Full Length Research Paper

ABSTRACT
Lead and di-(2-ethylhexyl) phthalate (DEHP) are common environmental toxicants of concern around the world. Although effects of individual exposures to both agents are well documented, there is a dearth of information on the effects of co-exposure to both agents. In this study, combined exposure to lead and DEHP was investigated for effects on ATPase activities in the liver, brain and kidney tissues of rats. Male albino rats were daily exposed to either 200 ppm lead as lead acetate in their drinking water and/or 100 mg DEHP kg-1 body weight in olive oil by gastric intubation for 30 days. Changes in total body weight, relative organ weights as well as brain, hepatic and renal activities of total, Na+K+ -, Ca2+ - and Mg2+-ATPases were used as biomarkers of toxicity. Hepatomegaly and brain atrophy heralded exposure to both agents. Individual exposure to lead and DEHP resulted in reduction in hepatic Ca2+- and Mg2+- ATPase activities but no significant effect on total ATPase activity, however combined exposure produced significant activation of Ca2+-, Na+K+- and total ATPase while restoring Mg2+ - ATPase towards control. A potentiating effect on lead by DEHP was observed in hepatic Na+K+ - ATPase. Lead stimulated the activities of renal Ca2+- and total ATPases while DEHP on the contrary caused significant reduction in total ATPase activity and no significant effects on Ca2+- ATPase activity. Co-treatment produced antagonistic effects leading to normal renal Ca2+- and total ATPase activities. Brain Na+K+ -, Ca2+ - and total ATPase activities were depressed in co-exposure while Mg2+ - ATPase was up-regulated. Lead potentiated DEHP-induced inhibition of brain total - ATPase while co-treatment produced antagonistic effects on brain Ca2+ - ATPase. The findings of this study highlight organ specific variations in response to combined lead and DEHP exposure in rats.
Key words: Hepatotoxic, neurotoxic, ATPases, DEHP, lead, co-exposure, hepatomegaly.
INTRODUCTION
Despite the global move to phase out leaded-gasoline, lead has persisted as an environmental pollutant of grave concern with its deleterious effects particularly felt among children (Nriagu et al., 1996). The emission of lead by power plants, smelters and boilers that burn used motor oil is frequently deposited in the soil, where it is absorbed by crops and leaching into underground water. Hence lead ends up in food, water and air (Allouche et al., 2011). In the recent past,lead poisoning of significant proportions broke out in northern Nigeria and claimed the lives of over 400 children (Medecins Sans Frontieres, 2012). Lead is known to cause diseases in several systems of the body such as the hematopoietic and cardiovascular (Onunkwor et al., 2004; Adeniyi et al., 2008), immunological (Basaran and Undeger, 2000), hepatic (Sharma et al., 2011b), renal (Rastogi, 2008) and central nervous system effects (Sanders et al., 2009; Ademuyiwa et al., 2007; Sharma et al., 2011a) and it has also been implicated as a co-carcinogen and a teratogen (Pracheta and Singh, 2009; Markovac and Goldstein, 1988). Several studies have shown that lead is ubiquitous in the environment (Ademuyiwa et al., 2002; Adeniyi and Anetor, 1999) and causes oxidative stress in the body by interacting with glutathione, a known natural antioxidant in the body (Jangid et al., 2016). Glutathione is a tripeptide consisting of γ-glutamic acid, cysteine and glycine. It is found in several tissues (Kosnet et al., 1998) and lead’s interaction with glutathione leads to replacement of the hydrogen on two sulphydryl groups on adjacent molecules by lead. The strong bond that results effectively eliminates the two glutathione molecules from further reaction thereby eliciting all the conditions that arise as a result of oxidative stress, such as diabetes, nervous system disorders, cardiovascular diseases, aging, cancer etc (Ademuyiwa et al, 2005; 2007; Dosumu et al., 2005).
Di-(2-ethylhexyl) phthalate (DEHP) is another equally widespread toxicant in the environment because of its common use as a plasticizer in the plastic making industry. It is added to plastic to make it softer and more flexible. However, DEHP easily dislodges from the complex with the plastic and leaches into the contents of the plastic containers especially at high temperatures (Fromme et al., 2012). This is how storing foods and drinks in plastic wraps and bottles could lead to phthalate toxicity. DEHP could also leach into blood when it is used in medical devices such as intravenous tubing and blood storage bags (Padmakumaran et al., 1998). Despite the potential hazard, there is a steady rise in the use of plastic bottles as against glass in the storage of drinks and beverages. Phthalates have been associated with endocrine disruption (Ye et al., 2017; Grün, 2010; Miodovnik et al., 2011), metabolic disorders, reproductive and developmental defects (Foster et al., 2001; Jiang, 2007; Engel et al., 2010). DEHP is one of the most commonly used of the phthalates and hence the most abundant in the environment (Kamrin, 2009). Since both lead and DEHP are quite ubiquitous in the environment and may be present in air, soil and drinking water at the same time, there is a high potential for a simultaneous exposure to both toxicants. Both agents are known to exert toxicity through the generation of oxidative stress (Onunkwor et al., 2004; Dosumu et al., 2005; Xu et al., 2008; Afolabi et al., 2015; Wójtowicz et al., 2019) and this makes the study of their co-exposure of particular interest. Lead and phthalates have attracted much research attention separately, however, there is scarce information on their combined effect on key biomarker enzymes such as the ATPases.
ATPases are membrane bound enzymes involved in the uphill transport of ions coupled with energy expenditure in the form of ATP. Previous studies on lead toxicity in humans showed that erythrocyte membrane ATPases were down-regulated by lead (Yücebilgic et al., 2003; Abam et al., 2008, Okediran et al., 2009). Specifically, erythrocyte membrane Na+K+-ATPase and Ca2+-ATPase were shown to be inhibited by lead in occupationally exposed workers (Yücebilgic et al., 2003) while phthalates were also shown to inhibit some ATPases in the organs of rats (Afolabi et al., 2016; Dhanya et al., 2003). The focus of this study therefore was to investigate the effects of combined exposure to lead and DEHP on membrane ATPases of brain, liver and kidney of rats.
MATERIALS AND METHODS
Chemicals
Lead acetate and DEHP were products of Sigma-Aldrich, Munich, Germany. All other chemicals used in this study were of the purest grade available and were obtained from British Drug House (BDH) Chemicals Limited, Poole, England.
Experimental design
Twenty-four (24) male Wistar rats were obtained from the animal house of University of Ibadan (UI), Ibadan, Oyo state, weighing an average of 140 g and housed in the animal house of Bells University of Technology, Ota, Ogun State Nigeria. The animals were acclimatized for 14 days prior to the commencement of the study. Afterwards, they were randomly distributed into four groups of six animals each. Group 1 animals served as control and were administered olive oil and lead-free distilled water. Group 2 animals received 200 ppm lead as lead acetate in their drinking water. Group 3 animals were administered a homologous mixture of 100 mg DEHP kg-1 body weight in olive oil by gastric intubation. Group 4 received both 200 ppm lead in drinking water and 100 mg DEHP kg-1 body weight in olive oil. Doses were chosen based on previous
studies (Ademuyiwa et al., 2009; Afolabi et al., 2015, 2016). These treatments were maintained for thirty days. Animals were placed on a standard pellet diet and distilled water (or their various lead-poisoned water) ad libitum for the duration of the study. Weights were monitored weekly using a weighing balance with accuracy ± 0.1g. At the end of the treatment, animals were fasted overnight and blood collected by cardiac puncture under light ether anesthesia. Plasma was obtained from the blood by centrifuging at 3,000 g for 10 minutes. The brain, liver and kidney were removed and homogenized in 150 mmol L-1 ice-cold KCl to obtain a 10 % homogenate which was further centrifuged at 15,000 g for 15 minutes at 4°C to obtain the post-mitochondrial fraction which was stored at -20°C until further biochemical analyses. The experiment was carried out in accordance with the Bells University of Technology, Department of Chemical and Food Sciences (Biochemistry Unit) guidelines for the care and use of laboratory animals.
Biochemical analyses
Total ATPase (E.C.3.6.1.3) activity in tissue homogenates was assayed using the method of Evans (1969). The final assay mixture of 2 ml contained 0.1 M Tris-HCl (pH 7.4), 0.1 M NaCl, 0.1 M MgCl2, 0.01 M ATP, 0.1 M KCl and 0.1 ml of the tissue homogenate. The reaction was incubated at 37°C in a water bath for 20 min and stopped by adding 1 ml of 10% trichloroacetic acid (TCA) and then centrifuged. The inorganic phosphorus (Pi) liberated was then estimated in the supernatant by the method of Lowry and Lopez (1946) which involved the use of ammonium molybdate and vitamin C.
Assay for Na+/K+-ATPase (E.C.3.6.1.37) activity followed the procedure of Hesketh et al. (1978) as described by Afolabi et al (2016). The reaction mixture contained 0.35 M NaCl, 17.5 mM KCl, 21.0 mM MgCl2, 10 mM Tris-HCl (pH 7.4), 8.0 mM ATP and tissue homogenate. The mixture was incubated at 37°C for 1 h and the reaction terminated by the addition of 0.8 ml of ice-cold 10% (w/v) TCA. Afterwards, the mixture was allowed to stand for 20 min at 4°C and then centrifuged. Pi in the supernatant was measured as described already.
The activity of Ca2+-ATPase (E.C.3.6.1.38) was assayed according to the method of Hjerten and Pan (1983). The reaction mixture contained 125 mM Tris-HCl buffer (pH 8), 50 mM CaCl2 and 10 mM ATP. The contents were incubated at 37°C for 15 min and the reaction arrested by the addition of 0.5 ml of ice-cold 10% TCA and centrifuged. Pi was then determined as described already. The activity of Mg2+-ATPase (E.C.3.6.1.39) was assayed according to the method of Ohnishi et al. (1982). The reaction mixture contained 0.1 ml each of 375 mM Tris-HCl buffer (pH 7.6), 25 mM MgCl2, 10 mM ATP and tissue homogenate. The contents were incubated at 37°C for 15 min and the reaction was then stopped with the addition of 0.5 ml of 10% TCA and centrifuged. Pi was then determined following the technique as described already.
Protein concentration was determined in the brain, liver and kidney homogenates using Bradford method (Bradford, 1976).
Statistical analysis
Results are expressed as mean ± SEM. The statistical significance was evaluated by one-way analysis of variance (ANOVA) followed by Tukey Post Hoc Multiple Comparisons using Statistical Package for the Social Sciences (SPSS) version 15.00 for Windows (SPSS Inc; CA, USA). A value of p < 0.05 was considered statistically significant between groups.
RESULTS
Summary of percentage change in body weight and relative organ weight is depicted in Table 1. Rats treated with lead and DEHP did not show any obvious changes in behavior throughout the period of the study. Animals in all the groups increased in weight at the end of the exposure period except the animals treated with DEHP that experienced a 12.54 ± 1.16% reduction in body weight. Treatment with lead-only resulted in a weight gain of 10.99 ± 3.97% and increases in relative organ weights of the liver and kidney of the rats. Such increases were not observed in the brain in all the treatment groups (relative brain weights decreased by 15, 16 and 29% in the lead-only, DEHP and combined treatment groups respectively). DEHP and co-treatment with both toxicants resulted in increases in relative organ weights of the liver (23 and 19% respectively). Combined treatment yielded the highest gain in body weight (amounting to 18.38%) and highest decrease in relative brain weight (29%).
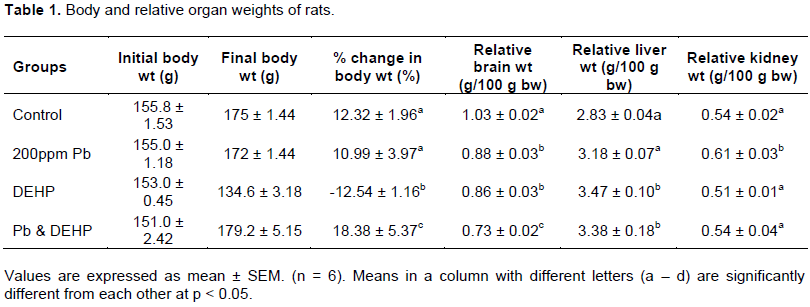
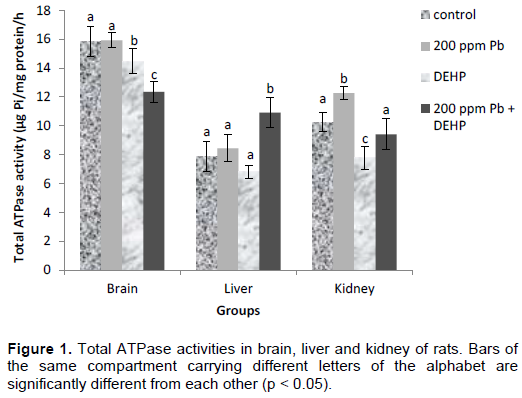
ATPase activities in the organs of control and experimental animals are depicted in Figures 1 to 4. Lead exposure resulted in increase in total ATPase activity of 20% in the kidney of the rats (Figure 1) while there was no significant difference in the liver and brain. Hepatic, renal and brain activities of total ATPases were significantly reduced by DEHP (14, 24 and 9% respectively), however, co-treatment produced a significant activation of total ATPase activity in the liver (39%) while lead showed a potentiating effect on DEHP in the brain. Lead and DEHP produced antagonistic effects in the kidney effectively restoring enzyme activity towards control value. There were significant decreases in Na+K+ ATPase activities between control and treatment groups in the brain (amounting to 11, 12 and 14% in the lead, DEHP and co-treatment groups respectively) as displayed in Figure 2. On the contrary, in the liver, the activity of Na+K+ ATPase was increased by as much as 132 and 32% by lead and DEHP respectively and co-treatment produced additive effects (211%). There was no significant difference in renal Na+K+ ATPase activity between control and lead treated groups, however DEHP and co-treatment caused approximately the same level of inhibition of the enzyme (16%).
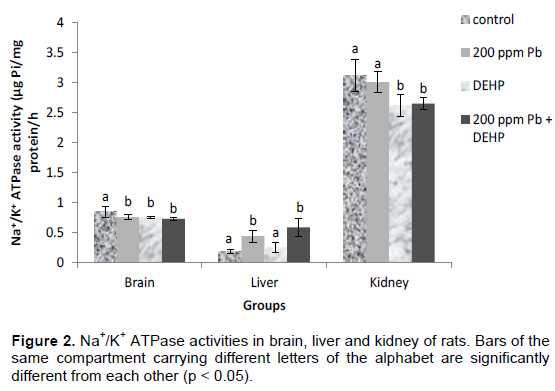
Activities of Ca2+ ATPases in the organs of the rats are presented in Figure 3. After 30 days, lead and DEHP inhibited Ca2+ ATPase to the tune of 6 and 36 % respectively in the brain while lead antagonized the effects of DEHP in the co-treatment group, reducing DEHP’s inhibition from 36 to 21%. In the liver, there was 14% inhibition of Ca2+ ATPase by lead and 24% by DEHP, while combined treatment led to the activation of the enzyme by 17 %. In the kidney, Ca2+ ATPase activity was activated by 25% by lead, while there was no significant difference (p>0.05) in the DEHP and co-treatment groups.
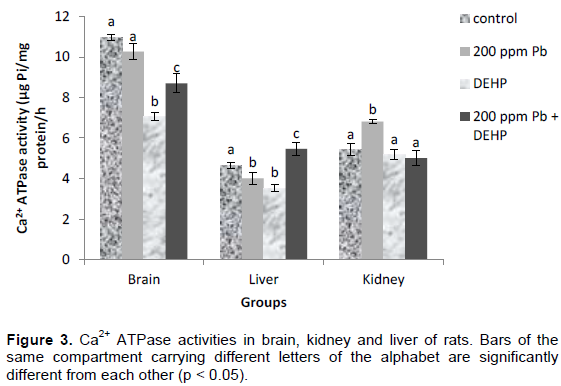
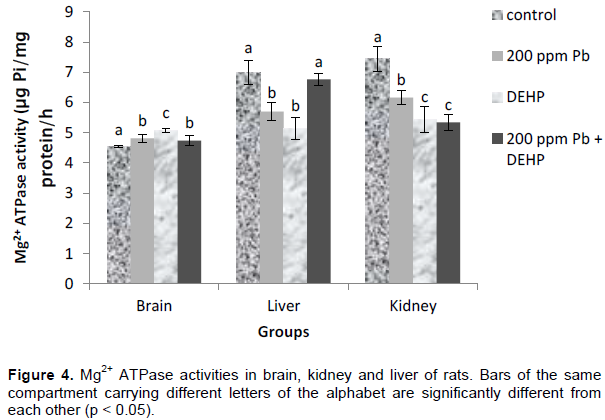
Lead, DEHP and combined lead and DEHP treatment stimulated Mg2+ ATPase activities by 6, 12 and 5 % respectively in the brain. On the contrary, liver Mg2+ ATPase activities were down-regulated by lead and DEHP treatments (19 and 26 % respectively) while co-treatment neutralized the individual inhibitions, restoring enzyme activity towards control. Kidney Mg2+ ATPase activities were inhibited by 17, 27 and 28% in the lead, DEHP and co-treatment groups respectively. Figure 4.
DISCUSSION
In this study, animals exposed to DEHP exhibited weight loss unlike the other treatment groups. This is contrary to other studies that have shown DEHP-exposed rats to have increased body weight and fat mass (Jia et al., 2016; Klöting et al., 2015). First and foremost, Klöting et al. (2015) worked with mice and not rats. Secondly, the mice were exposed to DEHP for 10 weeks, a much longer period than that employed in this study. Thirdly, the mice employed in the Klöting study were exposed to DEHP at a very low dose of 0.05 mg/kg bw/day. Hence summarily, the disparity in results could be attributed to the species of rodents used, duration of exposure and dose of DEHP administered factors that have been shown to influence the response of mammals to xenobiotics (Klaasen and Watkins, 2015). In this study, the DEHP-exposed rats displayed reduced appetite throughout the study compared to animals in the other groups and this might account for their weight loss. Reduced appetite could be attributed to inflammation-associated anorexia arising from the particular concentration of DEHP and the duration that it was administered (Wang et al., 2019). It was also observed that Pb antagonized DEHP - induced decrease in bodyweight in this study, and mechanisms that may be compensatory led to the highest increase in total body weight in this co-treated group. DEHP - induced hepatomegaly was observed and Pb had no effect on this observed hepatomegaly as there was no significant difference between relative liver weights of the DEHP and DEHP and Pb co-treatment groups. Other studies have also observed DEHP-induced hepatomegaly in rats (Musthag et al., 1980). The modes of action of DEHP in inducing hepatomegaly has been proposed to include: (1) activation of peroxisome proliferator-activated receptor-α (PPARα) (2) proliferation of peroxisomes and induction of peroxisomal proteins, (3) induction of non-peroxisomal metabolism proteins, (4) induction of cell proliferation, (5) suppression of apoptosis, (6) production of reactive oxygen species, (7) oxidative DNA damage, and (8) inhibition of gap junctional intercellular communication (Blystone et al., 2010; Mitchell et al., 1985; NTP-CERHR 2006; Rusyn et al., 2006; Zang et al., 2017; Rowdhwai and Cheng, 2018; Ito et al., 2019).
Unlike in the liver, there was brain atrophy observed in the Pb and DEHP exposed groups and this effect was additive in co-treatment. Atrophy of the brain is a common feature of lead poisoning (Cecil et al., 2008) and many of the diseases that affect the brain. Brain atrophy describes a loss of neurons and the connections between them. It could be generalized, meaning that all of the brain has shrunk; or it could be focal, affecting only a limited area of the brain resulting in a decrease of the functions that area of the brain controls. The work of Mayeux and Stern (2012) estimated the global prevalence of dementia to be as high as 24 million, and predicted it to double every 20 years until at least 2040. With the global rise in prevalence of Alzheimer’s and dementia correlating with a global rise in the use of phthalate – containing plastic products (Horn et al., 2004), and the result of this study that has shown an additive effect of lead and DEHP on brain atrophy, there is some scope in the hypothesis that world increase in prevalence of dementia in the aged could be linked to accumulation of interaction of environmental pollutants such as lead and phthalates in the system (Stewart and Schwartz, 2007).
This study has also shown that combined exposure to Pb and DEHP displayed different effects on organ ATPases from individual exposures to these toxicants. Response of the liver ATPases were especially significant and different in combined exposure. Ca2+ ATPase functions in the transfer of Ca2+ ions across cell membranes, mediating diverse physiological processes such as gene expression, contraction, secretion, fertilization, proliferation and apoptosis (Jiang and Zi-jiang, 2008; Mata and Sepulveda, 2010). One of the mechanisms through which Pb2+ exerts its toxic effects is by mimicking the action of Ca2+ in many physiological processes hence the supplementation of Ca2+ and Zn2+ has been shown to ameliorate Pb2+ toxicity (Prasanthi et al., 2006). This is because these divalent cations compete for the same binding sites as Pb2+ in the intestinal mucosa. In this study, individual exposure to Pb2+ and DEHP was found to inhibit hepatic Ca2+-ATPase - which is in consonance with the work of Abam et al. (2008) and Afolabi et al. (2016), however, co-treatment with both agents resulted in significant induction of the protein. An explanation for this increase in hepatic activity of Ca2+ ATPase could be that the interaction of Pb2+ and DEHP in co-treatment resulted in a greater bioavailability of Ca2+ ions leading to an increase in the activity of Ca2+-ATPase in order to maintain low intracellular levels of calcium. Calcium is tightly regulated because of its significance as a powerful second messenger crucial in proliferation, differentiation, mitosis, and motility (Capiod et al., 2011) hence calcium is removed to the extracellular environment and/or sequestered in the endoplasmic reticulum. Secondly, intracellular calcium is also known to be regulated by Na+K+-ATPase (Tian and Xie, 2008). For example, binding of ouabain (a well-known physiological inhibitor of Na+K+-ATPase) to the enzyme leads to increase in intracellular calcium in cardiac myocytes, resulting in increases in myocardial contraction. This is the basis for the administration of digitalis in treating congestive heart failure. Mechanistically, the Na+K+-ATPase has been shown to regulate calcium entry through the Na+/Ca2+ exchanger (NCX) by altering intracellular Na+ concentration in cardiac myocytes (Blaustein et al., 1998). Physical coupling among the Na+K+-ATPase, NCX, and sarcoplasmic reticulum calcium store has been demonstrated in smooth muscle cells (Moore et al., 1993). Other workers (Juhaszova and Blaustein, 1997; Song et al., 2006) have provided evidence that the Na+K+-ATPase interacts with NCX to form a specific calcium-signaling microdomain in many different cell types. The greater bioavailability of Ca2+ due to Pb2+ and DEHP co-treatment in the liver may have triggered an increase in NCX activity (Matsuoka, 1995) which in turn may have led to the observed increase in activity of Na+K+-ATPase activity. Renal Ca2+ ATPase activity was also observed to be upregulated by Pb2+ in this study, which could be explained as hormetic. Hormesis is the concept whereby non-nutrtional toxic substances impart beneficial or stimulatory effects at low to moderate concentrations but adverse effects at high concentrations (Klaassen and Watkins, 2015). DEHP imparted no effects on the enzyme while co-treatment with Pb2+ and DEHP produced antagonistic effects. A similar antagonistic association between Pb2+ and DEHP combined treatment was also observed in brain Ca2+ ATPase activity suggesting that the binding of one toxicant may decrease the binding of the other toxicant through a conformational change in the enzyme.
Unlike Ca2+ ATPase, individual exposures to Pb2+ and DEHP upregulated hepatic Na+K+-ATPase activities. This is contrary to various works that have recorded an inhibitory effect on the hepatic enzyme by both agents (Yücebilgiç et al., 2003; Dhanya et al., 2003; Krsti? et al., 2008; Afolabi et al., 2016). This disparity in results may be explained by differences in conditions, concentration of toxicant administered, ages and species of animals and subjects employed in the study – well established indices that have been shown to affect response to toxicants. Yücebilgiç et al. (2003) employed human subjects in their study and they observed the enzyme inhibition in the blood of the lead-exposed workers.
Although Dhanya et al. (2003) worked on rats, DEHP was administered intraperitoneally and at a much lower dose of 750 μg/kg body weight (compared to 100 mg/Kg body weight employed in this study). Krsti? et al. (2008) employed very large adult rats (average of 330 – 400 g) in their study unlike the present study that used rats with average weights of 140 g. Afolabi et al. (2016) employed rats with an average weight of 80 – 100 g (7 – 8 weeks old) unlike the present study. Co-treatment produced potentiating effects on Na+K+-ATPase activity. Three mechanisms proposed for the potentiation of Pb2+-induced activation of hepatic Na+K+-ATPase by DEHP in co-treatment are: (1) an increase in concentration of Na+ or K+ or Ca2+, (2) some natural physiologic inhibitor of Na+K+-ATPase such as ouabain may have been downregulated as a result of Pb2+ and DEHP interaction in co-treatment leading to an up-regulation in the activity of the enzymes, (3) Some compensatory mechanisms. Increase in Na+K+-ATPase activity as a result of higher bioavailability of Ca2+ has already been discussed in the preceding paragraph. In the brain, Na+K+-ATPase activities were generally downregulated by Pb2+ and DEHP (in accordance with the work of Okediran et al., 2009; Dhanya et al., 2003) while combined treatment produced no significantly different effects from the individual single exposures. Na+K+-ATPase plays a crucial role in the maintenance of the Na+ and K+ gradients across the plasma membrane of neuronal cells hence the function of these cells are hinged on the efficient regulation of the transport function of this ion pump. Research on human genetic diseases and various animal models have confirmed that neuronal activity and survival are dependent on a normal functioning of the Na+ pump. Adverse effects on the Na+,K+-ATPase pump may lead to neuronal death (Horisberger and Geering, 2009). Pb2+ had no effects on renal Na+K+ ATPase activity at the dose and duration administered, however DEHP and co-treatment caused approximately the same level of inhibition of the enzyme showing the absence of any Pb2+ effects on DEHP induced inhibition of the enzyme in the kidney.
Mg2+ ATPase activities showed organ-specific responses to Pb2+, DEHP and combined treatments with both toxicants just as already observed with the other ATPases. In the brain, Mg2+ ATPase activity was stimulated by individual exposure to Pb2+and DEHP while co-exposure to both toxicants produced an antagonistic effect of Pb2+ on DEHP. On the contrary, liver and kidney Mg2+ ATPase activities were down-regulated by lead and DEHP treatments while combined treatments with both toxicants restored enzyme activity towards control value in the liver but sustained DEHP induced inhibition in the kidney. Compensatory and adaptive mechanisms may explain the restoration of enzyme activities to control values in the liver. Such mechanisms are popular in organ responses to toxicants (Klaassen and Watkins, 2015). Total ATPase enzyme responses to Pb2+ and DEHP treatment were also different from one organ to the other. Pb2+ showed a potentiating effect on DEHP induced inhibition of total ATPase in the brain, while an antagonistic effect was observed between both toxicants in the kidney. On the contrary in the liver, a significant activation of the enzyme was observed, which was in consonant with liver Na+K+ ATPase and Ca2+ ATPase. Only in Mg2+ ATPase liver activity was the compensatory mechanism not complete in co-treatment, rather, the individualy induced inhibitions were reversed to control value.
In this study, it is observed that DEHP acted somewhat like a chelator for Pb2+ in the liver. The association between DEHP and Pb2+ seemed to relieve Pb2+ and DEHP induced inhibitions of the ATPases in the liver leading to neutralization of Pb2+ and DEHP individual inhibitions of Mg2+ ATPase, conversion of individual inhibitions of Ca2+ ATPase to a 17 % activation of the enzyme, additive increase in Na+K+ ATPase activity over and above the increase in activity of the enzyme observed in individual exposures to both toxicants. This shows that at some concentrations, combined exposures to toxicants may actually neutralize their individual toxic effects in some organs while magnifying the toxic effects in other organs.
CONCLUSION
The findings of this study demonstrate tissue specific responses of ATPases to lead and DEHP co-exposure in rat, with hepatic effects deviating significantly from the brain and renal compartments. More work is needed to evaluate the compensatory and adaptive mechanisms that could have accounted for these deviations and how this contributes to or protects from hepatic degeneration.
ACKNOWLEDGMENTS
The authors would like to express their appreciation to Dr. Olusegun Kayode Afolabi of the Biochemistry Department of Ladoke Akintola University of Technology (LAUTECH), Ogbomoso for his kind assistance in establishing this study. Also, the laboratory staff of the Biochemistry laboratory of Bells University of Technology, Ota, especially Mrs. Elizabeth Ogunbiyi and Miss Funke Ogunsakin are acknowledged for their technical support.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
REFERENCES
|
Abam E, Okediran BS, Odukoya OO, Adamson I, Ademuyiwa, O (2008). Reversal of ionoregulatory disruptions in occupational lead exposure by vitamin C. Environmental Toxicology and Pharmacology 26(3):297-304. |
|
|
Ademuyiwa O, Agarwal R, Chandra R, Behari, JR (2009). Lead-induced phospholipidosis and cholesterogenesis in rat tissues. Chemico-Biological Interactions 179(2-3):314-320. |
|
|
Ademuyiwa O, Arowolo T, Ojo DA, Odukoya OO, Yusuf AA, Akinhanmi TF (2002). Lead levels in blood and urine of some residents of Abeokuta, Nigeria. Trace elements. Electrolytes 19(2):63-69. |
|
|
Ademuyiwa O, Ugbaja RN, Idumebor F, Adebawo O (2005). Plasma lipid profiles and risk of cardiovascular disease in occupational lead exposure in Abeokuta, Nigeria. Lipids in Health and Disease 4(1):1-7. |
|
|
Ademuyiwa O, Ugbaja RN, Rotimi SO, Abam E, Okediran BS, Dosumu OA, Onunkwor BO (2007). Erythrocyte acetylcholinesterase activity as a surrogate indicator of lead-induced neurotoxicity in occupational lead exposure in Abeokuta, Nigeria. vironmental Toxicology and Pharmacology 24(2):183-188. |
|
|
Adeniyi FAA, Anetor JI (1999). Lead poisoning in two distant states of Nigeria an indication of the real size of the problem. African Journal of Medical and Health Sciences 28(1-2):107-112. |
|
|
Adeniyi TT, Ajayi GO, Akinloye OA (2008). Effect of Ascorbic acid and Allium sativum on tissue lead in female Rattus navigicus. Nigerian Journal of Health and Biomedical Sciences 7(2):38-41. |
|
|
Afolabi OK, Fatoki J, Ugbaja RN, Adekunle AS, Adedosu OT, Ademuyiwa O (2015). Arsenic and di(2- ethylhexyl) phthalate interact to alter individual effects on some oxidative stress indices in rat. Asian Journal of Pharmacology and Toxicology 3(8):24-30. |
|
|
Afolabi OK, Ugbaja RN, Ademuyiwa O (2016). Combined arsenic and di-(2-ethylhexyl) phthalate exposure elicits responses in brain ATPases different from hepatic and renal activities in rats. Journal of Toxicology and Environmental Health Sciences 8(2):6-14. |
|
|
Allouche L, Hamadouche M, Touabti A, Khennouf S (2011). Effect of Long-term Exposure to Low or Moderate Lead Concentrations on Growth, Lipid Profile and Liver Function in Albino Rats. Advances in Biological Research 5(6):339-347. |
|
|
Basaran N, Underger U (2000). Effects of lead on immune parameters in occupationally exposed workers. American Journal of Industrial Medicine 38(3):349-354. |
|
|
Blaustein MP, Juhaszova M, Golovina VA (1998). The cellular mechanism of action of cardiotonic steroids: a new hypothesisClinical and Experimental Hypertension 20(5-6):691-703. |
|
|
Blystone CR, Kissling GE, Bishop JB, Chapin RE, Wolfe GW, Foster PM (2010). Determination of the di-(2-ethylhexyl) phthalate NOAEL for reproductive development in the rat: importance of the retention of extra animals to adulthood.Toxicological Sciences 116(2):640-646. |
|
|
Bradford MM (1976). A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 7(72):248-254. |
|
|
Capiod T (2011). Cell proliferation, calcium influx and calcium channels. Biochimie 3(12):2075-2079. |
|
|
Cecil KM, Brubaker CJ, Adler CM, Dietrich KN, Altaye M, Egelhoff JC, Wessel S, Elangovan I, Hornung R, Jarvis K, Lanphear BP (2008). Decreased Brain Volume in Adults with Childhood Lead Exposure. Journal Medicine 5(5):0741-0750. |
|
|
Dhanya CR, Indu AR, Deepadevi KV, Kurup PA (2003). Inhibition of membrane Na+ K+ ATPase of the brain, liver and RBC in rats administered Di-(2-ethylhexyl) phthalate (DEHP), a plasticizer used in polyvinyl chloride (PVC) blood storage bag. Indian Journal of Experimental Biology 41(8):814-820. |
|
|
Dosumu O, Onunkwor B, Odukoya O, Arowolo T, Ademuyiwa O (2005). Biomarkers of lead exposure in auto-mechanics in Abeokuta, Nigeria. Trace Elem. Electrolyte 22(3):185-191. |
|
|
Engel SM, Miodovnik A, Canfield RL, Zhu C, Silva MJ, Calafat AM, Wolff MS (2010). Prenatal phthalate exposure is associated with childhood behavior and executive functioning. Environmental Health Perspectives 118(4):565-571. |
|
|
Evans DJ (1969). Membrane adenosine triphosphatase of Escherichia coli: activation by calcium ions and inhibition by monovalent cations. Journal of Bacteriology 100(2):914-922. |
|
|
Foster PMD, Mylchreest E, Gaido KW, Sar M (2001). Effects of phthalate esters on the developing reproductive tract of male rats. Human Reproduction Update 7(3):231-235. |
|
|
Fromme H, Kutcher T, Otto T, Pilz K, Muller J, Wenzer A (2012). Occurrence of phthalates and bisphenol A and F in the environment. Water Research 36(6):1429-1438. |
|
|
Grün F (2010). Obesogens. Current Opinion in Endocrinology, Diabetes and Obesity 17(5):453-459. |
|
|
Hesketh JE, Loudon JB, Reading HW, Glen IM (1978). The effect of lithium treatment on erythrocyte membrane ATPase activities and erythrocyte ion content. British Journal of Clinical Pharmacology 5(4):323-329. |
|
|
Hjerten S, Pan H (1983). Purification and characterization of two forms of a low-affinity Ca2+-ATPase from erythrocyte membrane. Biochimica et Biophysica Acta 728(2):281-288. |
|
|
Horisberger JD, Geering K (2009). Brain Na K-ATPase. Encyclopedia of Neuroscience. Science Direct. Elsevier Ltd. pp. 395-399. |
|
|
Horn O, Nalli S, Cooper D, Nicell J (2004). Plasticizer metabolites in the environment. Water Research 38(17):3693-3698. |
|
|
Ito Y, Kamijima M, Nakajima T (2019). Di(2-ethylhexyl) phthalate-induced toxicity and peroxisome proliferator-activated receptor alpha: a review. Environmental Health and Preventive Medicine 24(1):1-9. |
|
|
Jangid AP, Shekhawat VPS, Pareek H, Yadav D, Sharma P, John PJ (2016). Effect of lead on human blood antioxidant enzymes and glutathione. International Journal of Biochemistry Research and Review 13(1):1-9. |
|
|
Jia Y, Liu T, Zhou L, et al. Effects of Di-(2-ethylhexyl) Phthalate on lipid Metabolism by the JAK/STAT Pathway in Rats. International Journal of Environmental Research and Public Health 13(11):1085. |
|
|
Jiang T, Zi-jiang X (2008). The Na-K-ATPase and Calcium-Signaling Microdomains. Physiology 23(4):205-211. |
|
|
Juhaszova M, Blaustein MP (1997). Na+ pump low and high ouabain affinity alpha subunit isoforms are differently distributed in cells. Proceedings of the National Academy of Sciences of the United States of America 94(5):1800-1805. |
|
|
Kamrin MA (2009). Phthalate risks, phthalate regulation, and public health: a review. Journal of Toxicology and Environmental Health 12(2):157-174. |
|
|
Klaassen CD, Watkins JB (Eds) (2015). In Cassaret and Doull's Essentials of Toxicology. McGraw Hill Education, LANGE, New York P 277. |
|
|
Klöting N, Hesselbarth N, Gericke M, Kunath A, Biemann R, Chakaroun R, Kosacka J, Kovacs P, Kern M, Stumvoll M, Fischer B (2015). Di-(2-ethylhexyl)-phthalate (DEHP) causes impaired adipocyte function and alters serum metabolites. PloS one 10(12):e0143190. |
|
|
Kosnet JM, Charles E, Becker T (1998). Basic Chemical Pharmacology. 7th ed. Appleton and Langs. Pub. pp. 321-322. |
|
|
Krsti? D, Krinulovi? K, Vasi? V (2008). Inhibition of Na+K+ -ATPase and Mg2+-ATPase by metal ions and prevention and recovery of inhibited activities by chelators. Journal of Enzyme Inhibition and Medicinal Chemistry 20(5):469-476. |
|
|
Lowry OH, Lopez JA (1946). The determination of inorganic phosphate in the presence of labile phosphate esters. Journal of Biological Chemistry 162:421-428. |
|
|
Markovac J, Goldstein GW (1988). Lead activates protein kinase C in immature rat brain microvessels. Toxicology and Applied Pharmacology 96(1):14-23. |
|
|
Mata AM Sepulveda MR (2010). Plasma membrane Ca2+-ATPases in the nervous system during development and ageing. World Journal of Biological Chemistry 26(7):229-234. |
|
|
Matsuoka S, Nicoll DA, Hryshko LV, Levitsky DO, Weiss JN, Philipson KD (1995). Regulation of the cardiac Na+-Ca2+ exchanger by Ca2+ -mutational analysis of the Ca2+ - binding domain. Journal of General Physiology 105(3):403-420. |
|
|
Mayeux R, Stern Y (2012). Epidemiology of Alzheimer DiseaseCold Spring Harbor Perspectives in Medicine 2(8):a006239. |
|
|
Medesins Sans Frontiers (2012). Lead poisoning crisis in Zamfara state northern Nigeria paper. MSF Briefing Paper. May, 2012. |
|
|
Miodovnik A, Engel SM, Zhu C, Ye X, Soorya LV, Silva MJ, Calafat AM, Wolff MS (2011). Endocrine disruptors and childhood social impairment. NeuroToxicology 32(2):261-267. |
|
|
Mitchell FE, Price SC Hinton RH, Grasso P, Bridges JW (1985). Time and dose-response study of the effects on rats of the plasticizer di(2-ethylhexyl) phthalate.Toxicology and Applied Pharmacology 81(3):371-392. |
|
|
Moore ED, Etter EF, Philipson KD, Carrington WA, Fogarty KE, Lifshitz LM, Fay FS (1993). Coupling of the Na+/Ca2+ exchanger, Na+/K+ pump and sarcoplasmic reticulum in smooth muscle. Nature 365(6447):657-660. |
|
|
Mushtaq M, Srivastava SP, Seth PK (1980). Effect of di-2-ethylhexyl phthalate (DEHP) on glycogen metabolism in rat liver. Toxicology 16(2):153-161. |
|
|
National Toxicology Program - Center for the evaluation of risks to human reproduction (NTP-CERHR) (2006). Monograph on the potential human reproductive and developmental effects of di(2-ethylhexyl)phthalate (DEHP). |
|
|
Nriagu JO, Blankson ML, Ockran K (1996). Childhood lead poisoning in Africa, a growing public health problem.Science of the Total Environment 81(2):93-1000. |
|
|
Ohnishi T, Suzuki T, Suzuki Y, Ozawa K (1982). A comparative study of plasma membrane Mg2+- ATPase activities in normal, regenerating and malignant cells. Biochem. Biophys. Acta 684(1):67-74. |
|
|
Okediran BS, Abam E, Odukoya OO, Adamson I, Ademuyiwa O (2009). Membrane, intracellular, plasma and urinary sodium and potassium in occupational lead exposure: effects of vitamin C supplementation. Trace Elements and Electrolytes 26(2):49-59. |
|
|
Onunkwor B, Dosunmu O, Odukoya OO, Arowolo T, Ademuyiwa O (2004). Biomarkers of lead exposure in petrol station attendants and auto-mechanics in Abeokuta, Nigeria: effects of 2-week ascorbic acid supplementation. Environmental Toxicology and Pharmacology 17(3):169-176. |
|
|
Padmakumaran NKG, Deepadevi KV, Arun P, Manoj KV, Anitha S, Lekshmi LR, Kurup PA (1998). Toxic effect of systemic administration of low doses of the plasticiser di- (2 ethylhexyl) phthalate (DEHP) in rats. Indian Journal of Experimental Biology 36:264. |
|
|
Pracheta M, Singh L (2009). Effect of lead nitrate (Pb(NO3)2 on plant nutrition, as well as physical and chemical parameters on Lobia (Vigna unguiculata Linn. Walp.). Journal of Plant Development Sciences 1(2):49-56. |
|
|
Prasanthi RPJ, Reddy GH, Chetty CS, Reddy GR, (2006). Influence of calcium and zinc on lead-induced alterations in ATPases in the developing mouse brain. Journal of Pharmacology and Toxicology 1(3):245-252. |
|
|
Rastogi SK (2008). Renal effects of environmental and occupational lead exposure. Indian Journal of Occupational and Environmental Medicine 12(3):103-106. |
|
|
Rowdhwal SSS, Cheng J (2018). Toxic Effects of Di-2-ethylhexyl Phthalate: An Overview. Hindawi BioMed Research International pp. 1-10. |
|
|
Rusyn I, Peters J, Cunningham M (2006). Modes of action and species-specific effects of di-(2-ethylhexyl) phthalate in the liver. Critical Reviews in Toxicology 36(5):459-479. |
|
|
Sanders T, Liu Y, Buchner V, Tchounwou PB (2009). Neurotoxic Effects and Biomarkers of Lead Exposure: A Review. Reviews on Environmental Health 24(1):15-45. |
|
|
Sharma S, Sharma V, Pracheta M, Sharma SH (2011a). Therapeutic potential of hydromethanolic root extract of Withania somnifera on neurological parameters in swiss albino mice subjected to lead nitrate. International Journal of Current Pharmaceutical Research 3(2):52-56. |
|
|
Sharma V, Sharma S, Pracheta M, Sharma SH (2011b). Lead Induced hepatotoxicity in male swiss albino mice: The protective potential of the hydromethanolic extract of Withania somnifera. International Journal of Pharmaceutical Sciences Review and Research 7:116-121. |
|
|
Song H, Lee MY, Kinsey SP, Weber DJ, Blaustein MP (2006). An N-terminal sequence targets and tethers Na+ pump alpha2 subunits to specialized plasma membrane microdomains. Journal of Biological Chemistry 281(18):12929-12940. |
|
|
Stahlhut RW, van Wijngaarden E, Dye TD, Cook S, Swan SH (2007). Concentrations of urinary phthalate metabolites are associated with increased waist circumference and insulin resistance in adult U.S. males. Environmental Health Perspectives 115(6):876-882. |
|
|
Stewart WF, Schwartz BS (2007). Effects of lead on the adult brain: a 15-year exploration. American Journal of Industrial Medicine 50(10):729-739. |
|
|
Tian J, Xie ZJ (2008). The Na+-K+-ATPase and calcium-signaling microdomains. Physiology 23:205-211. |
|
|
Wang Y, Kim J, Schmit MB, Cho TS, Fang C, Cai H, (2019). A bed nucleus of stria terminalis microcircuit regulating inflammation-associated modulation of feeding. Nature Communications 10:2769. |
|
|
Wójtowicz AK, Sitarz-G?ownia AM, Szcz?sna M, Szychowski KA (2019). The Action of di-(2-ethylhexyl) phthalate (DEHP) in mouse cerebral cells involves an impairment in aryl hydrocarbon receptor (AhR) signaling. Neurotoxicity Research 35(1):183-195. |
|
|
Xu J, Ling-jun L, Chen WU, Xiao-feng W, Wen-yu FU, Lihong X (2008). Lead induces oxidative stress, DNA damage and alteration of p53, Bax and Bcl-2 expressions in mice. Food and Chemical Toxicology 46(5):1488-1494. |
|
|
Ye H, Ha M, Yang M, Yue P, Xie Z, Liu C (2017). Di2-ethylhexyl phthalate disrupts thyroid hormone homeostasis through activating the Ras/Akt/TRHr pathway and inducing hepatic enzymes. Scientific Reports 7:40153; |
|
|
Yücebilgiç G, Bilgin R, Tamer L, Tükel S (2003). Effects of lead on Na+K+ ATPase and Ca2+ ATPase activities and lipid peroxidation in blood of workers. International Journal of Toxicology 2(2):95-97. |
|
|
Zhang W, Shen XY, Zhang WW, Chen H, Xu WP, Wei W (2017). The effects of di 2-ethyl hexyl phthalate (DEHP) on cellular lipid accumulation in HepG2 cells and its potential mechanisms in the molecular level. Toxicology Mechanisms and Methods 27(4):245-252. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


