cytogenetic studies of putative genome donors. Euphytica 27:665-675. |
|
|
Sobolev VS, Cole RJ (1999). Trans-Resveratrol content in commercial peanuts and peanut products. Journal of Agricultural and Food Chemistry 47:1435-1439. |
|
|
Sobolev VS, Guo BZ, Holbrook CC, Lynch RE (2007). Interrelationship of phytoalexin production and disease resistance in selected peanut genotypes. Journal of Agricultural and Food Chemistry 55:2195-2200. |
|
|
Stalker HT, Moss JP (1987). Speciation, citogenetics and utilization of Arachis species. Advances in Agronomy 41:1-40. |
|
|
Stalker HT (1984). Utilizing Arachis cardenasii as a source of Cercospora leafspot resistance for peanut improvement. Euphytica 33:529-538. |
|
|
Stalker HT (2017). Utilizing wild species for peanut improvement. Crop Science 57:1102-1120. |
|
|
Wang SY, Zheng W (2001). Effect of plant growth temperature on antioxidant capacity in strawberry. Journal of Agricultural and Food Chemistry 49:4977-4982. |
|
|
Wang ML, Pittman RN (2009). Resveratrol content in seeds of peanut germplasm quantified by HPLC. Plant Genetic Resources 7:80-83. |
|
|
Zorzete P, Reis TA, Felicio JD, Baquião AC, Makimoto P, Corrêa B (2011). Fungi, mycotoxins and phytoalexin in peanut varieties, during plant growth in the field. Food Chemistry 129:957-964. |
|
|
Yang L, Wen K-S, Ruan X, Zhao Y-X, Wei F, Wang Q (2018). Response of Plant Secondary Metabolites to Environmental Factors. Molecules 23:762. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


