Full Length Research Paper

ABSTRACT
Although iron is an essential element for life, its excess is linked to many disorders. The aim of this study is to investigate the effect of quercetin on iron induced toxicity on the heart and brain in adult male albino rats. 50 adult male albino rats were equally divided into groups; Group I (negative control), Group II (positive control) received normal saline 0.9%. Group III (Quercetin group) was treated with quercetin (2 g/kg daily) for 8 weeks and Group IV (Iron group) was injected intraperitoneal daily with iron dextran (300 mg/kg) for 4 weeks. Group V (Iron and quercetin group) was injected intraperitoneal daily with iron dextran (300 mg/kg) for 4 weeks, and then gavaged orally with quercetin (2 g/kg once a day) for another 4 weeks. At the end of the study, blood samples were taken to estimate the serum levels of iron, malondialdehyde (MDA), and total antioxidant capacity (TAC). Heart and brain specimens were dissected from scarified rats to estimate tissue level of iron, histopathological examination, immunohistochemical staining for tumor necrosis factor (TNFα) and comet assay. Iron overload caused increases in serum, heart and brain iron levels; increase in serum MDA and decrease in serum TAC with degenerative changes in the examined tissues and increasing expression of TNFα and DNA degradation. After administration of quercetin, a significant improvement in all these parameters was observed. Quercetin acts as iron chelator decreasing serum and tissue iron levels, ameliorating oxidative stress, inflammatory effects and DNA damage induced by iron overload. More studies are recommended to evaluate the beneficial effects of quercetin with iron excess.
Key words: Iron, quercetin, heart toxicity, brain toxicity.
INTRODUCTION
Iron is one of the essential nutrients (Pari et al., 2015). It has two states, ferrous and ferric states that can turn reversibly between these forms (Halliwell and Gutteridge, 1990). It enters in composition of different elements in human body as hemoglobin, myoglobin and many enzymes implicated in redox reactions and metabolism of energy (Beutler, 2006). So that, iron is an important element in many procedures such as RCBs production, carrying oxygen, and mitochondrial respiration (AbdElkader and Aly, 2015).
There is no definite way to get rid of excess iron except regulating its absorption which mainly occurs in the small Bowel to keep body iron level normal (Goodman and Gilman, 2006; Anderson, 2007). So, excess oral intake of iron for a long interval results in its accumulation in various organs causing serious adverse effects as heart failure, liver cirrhosis, kidney injury, diabetes, depression, infertility and even cancer (Kohgo et al., 2008; Kulaksiz et al., 2008; Gholampour et al., 2017). Iron has an important role in production and removal of reactive oxygen species (Frey and Reed, 2012). By Fenton reaction, it can create reactive oxygen species (Kehrer, 2000). Excessive amounts of oxygen species are produced as in cases of iron overload leading to oxidative stress and cell damage (Pari et al., 2015; Gholampour et al., 2017).
Iron chelation therapy is an effective life-saving treatment in diseases created by iron excess (AbdElkader and Aly, 2015). Several compounds, either natural or synthetic can chelate and remove iron from the tissues. Although deferoxamine is a good iron chelator and has also antioxidant property, high doses of it can produce frightened side effects as growth retardation, ototoxicity and ocular toxicity as well as bone deformities (Porter, 1997).
Quercetin is an example of naturally occurring flavonoids involved under a category named flavonols. Different fruit and vegetables contain quercetin as apples, green tea, onions, nuts, berries, broccoli, cauliflower, and cabbage. Many in vivo and in vitro studies have shown that quercetin could have diverse effects like anti-inflammatory, anti-cancer, immunomodulatory, anti-diabetic, anti-aggregatory, and vasodilating effects (Kleemann et al., 2011; Sinha et al., 2014; Qi et al., 2017). Also, it is considered as a powerful antioxidant agent. It can antagonize cell damage induced by oxidative stress by chelating metals, scavenging free radicals and protecting from lipid peroxidation (Bu et al., 2011).
Multiple experimental studies had demonstrated the protective effects of quercetin against toxicity from different toxins, such as organophosphorus compounds, poly aromatic hydrocarbons, and metals (Barcelos et al., 2011). This work aimed to investigate the ability to use oral quercetin as a natural substance in treatment of iron overload in rats through measuring serum iron level, tissue iron level, serum levels of MDA, and TAC, histopatholgical examination of heart and brain tissues, immunohistochemical staining for TNF-α and estimating DNA damage by comet assay.
MATERIALS AND METHODS
Chemicals
Iron dextran (fercayl ample 100 mg/2 ml) was manufactured by Sunny Pharmaceutical, Egypt under license of Sterop Laboratoires,
Belgium. Quercetin (yellow powder) was obtained from Sigma, Aldrich in Germany imported by Cairo Chemical Company. Reagents and commercial kits were supplied from Sigma and Biodiagnostic Chemical Companies.
Experimental animals
Fifty adult male Wister albino rats (10 to 12 weeks) weighing 190 to 210 g were obtained from the breading animals house of the Faculty of Veterinary Medicine, Zagazig University. The experimental rats were put in suitable cages supplied with water and food and left for two weeks for adaptation. The experiment was carried out in compliance with the “Guide of the Care and Use of Laboratory Animals” and the institutional guidelines for the care and use of experimental animals approved by the Medical Research Ethics Committee of Zagazig University, Egypt (Institute of laboratory animals resources, 1996).
Experimental design
(1) Group I (Negative control): 10 Rats received only regular diet and tap water for 8 weeks to measure the basic parameters.
(2) Group II (Positive control): 10 Rats orally gavaged by normal saline (0.9% Nacl) (1 ml/kg/day) (the solvent of quercetin) for 8 weeks.
(3) Group III (Quercetin group): 10 rats were orally gavaged by quercetin (2 g/kg b.w daily dissolved in 1 ml of normal saline) (Zhang et al., 2006) for 8 weeks.
(4) Group IV (Iron group): 10 rats were injected intraperitoneal daily with iron dextran (300 mg/kg) (1/10 of LD50) for 4 weeks (Berdoukas et al., 2013).
(5) Group V (Iron and Quercetin group): 10 rats were injected intraperitoneal daily with iron dextran (300 mg/kg) for 4 weeks, then received daily quercetin orally by gavage (2 g/kg b.w) for another 4 weeks.
Biochemical studies
Twenty-four hours after the last dose, the rats of all groups were anesthetized with light ether anesthesia; venous blood samples were obtained from them by means of capillary glass tubes from the retro-orbital venous as described by Joslin (2009). After centrifuging blood samples, sera were obtained and used for estimating the following:
(1) Serum Malondialdehyde and Total Antioxidant Capacity assay using colorimetric method as described by Ohkawa et al. (1979) and Koracevic and Koracevic (2001) and the data were expressed as nmol/ml and mM/L, respectively.
(2) Serum iron level (ðœ‡mol/L) according to the method described by Stookey (1970).
Heart and brain iron level
Rats were sacrificed by decapitation; the brain and the heart were obtained to assess the tissues iron concentrations. Portion of brain and heart tissue of each rat underwent digestion by acid mixture (3 ml Nitric acid: 2 ml Perchloric acid); the remaining dissolving metal matter and resultant solution were aspirated into Atomic Absorption Spectrophotometer (Buck scientific 210VGP Atomic Absorption Spectrophotometer) for determination of iron level (Zantopoulos et al., 1996) and Perkin Elmer model (spectra- AA10, USA) flame atomic absorption spectrometer with computer system was employed. The data were expressed as mg/g wet weight (ppm).
Histopathological studies
Light microscope technique
The brain and heart tissues were fixed in 10% formalin saline. After fixation, they were embedded in paraffin blocks and processed for the preparation of 5 µ thickness sections. These sections were stained with Hematoxylin and Eosin (H&E) stains and examined by light microscope (Bancroft and Stevens, 1996).
Immuno-histochemical study
The sections were deparaffinized, rehydrated, and heated in a microwave oven in 0.01 M citrate buffer (pH 6.0; Química Contemporânea, Diadema, Brazil) for 30 min. Endogenous peroxidase activity was blocked by 3% hydrogen peroxide for 10 min, followed by washing with phosphate buffered saline. The sections were incubated overnight at 4°C with the following primary antibodies: Anti-TNF-α (rabbit polyclonal IgG, 100 μg/ml, 1:50 dilution, cat. no. sc-130220; Santa Cruz Biotechnology, Inc., Dallas, TX, USA) followed by addition of secondary antibody (Vector Laboratories, Burlingame, CA, USA). Slides were counterstained with Harris’s hematoxylin, dehydrated, cleared and mounted. The cells were initially observed at a low magnification (×200) to assess the general distribution of the antibody. The samples were subsequently examined at a higher magnification (×400). Immunohistochemical staining was evaluated in the cytoplasm of brain and heart tissues (Jammal et al., 2015).
Comet assay (alkaline single-cell gel electrophoresis)
Comet assay was performed according to Singh et al. (1988).100 mg of crushed brain and heart samples was suspended in 1ml ice cold PBS, stirred for 5 min and filtered. 100 μl of cell suspension was carefully mixed with 600 μl of low-melting agarose, followed by spreading of 100 μl of the mixture on agarose pre-coated slides. The slides were left to solidify at 4°C, and then they were immersed in chilled lysing solution for 1 h at 4°C. The slides were removed and sited in a horizontal electrophoresis chamber, filled with freshly prepared electrophoretic alkaline buffer for 20 min. After electrophoresis, the slides were washed gently in 0.4 M Tris–HCl buffer and stained with ethidium bromide. The DNA migration patterns of 100 cells for each sample were observed using fluorescence microscope, and images were captured by a Nikon CCD camera. The qualitative and quantitative amount of DNA damage in the cells was estimated using the Comet 5 image analysis software developed by Kinetic Imaging Ltd. (Liverpool, UK) linked to a CCD camera was used to assess the quantitative and qualitative extent of DNA damage in the cells by measuring the length of DNA migration [tail length] and the percentage of migrated DNA in the tail [tail DNA %]. Finally, the program calculates tail moment [correlation between tail length and tail DNA %]. Generally, images of 100 (50 X 2) randomly selected cells are analyzed per sample. The mean value (for 100 cells) was calculated (Ali et al., 2017). Data for all groups were collected, computerized and statistical analysis by statistically analyzed by using statistical package for social science (SPSS) program.
RESULTS
No statistically significant differences were observed in the studied parameters between negative, positive control and quercetin groups, so we used negative control group as a standard reference for comparison with other treated groups.
Biochemical results
Iron group of this work showed a highly significant elevation in the serum iron level in comparison with control group (P<0.001). Meanwhile, there was a highly significant reduction in serum iron level (P< 0.001) (iron + quercetin group) when compared to that of iron group. This reduction was non- significant compared with that of the control group (P>0.05) (Table 1). As regard iron level in tissues, there were highly significant elevations in the mean values of iron level in brain and heart tissues in iron group when compared with other groups of the study (P<0.001). There was a highly significant reduction in the mean values of iron level in brain and heart tissues
(P< 0.001) in (iron + quercetin group) when compared with iron group. When iron + quercetin group compared with (-ve) control group, there was a highly significant elevation in the mean values of iron level in brain tissues (P< 0.001) and non significant changes in heart tissues (P> 0.05) at the end of the study (Table 1). At the end of the study, serum MDA was increased significantly with significant decrease in serum levels of TAC in iron group compared to the other groups (P<0.001). Comparing with iron group, treatment with quercetin after iron intake (group V) showed significant reduction in MDA and significant elevation in the mean values of TAC (P<0.001); however, these improvements were partial when iron + quercetin group was compared with (-ve) control group; there were still a highly significant elevation in MDA and a highly significant reduction in TAC (p< 0.001) (Table 1).
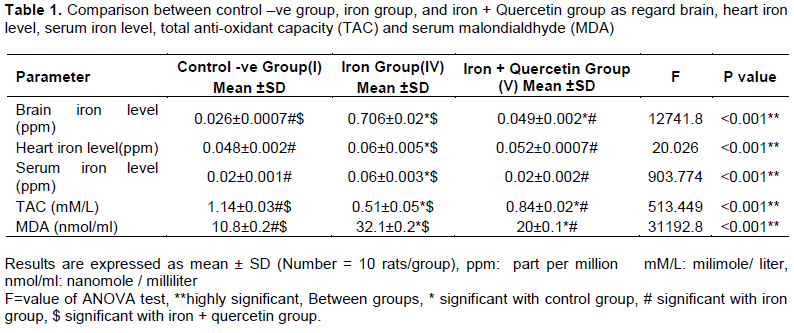
Histopathological examinations
Examination of H&E stained heart sections of control –ve, control +ve and quercetin groups showed normal heart tissues (Figure 1A). After 4 weeks of iron administrations, the heart tissue sections showed hypertrophy and degeneration of muscle fibers (Figure 1B and C). While after another 4 weeks of treatment with quercetin, there was mild hypertrophy and reversal of degenerative changes of muscle fibers in heart sections (Figure 1D).
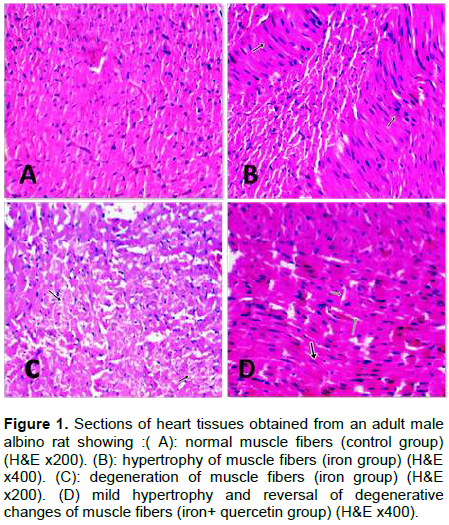
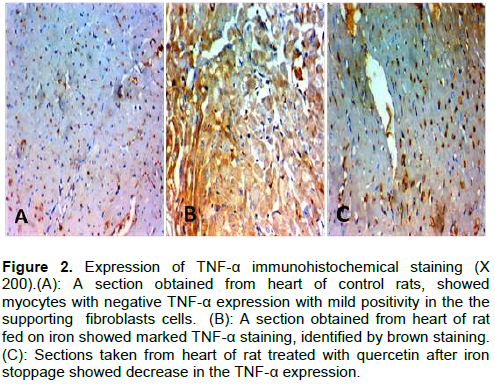
By immunohistochemical staining for TNF-α, the control and quercetin groups showed negative TNF-α expression with mild positivity in the supporting fibroblasts cells (Figure 2A). However, with iron administration, there was marked TNF-α staining (Figure 2B). This marked staining decreased after quercetin treatment for 4 weeks after cessation of iron administrations (Figure 2C). As regard brain tissues, H&E stained sections of control –ve, control +ve and quercetin groups showed normal brain tissues (Figures 3A and B). After 4 weeks of iron administrations, the brain tissue sections showed degenerative changes in neurons with thinning of hippocampus (Figure 3C and D). While after another 4 weeks of treatment with quercetin, there was an improvement in these degenerative changes (Figure 3E).
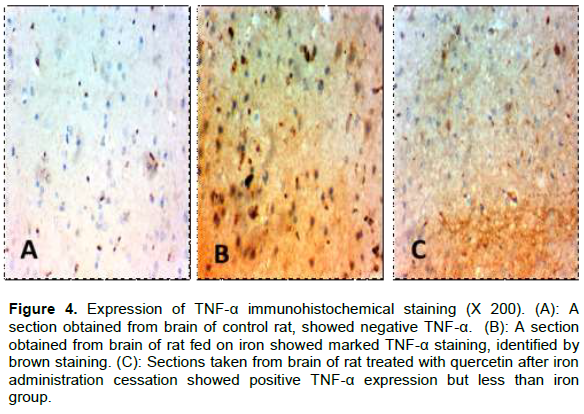
By immunohistochemical staining for TNF-α, the control and quercetin groups showed negative TNF-α expression (Figure 4A), but with iron administration, there was marked TNF-α staining (Figure 4B). This marked staining decreased after quercetin administration for another 4 weeks after cessation of iron administration (Figure 4C).
Comet assay
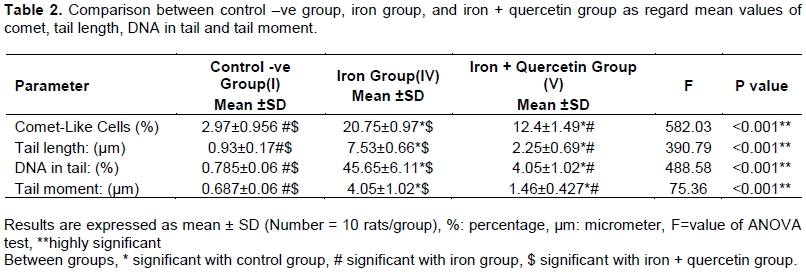
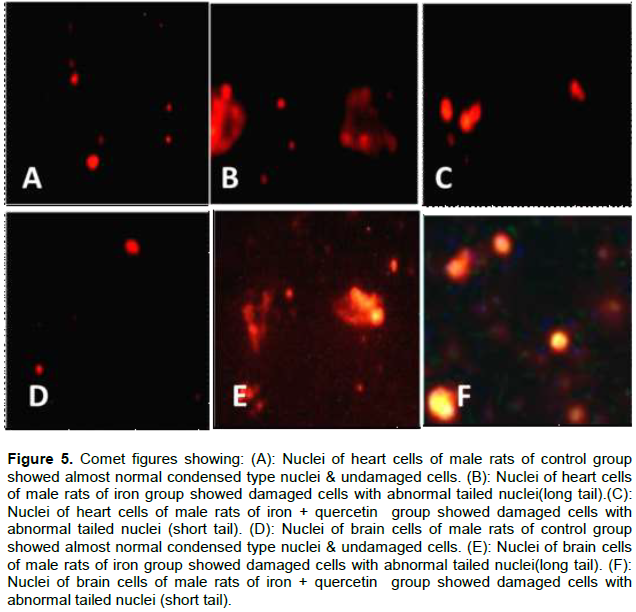
The brain and heart tissues of the rats treated with iron revealed highly significant increased DNA degradation represented in the following parameters: increase in the frequency of comet-like cells, comet tail-length, percentage of tail DNA, comet tail moment when compared to other groups (P< 0.001) (Table 2, Figure 5B and E). While after administration of quercetin for another 4 weeks after stopping iron administration, there were highly significant reductions in comet-like cells, comet tail-length, percentage of tail DNA, comet tail moment (P< 0.001) when compared with iron group (P< 0.001). However, there were still significant elevations in these parameters when iron + quercetin group was compared with (-ve) control group (P< 0.001) (Table 2, Figure 5C and F). Tail moment results from an equation between comet tail length and tail DNA%. It is the most important factor in comparison among tested groups in revealing DNA damage as it is affected by any change in both tail length and tail DNA%.
DISCUSSION
Although iron is an important element for life, different disorders including neurodegenerative diseases, cardiovascular disease and cancer were found to be related to excess free-iron in the body. Under physiological conditions, free iron reacts with peroxide and superoxide ions and enhances the formation of the pro- inflammatory hydroxyl radical (Kozlowski et al., 2009; Torti and Torti, 2013).
Desferal is a well-known iron chelator for a long time, but it has many side effects (Porter, 1997). Quercetin is a natural occurring flavonoid with multifunctional benefits including its antioxidant capacity (Qi et al., 2017). Moreover, it has been studied by many researchers as a method to chelate various metals like copper, magnesium, zinc, lead, and others (Raza et al., 2016). So that, the aim of the current work was to investigate the ameliorating and chelating effect of quercetin on iron induced toxicity on heart and brain through evaluation some biochemical iron indices, histopathological, immunohistochemical changes and comet assay.
The significant rise in serum iron and tissue (brain and heart) iron levels was a proof of iron toxicity in the present study. Iron group of this study showed highly significant elevations in the iron level in brain and heart tissues when compared with other groups; while after administration of quercetin, a significant reduction in these levels in comparison to iron group was seen. These results are in line with many studies which noted that quercetin could suppress iron overload-induced injury and therefore it may have properties of an iron chelator (Leopoldini et al., 2006; Zhang et al., 2011; Raza et al., 2016; Horniblow et al., 2017; El-Sheikh et al., 2018).
Quercetin structurally consists of three phenolic rings including A, B and C rings. It possesses three places available to chelate metals involving 3-hydroxy-4-keto group, 5-hydroxy-4-keto group and ortho-dihydroxyl (catechol) groups of B ring. Both hydroxy and keto groups of quercetin have ability to bind with metals forming metal complexes (Torreggiani et al., 2005; Ahmadi et al., 2011) which shuttled through glucose transporter 1 diminishing free radical formation (Baccan et al., 2012).
One of the broadly used biomarker of lipid peroxidation is MDA. It is a good indicator of ROS production with oxidative stress occurrence (Ayer et al., 2014). Total antioxidant capacity (TAC) acts as an indicator of all antioxidants either enzymatic or non-enzymatic accumulated in different tissues of the body which balance the effect produced by reactive oxygen and nitrogen species (Valko et al., 2006). With xenobiotic exposure, changes in their concentrations happen fast (Yang et al., 2012).
Our findings in this investigation emphasized the ability of iron overload to create oxidative stress with alterations of antioxidants as there was a high elevation in serum MDA and a high significant reduction in serum TAC in iron group compared to their corresponding values in other groups; there was a significant reduction in serum MDA and a highly significant elevation in serum TAC after quercetin administrations in this work. All these are in agreement with the findings of Abd El-Baky et al. (2009); El-Maraghy et al. (2009); Lebda (2014); Abdelkader and Aly, 2015); El-Sheikh et al. (2018) who confirmed the capacity of iron overload in provoking oxidative stress with disturbance in antioxidants. Quercetin can reduce reactive oxygen species and inhibit oxidative stress caused by iron by directly chelating iron, and disabling iron’s ability to take part in redox reactions (Uzun et al., 2010; Uzun and Kalender, 2013).
Anti-oxidant properties of quercetin was referred to for their interference with inducible nitric-oxide synthase activity (iNOS) which react with free radicals producing the highly damaging peroxynitrite inducing irreversible cell membrane damage (Nijveldt et al., 2001; Li et al., 2016a).
In concomitant with the biochemical results, our histological result revealed hypertrophy and degeneration of cardiac muscle fibers and degenerative changes in neurons with thinning of hippocampus in iron treated group; while, there was an improvement in these degenerative changes in both heart and brain tissues after treatment with quercetin. These results are consistent with Elseweidy and Abd El-Baky (2008) in which iron overload conditions were obtained by feeding the rats with biscuits enriched with ferrous sulphate for 10 weeks. Increasing iron level might accelerate dopamine oxidation with subsequent quinone formation which degenerates the brain cells and severs meningeal hemorrhage, congestion and edema.
In human beings, Kell (2010) observed iron deposition in specific brain regions mainly in the cerebral cortex and hippocampus in patients with chronic neurodegenerative diseases as in the case of Alzheimer’s disease. Also another study by Hametner et al. (2013) reported abnormal distribution of iron in the brains of multiple sclerosis patients with amplifying demyelination and neurodegeneration lesion in brain.
Iron can induce oxidative stress, combined with defective antioxidant capacities. This oxidative stress is able to damage cell membranes, DNA, and mitochondria, which are involved in neuronal death and neurodegeneration pathogenesis (Nunez et al., 2012; Uranga and Salvador, 2018). The results of the present study are also supported by Madhusudhan and Oberoi (2011); Torti and Torti (2013); Lebda (2014); Abdelkader and Aly (2015); Gholampour et al. (2017) who stated that administration of iron resulted in multiple organs dysfunction and histological observations including heart. Thalassemia major occurs, a condition of iron overload where iron accumulates in cardiac muscle cells and leads to cardiac cellular damage and heart failure causing high rate of death in these patients (Kremastinos et al., 2010). By activating various cell signaling pathways, quercetin plays key roles in the prevention of disorders, which are mainly responsible for the development of cancers, neurodegenerative and cardiovascular diseases (Khan et al., 2016).
Tumour necrosis factor-alpha (TNF-α) is a cytokine that is secreted by a group of cells like macrophages, lymphocytes, smooth muscle cells, and tumour cells. In brain, it is released by astrocytes, microglial cells, and neurons. It plays a key role in the regulation of many processes, such as inflammation, development, and apoptosis (Howerton and Tarzami, 2017).
Regarding the result of immunohistochemical staining for TNF-α, iron group showed marked TNF-α staining in both brain and cardiac tissues. This marked staining decreased after quercetin treatment. These data are in agreement with Zivadinov et al. (2011); von Bernhardi and Eugenin (2012) who suggested that excess iron accumulation promotes adverse effects of inflammatory cytokines including TNF-α, which is involved in the incidence of neurodegenerative diseases, Parkinson’s disease and Alzheimer’s disease. Changes in divalent metal transporter 1 (DMT1) and ferroportin 1 (FPN1) expression mediated by hepcidin and caused by iron accumulation in brain cells confirmed its relation with inflammation in brain tissues. This connection may have a role in the progression of neurodegenerative diseases by enhancing iron-induced oxidative damage (Urrutia et al., 2013). It was also identified that myocardial TNF-α is an important contributor to many cardiac disorders such as myocardial contractile dysfunction, cardiomyocyte death after myocardial ischemia and chronic heart failure (Howerton and Tarzami, 2017).
Several in vitro studies that used different cell lines had shown that quercetin inhibited lipopolysaccharide (LPS)-induced TNF-α production in macrophages (Gerates et al., 2007). In glial cells, quercetin can inhibit LPS-induced mRNA levels of TNF-α and interleukin (IL)-1, diminishing neuronal cell apoptosis induced by microglial activation (Bureau et al., 2008). Moreover, It can inhibit Fc"RI-mediated release of pro-inflammatory cytokines, tryptase and histamine from human umbilical cord blood-derived cultured mast cells (hCBMCs), leading to inhibiting calcium influx and phospho-protein kinase C (Kempuraj et al., 2005). It was found that quercetin can prevent activation of a group of potent inducers of inflammatory gene expression and protein secretion including TNFα such as signal related kinase, c-Jun, NH2 terminal kinase (JNK), and nuclear factor-κB (NF-κB). Quercetin can increase peroxisome proliferator activated receptor c activity, by antagonizing NF-κB or activator protein-1(AP-1) transcriptional activation of inflammatory genes, leading to prevention of inflammation. Together, these prohibit TNF-α mediated inflammatory response (Li et al., 2016b).
The alkaline single-cell gel electrophoresis (comet assay) is one of the most sensitive and rapid techniques for quantifying DNA lesions in mammalian cells. It has wide applications in the assessment of different types of DNA damage and repair and has been widely used in the genetic toxicology and environmental biomonitoring, and cancer research studies (Singh et al., 1988; Fairbairn et al., 1995). Comet assay has many advantages including high sensitivity for detecting little DNA damage, the need of small number of cells, its flexibility, low cost and ease of application (Tice et al., 2000).
The current study showed that exposure to iron signiï¬cantly caused marked DNA damage in heart and brain rat tissues evidenced by significant increase in the mean values of comet tail length, tail DNA% and tail moment while the administration of quercetin ameliorated that effect evidenced by significant decrease in the mean values of comet tail length, tail DNA% and tail moment in brain and cardiac tissue in iron and quercetin group. Iron plays a key role in the generation of ROS which reacts directly with proteins, lipids and nucleic acids and induces oxidative stress by depleting cellular stores of antioxidants. ROS also influences multiple cell signaling pathways important for cell survival, proliferation and death (Valko et al., 2005).
Several studies demonstrated that DNA damage was created by iron using comet assay. Matos et al. (2001) demonstrated that ferric nitrilotriacetate induced oxidative stress with hepatocytes apoptosis, DNA damage and liver necrosis in rats. Also, in mice exposed to ferrous sulfate by a dose of 12% of the LD50, they revealed genotoxicity after 24 h of treatment, DNA damage in the comet assay (24 h and 48 h) and micronucleus test (48 h) (Horta et al., 2016). The results of the present study are supported by the results of Chan et al. (2014) who showed that the use of quercetin could prevent trichostatin A induced DNA damage and lipid peroxidation. Another study by El-Nekeety et al. (2014) showed the protective action of quercetin on the changes of genes expression and oxidative stress induced by aflatoxin in hepatic tissues of rats.
Moreover, Qi et al. (2017) reported the protective effect of quercetin against DNA damage induced by organophosphorus compounds. This protective effect was explained by antioxidant effect of quercetin.
CONCLUSION
This study showed that quercetin has a potential role as iron chelator by decreasing serum and tissue iron levels, its ameliorative effect on oxidative stress, inflammatory effects and DNA damage induced by iron overload.
RECOMMENDATION
In the light of this study, we recommended more studies on qurecetin especially in comparison with desferal in chronic iron toxicity. Also there should be great use of natural metal chelators as querecetin with good outcome and lower side effects than the synthetics ones.
CONFLICT OF INTERESTS
The authors have not declared any conflict of interests.
ACKNOWLEDGEMENTS
Special thanks to Dr/ Eman Al-Shahat (assistant professor of community medicine, Zagazig University) for her great help in statistical analysis.
REFERENCES
|
Abd El-Baky A, Abdullah A, Abd-El Hay E (2009). Amelioration of iron-overload adverse effect by iron chelator in rats. Journal of Applied Sciences Research 5:1155-1162. |
|
|
AbdElkader A, Aly F (2015). Protective Effect of Melatonin Against Iron Overload-Induced Toxicity In Rats. International Journal of Pharmacy and Pharmaceutical Sciences 7(9):116-121. |
|
|
Ahmadi M, Dehghan G, Hosseinpourfeizi A, Dolatabadi N, Kashanian S (2011). Preparation, characterization, and DNA binding studies of water-soluble quercetin–molybdenum (VI) complex. DNA and Cell Biology 30(7):517-523. |
|
|
Ali M, Gomaa M, Shalby S, Mohammed Z (2017). Study of chronic toxic effect of deltamethrin and dimethoate on brain of adult male albino rats. Zagazig Journal of Forensic Medicine 15(1):29-46. |
|
|
Anderson J (2007). Mechanisms of iron loading and toxicity. American Journal of Hematology 82:1128-1131. |
|
|
Ayer A, Mario M, Sandro A (2014). Lipid Peroxidation: Production, Metabolism, and Signaling Mechanisms of Malondialdehyde and 4-Hydroxy-2-Nonenal Oxidative. Med. Cell. Longevity, Article ID 360438, 31. |
|
|
Baccan M, Chiarelli-Neto O, Pereira S, Espósito P (2012). "Quercetin as a shuttle for labile iron," Journal of Inorganic Biochemistry 107(1):34-39. |
|
|
Bancroft D, Stevens A (1996). Theory and Practice of Histological Techniques. (4thEd.).Churchill Livingstone Press, Edinburgh, London, Melbourne, New York pp. 99-112. |
|
|
Barcelos R, Grotto D, Serpelon M, Angeli P, Rocha A, de Oliveira Souza C, Vicentini T, Emanuelli T, Bastos K, Antunes M, Knasmüller S, Barbosa J (2011). Protective properties of quercetin against DNA damage and oxidative stress induced by methylmercury in rats. Archives of Toxicology 85:1151-1157. |
|
|
Berdoukas V, Nord A, Carson S, Puliyel M, Hofstra T, Wood J, Coates D (2013). Tissue iron evaluation in chronically transfused children shows significant levels of iron loading at a very young age. American Journal Of Hematology 88:283-285. |
|
|
Beutler E (2006). Hemochromatosis: genetics and pathophysiology. Annual Review of Medicine 57:331-347. |
|
|
Bu T, Mi Y, Zeng W, Zhang C (2011). Protective effect of quercetin on cadmium-induced oxidative toxicity on germ cells in male mice. Anatomical Record 294:520-526. |
|
|
Bureau G, Longpre F, Martinoli G (2008). Resveratrol and quercetin, two natural polyphenols, reduce apoptotic neuronal cell death induced by neuroinflammation. Journal of Neuroscience Research 86:403-410. |
|
|
Chan T, Lin C, Chuang H, Shiau J, Liao W, Yeh L (2014). "Oral and Intraperitoneal Administration of Quercetin Decreased Lymphocyte Dna Damage and Plasma Lipid Peroxidation Induced by Tsa in Vivo," BioMed Research International. |
|
|
El-Maraghy A, Rizk M, El-Sawalhi B (2009). Hepatoprotective potential of crocin and curcumin against iron overload-induced biochemical alterations in rat. African Journal of Biochemistry Research 3:215-221. |
|
|
El-Nekeety A, Abdel-Azeim H, Hassan M, Hassan S, Aly E, Abdel-Wahhab A (2014). "Quercetin inhibits the c ytotoxicity and oxidative stress in liver of rats fed aflatoxin-contaminated diet," Toxicology Reports 1:319-329. |
|
|
Elseweidy M, Abd El-Baky E (2008). Effect of dietary iron overload in rat brain: oxidative stress, neurotransmitter level and serum metal ion in relation to neurodegenerative disorders. Indian Journal of Experimental Biology 46(12):855-858. |
|
|
El-Sheikh A, Ameen H, AbdEl-Fatah S (2018). Ameliorating Iron Overload in Intestinal Tissue of Adult Male Rats: Quercetin vs Deferoxamine. International Journal of Toxicology 2018. |
|
|
Fairbairn W, Olive L, O'Neill L (1995). The comet assay: A comprehensive review. Mutation Research 339:37-59. |
|
|
Frey A, Reed H (2012). The ubiquity of iron. ACS Chemical Biology 7:1477-1481. |
|
|
Gerates L, Moonen J, Brauers K, Wouters M, Bast A, Hageman J (2007). Dietary flavones and flavonols are inhibitor of poly (ADP-ribose) polymerase-1 in pulmonary epithelial cells. Journal of Nutrition 137:2190-2195. |
|
|
Gholampour F, Ghiasabadi B, Owji M, Vatanparast J (2017). The protective effect of hydroalcoholic extract of Ginger (Zingiber officinale Rosc.) against iron-induced functional and histological damages in rat liver and kidney. Avicenna Journal of Phytomedicine 7(6):542-553. |
|
|
Goodman S, Gilman A (2006). The Pharmacological Basis of Therapeutics.11th edition. New York: McGraw-Hill. |
|
|
Halliwell B, Gutteridge M (1990). Role of free radicals and catalytic metal ions in human disease: An overview.Methods in Enzymology186: 1-85. |
|
|
Hametner S, Wimmer I, Haider L, Pfeifenbring S, Br€uck W, Hans H (2013). Iron and Neurodegeneration in the Multiple Sclerosis Brain. Annals of Neurology 74(6):1-14. |
|
|
Horniblow D, Henesy D, Iqbal H, Tselepis C (2017). Modulation of iron transport, metabolism and reactive oxygen status by quercetin-iron complexes in vitro. Molecular nutrition and food research 61(3):1600692. |
|
|
Horta N, Kahl F, Sarmento S, Nunes S, Maglione R, Andrade M FF, Da Silva J (2016). Protective effects of acerola juice on genotoxicity induced by iron in vivo Molecular genetics. Biology 39(1):122-128. |
|
|
Howerton E, Tarzami T (2017). Tumor Necrosis Factor-Alpha and Inflammation-Mediated Cardiac Injury. Journal of Cell Science and Therapy 8:268. |
|
|
Institute of laboratory animals' resources, Commission on life sciences, National research council (1996). Guide for the care and use of laboratory animals. National Academy Press. |
|
|
Jammal M, Da silva A, Filho M, Côbo C, Adad J, Murta C, Nomelini S (2015). Immunohistochemical staining of tumor necrosis factor-α and interleukin-10 in benign and malignant ovarian neoplasms. Oncology Letter 9(2):979-983. |
|
|
Joslin J (2009): Blood collection: Techniques in exotic small mammals. Journal of Exotic Pet Medicine 18(2):117-139. |
|
|
Kehrer P (2000). The Haber–Weiss reaction and mechanisms of toxicity. Toxicological 149:43-50. |
|
|
Kell B (2010).Towards a unifying, systems biology understanding of large-scale cellular death and destruction caused by poorly liganded iron: Parkinson's, Huntington's, Alzheimer's, prions, bactericides, chemical toxicology and others as examples," Archives of Toxicology 84(11):825-889. |
|
|
Kempuraj D, Madhappan B, Christodoulou S, Boucher W, Cao J, Papadopoulou N, Cetrulo L, Theoharides C (2005). Flavonols inhibit proinflammatory mediator release, intracellular calcium ion levels and protein kinase C theta phosphorylation in human mast cells. British Journal of Pharmacology 145:934-944. |
|
|
Khan F, Niaz K, Maqbool F, Hassan H, Abdollahi M, Kalyan C, Venkata N, Nabavi M, Bishayee A (2016). A Molecular Targets Underlying the Anticancer Effects of Quercetin: An Update Nutrients 8(9):529. |
|
|
Kleemann R, Verschuren L, Morrison M, Zadelaar S, van EJ, Wielinga, Y, Kooistra T (2011). Anti-inflammatory, anti-proliferative and antiatherosclerotic effects of quercetin in human in vitro and in vivo models. Atherosclerosis 218:44-52. |
|
|
Kohgo Y, Ikuta K, Ohtake T, Torimoto Y, Kato J (2008). Body iron metabolism and pathophysiology of iron overload. International Journal of Hematology 88(1):7-15. |
|
|
Koracevic D, Koracevic G (2001). Total antioxidant capacity. Journal of Clinical Pathology 54:356-361. |
|
|
Kozlowski H, Janicka-Klos A, Brasun J, Gaggelli E (2009). Copper, iron, and zinc ions homeostasis and their role in neurodegenerative disorders (metal uptake, transport, distribution and regulation). Coordination Chemistry Reviews 253:2665-2685. |
|
|
Kremastinos T, Farmakis D, Aessopos A, Hahalis G, Hamodraka E, Tsiapras D (2010). Betathalassemia cardiomyopathy: history, present considerations, and future perspectives. Circulation Heart Failure 3(3):451-458. |
|
|
Kulaksiz H, Fein E, Redecker P, Stremmel W, Adler G, Cetin Y (2008). Pancreatic b-cells express hepcidin, an iron-uptake regulatory peptide. Journal of Endocrinology 197:241-249. |
|
|
Lebda M (2014). Acute Iron Overload and Potential Chemotherapeutic Effect of Turmeric in Rats. International Journal of Pure and Applied Bioscience 2(2):86-94. |
|
|
Leopoldini M, Sandro C, Marisosa T (2006). Iron chelation by the powerful antioxidant flavonoid quercetin. Journal of Agricultural and Food Chemistry 54:6343-6351. |
|
|
Li Y, Hansen L, Borst B, Spears W, Moeser J (2016a). "Dietary iron deficiency and over supplementation increase intestinal permeability, ion transport, and inflammation in pigs," Journal of Nutrition 146(8):1499-1505. |
|
|
Li Y, Yao J, Han Ch, Yang J, Chaudhry T, Wang S, Liu H, Yin Y, (2016b). Quercetin, Inflammation and Immunity. Nutritional 8:167 |
|
|
Madhusudhan S, Oberoi R (2011). Renal iron deposition in aplastic ane-mia: magnetic resonance imaging appearance. Indian Journal of Nephrology 21:134-135. |
|
|
Matos R, Capelozzi L, Gomes F, Mascio D, Medeiros H (2001). Lycopene inhibits DNA damage and liver necrosis in rats treated with ferric nitrilo tri acetate. Archives of Biochemistry and Biophysics 396:171-177. |
|
|
Nijveldt J, van Nood E, van Hoorn C, Boelens G, van Norren K, van Leeuwen M (2001)."Flavonoids: a review of probable mechanisms of action and potential applications," American Journal of Clinical Nutrition 74(4):418-425. |
|
|
Ohkawa H, Ohishi W, Anal K (1979). Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Analytical Biochemistry 95:351-358. |
|
|
Nunez T, Urrutia P, Mena N, Aguirre P, Tapia V, Salazar J (2012): "Iron toxicity in neurodegeneration," Biometals 25(4):761-776. |
|
|
Pari L, Asaithambi K, Paramasivam K, Ayyasamy R (2015). Protective effects of hesperidin on oxidative stress, dyslipidaemia and histological changes in iron-induced hepatic and renal toxicity in rats. Toxicology Reports 2:46-55. |
|
|
Porter B (1997). A risk-benefit assessment of iron-chelation therapy. Drug Safety 17:407-21. |
|
|
Qi L, Cao C, Hu L, Chen S, Zhao X, Sun C (2017). Metabonomic analysis of the protective effect of quercetin on the toxicity induced by mixture of organophosphate pesticides in rat urine:Human and Experimental Toxicology 36(5):494-507. |
|
|
Raza A, Xu X, Xia L, Xia C, Tang J, Ouyang Z (2016). Quercetin-Iron Complex: Synthesis, Characterization, Antioxidant, DNA Binding, DNA Cleavage, and Antibacterial Activity Studies: Journal of Fluorescence 26:2023-2031. |
|
|
Singh P, McCoy T, Tice R, Schneider L (1988). A simple technique for quantitation of low levels of DNA damage in individual cells. Experimental Cell Research 175(1):184-191. |
|
|
Sinha R, Srivastava S, Joshi A, Joshi J, Govil G (2014). In-vitro anti-proliferative and anti-oxidant activity of galangin, fisetin and quercetin: role of localization and intermolecular interaction in model membrane. European Journal of Medicinal Chemistry 79: 102-109. |
|
|
Stookey LL (1970). "Ferrozine-A New Spectrophotometric Reagent for Iron," Analytical Chemistry 42(7):779-781. |
|
|
Tice R, Agurell E, Anderson D, Burlinson B, Hartmann A, Kobayashi H, Miyamae E, Ryu C, Sasaki F (2000). Single cell gel/ cometassay: guidelines for in vitro and in vivo genetic toxicology testing. Environmental and Molecular Mutagenesis 35:206-221. |
|
|
Torreggiani A, Tamba M, Trinchero A, Bonora S (2005). Copper (II)–Quercetin complexes in aqueous solutions: spectroscopic and kinetic properties. Journal of Molecular Structure 744:759-766. |
|
|
Torti V, Torti M (2013). Iron and cancer:more ore to be mined. Nature Reviews Cancer 13: 342-355. |
|
|
Uranga M, Salvador A (2018). Unraveling the Burden of Iron in Neurodegeneration: Intersections with Amyloid Beta Peptide Pathology. Oxidative Medicine and Cellular Longevity, Article ID 2850341, 12. |
|
|
Urrutia P, Aguirre A, Esparza A, Tapia V (2013). Inflammation alters the expression of DMT1, FPN1 and hepcidin and causes iron accumulation in central nervous system cells. International Society for Neurochemistry. Journal of Neurochemistry 126:541-549. |
|
|
Uzun G, Demir F, Kalender S (2010). Protective effect of catechin and quercetin on chlorpyrifos-induced lung toxicity in male rats. Food and Chemical Toxicology 48:1714-1720. |
|
|
Uzun G, Kalender Y (2013). Chlorpyrifos induced hepatotoxic and hematologic changes in rats: the role of quercetin and catechin. Food and Chemical Toxicology 55:549-556. |
|
|
Valko M, Rhodes J, Moncol J, Izakovic M, Mazur M (2006). Free radicals, metals and antioxidants in oxidative stress-induced cancer. Chemico- Biological Interactions 160:1-40. |
|
|
Valko M, Morris H, Cronin T (2005). Metals, toxicity and oxidative stress. Current Medicinal Chemistry 12:1161-1208. |
|
|
Von Bernhardi R, Eugenin J (2012). Alzheimer's disease: redox dysregulation as a common denominator for diverse pathogenic mechanisms. Antioxidants and Redox Signaling 16:974-1031. |
|
|
Yang J, Cao J, Sun X, Feng Z, Hao D, Zhao X, Sun C (2012). Effects of long-term exposure to low levels of organophosphorous pesticides and their mixture on altered antioxidative defense mechanisms and lipid peroxidation in rat liver. Cell Biochemistry and Function 30:122-128. |
|
|
Zantopoulos V, Antoniou V, Pet saga V, Zdrags A (1996). "Copper concentration in sheep liver and kidney in Greece," Veterinary and Human Toxicology 38(3):184-185. |
|
|
Zhang Y, Li H, Zhao Y, Gao Z (2006). "Dietary supplementation of baicalin and quercetin attenuates iron overload induced mouse liver injury," European Journal of Pharmacology 535(30):263-269. |
|
|
Zhang Y, Gao Z, Liu J, Xu Z (2011). Protective effects of baicalin and quercetin on an iron-overloaded mouse: comparison of liver, kidney and heart tissues. Natural Product Research 25(12):1150-1160. |
|
|
Zivadinov R, Weinstock-Guttman B, Pirko I (2011). Iron deposition and inflammation in multiple sclerosis. Which one comes first? BMC Neuroscience 12:60. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


