ABSTRACT
The application of bacteriophages biocontrol requires the formulation of genetically distinct bacteriophages in a phage cocktail. Random Amplified Polymorphic DNA (RAPD) - PCR is considered a cheap, reproducible, and readily applicable tool in detecting phage diversity compared to other molecular techniques such as whole-genome sequencing. We used in this study the RAPD-PCR technique to assess the genetic diversity of 28 bacteriophages infecting Pseudomonas aeruginosa and Staphylococcus aureus. According to their RAPD profiles, isolated phages were grouped into 2 main clusters which included phages from the same host. The typing by RAPD-PCR of newly isolated phages was useful to assess the genetic diversity bypassing previous whole-genome sequencing analysis. These genetically distinct phages lytic against P. aeruginosa and S. aureus could potentially be used in a phage cocktail for biocontrol against these clinically and industrially relevant bacteria.
Key words: Phages, genetic diversity, RAPD PCR, P. aeruginosa, S. aureus.
Bacteriophages (phages), the viruses of bacteria are the most abundant biological entities in the biosphere and are always associated with their host bacteria in the same ecological niche. Consequently, phages have an impact on the bacterial ecosystems both locally in our immediate communities and globally (Abedon, 2008; de Leeuw et al., 2020; Harada et al., 2018). Generally, both Pseudomonas aeruginosa and Staphylococcus aureus are virulent pathogenic organisms for many clinical conditions. P. aeruginosa is one of the major life-threatening opportunistic bacteria responsible for nosocomial infections in immunocom promised people, a leading cause of lung infections and ventilator-associated pneumonia (Essoh et al., 2015; Wang et al., 2017), while S. aureus is responsible for skin and soft tissues infections and invasive diseases such as endocarditis, osteomyelitis (Wang et al., 2016). Furthermore, both species are notorious for their resistance to antimicrobials. P. aeruginosa and S. aureus are members of the ESKAPE pathogens which are known for widespread resistance to almost all classes of antimicrobials used in a hospital setting (Santajit and Indrawattana, 2016). Due to the importance of these infections worldwide, phages’ activity against these pathogens were considered as an eco-friendly alternative to antibiotics (Abatángelo et al., 2017; Essoh et al., 2013; Kifelew et al., 2020; Oliveira et al., 2015; Song et al., 2021). Phages with lytic activity against these bacterial pathogens are sourced from the environment or clinical specimens linked to diseases caused by P. aeruginosa and S. aureus.
Bacteriophages have a narrow host range as a single phage may infect only specific strains. However, some phages are able to infect various bacterial species (Hamdi et al., 2017). The narrow host range of phages has not limited their use in industries and therapeutically. This nature and the ability of bacteria to evolve to resist a lytic phage, therefore, necessitate the use of phage cocktails in biocontrol. However, one of the best methods for the assessment of genetic diversity is whole-genome sequencing (WGS) and analysis. These techniques are expensive and not always available in some countries. Thus, a reproducible and affordable approach would be very valuable to readily assess diversity among newly isolated phages. Random amplified polymorphic DNA (RAPD)-PCR has been used to discriminate among different phage lineage and for assessment of their genotypic diversity in terms of hours (Ács et al., 2020).
In this study, we investigate lytic phages against P. aeruginosa and S. aureus specific-phages. The phage isolation was performed with samples collected from urban sewage and Ebrié Lagoon of Abidjan, Côte d’Ivoire, given that sewage and environmental water are optimal sources of phages (Alharbi and Ziadi, 2021; Weber-D?browska et al., 2016). The RAPD PCR of isolated phage DNA extracts helped to discriminate between the phages and to assess their genetic diversity.
Bacterial strains and growth conditions
Two bacterial strains isolated in 2017 were used for the phage isolation. P. aeruginosa strain “P02” was isolated in sewage sample and S. aureus strain “S04” in street vendor hands. Both strains were provided by Plateforme de biologie moléculaire of Institut Pasteur de Côte d’Ivoire. The strains were grown overnight in Lysogeny Broth (LB) (10 g.l-1 NaCl, 10 g.l-1 Tryptone, 5 g.l-1 Yeast Extract) and stored in 200 µl aliquots in medium with 30% glycerol at -20°C. A hundred microliters from stored bacteria were added to 3 ml of LB and grown overnight at 37°C for isolation purposes.
Phage isolation and purification
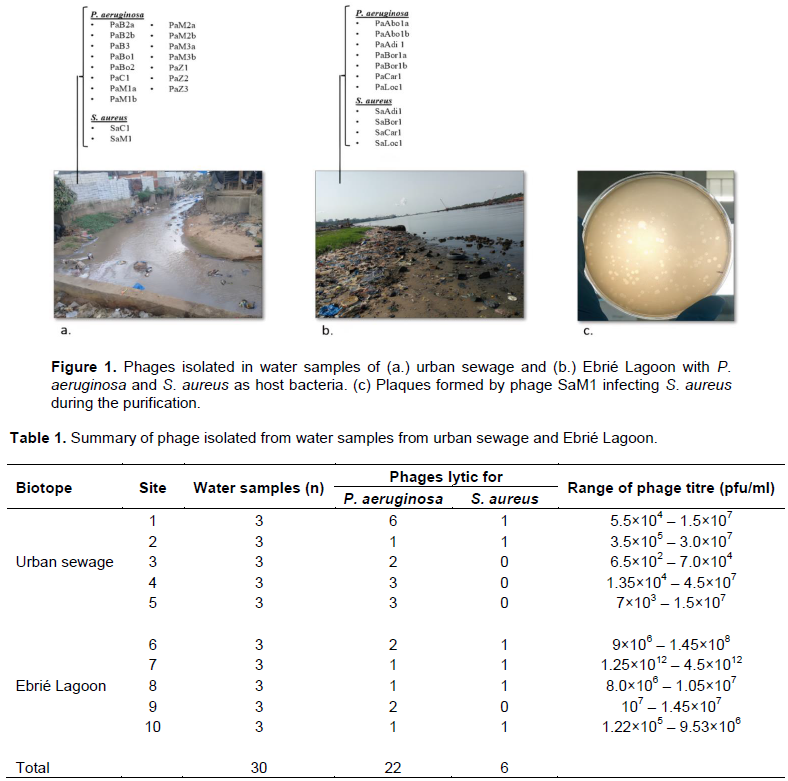
Fifteen samples of urban sewage were collected from wastewater collectors of the municipality of Yopougon, Abidjan. Fifteen other samples were collected in the Ebrié Lagoon of Abidjan. Samples were collected using sterile 500-ml glass vials, connected to a rope. Biosafety rules were applied for the protection of the sampler and the samples (Personal protective equipment). Samples were taken instantaneously by immersing the vial in the effluent to be sampled. The opening of the vial was placed facing the effluent stream, under the surface of the liquid, and a 300 ml sample was taken. A space of 2.5 cm is left between the level of the sample and the lid of the bottle. The vial is sealed and kept in a container at 4°C for transport to the laboratory.
The samples were then enriched to detect P. aeruginosa and S. aureus lytic phages according to a modification of (Van Twest and Kropinski, 2009)procedure. Briefly, samples were centrifuged at 12000 g for 10 min and filtered through 0.22 µm pore membranes. Ten milliliters of filtered water were added to 10 ml of double-strength LB broth (10 g.l-1 NaCl, 10 g.l-1 Tryptone and 5 g.l-1 Yeast Extract) with 100 µl of the enrichment bacterial strain. The mixture was incubated at 37°C, 48 h. Thereafter, the culture was centrifuged at 12000 g, 5 min, and the supernatant was filtered and evaluated for the presence of lytic phages using the double-layer agar method. Serial decimal dilution up to 10-8 of filtered lysate was carried out. Then, 100 µl of overnight bacterial culture (18 – 24 h) was mixed with 0.6% soft LB agar and poured on a 1.5% bottom LB agar plate. When the top agar layer was set, serial dilutions of filtered lysate were spotted onto the overlay, plates were incubated overnight at 37°C and observed for isolated plaques. Phage plaques were purified three times to ensure phage purity (Salem et al., 2015). A sterile truncated tip was stabbed through a single well-isolated plaque that was immediately added to 500 µl of SM buffer (50 mM Tris-HCl pH7.5, 100 mM NaCl, 8.1 mM MgSO4), mixed and refrigerated for 1 h at 4°C. After incubation, the mixture was centrifuged and 300 µl of supernatant was filtered and transferred to a new tube. Purified phages were preserved at 4°C before genomic DNA extraction and at -20°C for long-term conservation.
Genomic fingerprinting by RAPD analysis
Phage DNA was extracted using DNeasy Blood and Tissue Kit (Qiagen, Hilden, Germany) as described previously (Jako?i?n? and Moodley, 2018)from 450 µL of phage stocks with a titer between 109 to 1014 plaque-forming unit (pfu)/ml. RAPD-PCR was carried out according to a modification of the method described previously (Kumari et al., 2009). Primers P1 (5’-CCGCAGCCAA-3’) and P2 (5’-AACGGGCAGA-3’) were used as designed in the study of Gutiérrez et al. (2011). PCR mixture (50 μl) consisted of 1 μl of phage DNA, 5.0 μl 5X Green Reaction Buffer, 5.0 µl 5X Green Reaction Buffer, 3.0 µl MgCl2 (25 mM), 1.0 μl dNTP’s (25 mM) (Promega, Wisconsin, USA), 1 μl primer, 0.5 μl GoTaq G2 Flexi DNA polymerase (5 U/µl) (Promega, Wisconsin, USA) and 29.5 μl DNase/RNase free water (Promega, Wisconsin, USA). Reactions were performed at 95°C for 5 min followed by 45 cycles of consecutive primers annealing (26 and 31°C for 3 min), extension (72°C for 2 min), denaturation (94°C for 1 min), and final extension (72°C for 10 min).
DNA band patterns were obtained after gel electrophoresis on 2% agarose gel of the RAPD-PCR reaction products (15 µL). Gels were run for 120 min at 80 V using a 1kb DNA ladder (Promega) as a molecular weight marker. The gel images were recorded using the Gel Doc EZ Gel Documentation System (BioRad). The RAPD profile was analyzed using the software GelJ v2.0 (Heras et al., 2015). The similarity matrix was calculated based on the Pearson correlation coefficient, and its corresponding dendrogram was deduced using the complete linkage clustering method in GelJ.
Phage collection
In this study, 28 lytic phages were isolated, 17 in urban sewage samples and 11 in Ebrie lagoon water samples (Figure 1). P. aeruginosa is the host of 75% of isolated phages, compared to 25% for S. aureus. Site 1 provided the largest number of P. aeruginosa specific-phages. This latter could be isolated at least once in all studied sites. On the contrary, S. aureus-specific-phages could not be recovered in four sites (Table 1).
Phage genetic diversity analysis
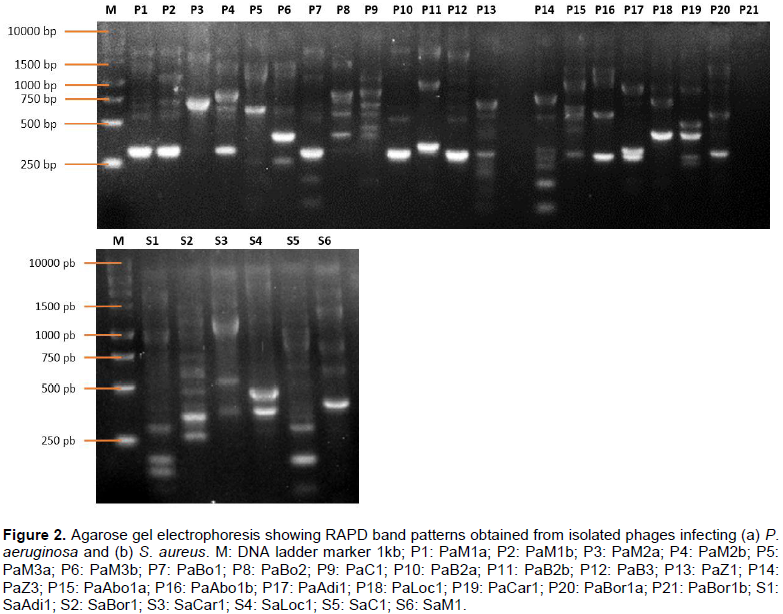
RAPD-PCR on phage DNA constitutes a suitable approach to quickly assess the genetic diversity among newly isolated bacteriophages infecting the same species. Two different primers were used, out of which only P2 (5’-AACGGGCAGA-3’) provided bands with almost all the phages, except for phages PaZ1 and PaBor1b both infecting P. aeruginosa. No profile was observed with primer P1. The genomic fingerprints from the phages infecting the same host were likely distinct and have shown different numbers of bands ranging from 2 to 8 respectively for PaM2a and PaC1 infecting P. aeruginosa, and from 3 to 10 respectively for SaLoc1 and SaC1 infecting S. aureus. Also, the fingerprinting differed by the bands' intensity, and the fragment size varied ranging from less than 250 bp to more than 10.000 bp (Figure 2).
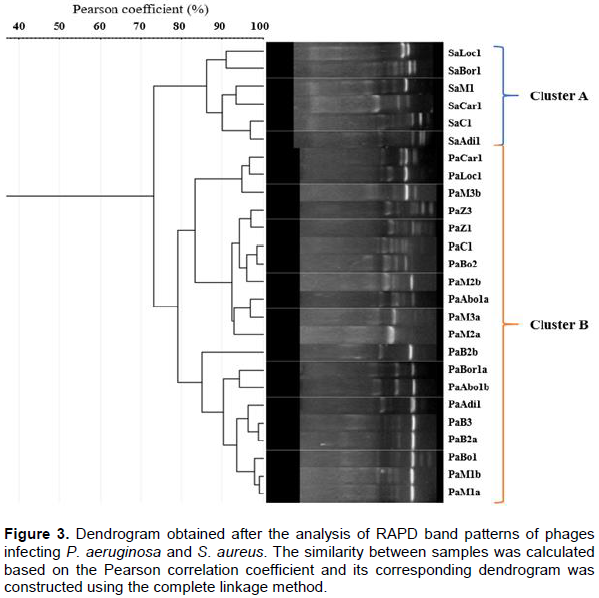
However, phages PaM1a, PaM1b, PaBo1, PaB2a, and PaB3 are closely related with a similarity of Pearson greater than 98% (Figure 3). They are all isolated in urban sewage samples. Unsurprisingly, the dendrogram revealed 2 main clusters: cluster A including all phages infecting S. aureus, and cluster B including phages infecting P. aeruginosa. Similarity among phages infecting S. aureus is more than 86%, while similarity among P. aeruginosa phages is 78%.
In this study, water samples from urban sewage of Yopougon and Ebrie Lagoon in Abidjan, Côte d’Ivoire were screened for the presence of P. aeruginosa and S. aureus specific lytic phages. Bacteriophages infecting P. aeruginosa were easily isolated from sewage as reported by previous studies conducted in urban sewage of the municipality of Cocody in Abidjan (Essoh et al., 2015). However, there are no previous documented reports of S. aureus-specific phages isolated in sewage and environmental water in Côte d’Ivoire. We noticed also that isolated phages are unevenly distributed regarding the sampling ecosystem and the isolation strain. Phages are known to be found where their host is present (Son et al., 2012). This collection has shown a high rate of phages from urban sewage infecting P. aeruginosa. Based on the “kill the winner” theory, it can be suggested that these strains are the most abundant and common in the studied ecosystems. This theory assumes that phages adapt to infect preferentially lineage of bacteria with the highest frequencies in the population (Koskella and Meaden, 2013). Indeed, studies performed in Côte d’Ivoire revealed a high prevalence of these bacteria in urban wastewater (Coulibaly-Kalpy et al., 2016; Guessennd et al., 2013). These latter are directly discharged without prior treatment within the Ebrié Lagoon (Dongo et al., 2013; Scheren et al., 2004). Concerning S. aureus infecting phages, we noticed a low rate of isolation, especially from sewage. While, urban sewages and environmental water are known to be a source of phages infecting P. aeruginosa (Azizian et al., 2015; Mattila et al., 2015; Weber-D?browska et al., 2016), S. aureus’ phages isolation seems to be laborious. An evaluation of on-demand isolation of phages revealed that the probability for discovering a phage from a sewage sample against the most common hospital pathogens was 6.1% for S. aureus and 79.4% for P. aeruginosa (Mattila et al., 2015).
The genetic diversity revealed distinct RAPD profiles showing that the phages were unique and genetically diverse. The RAPD PCR technique allowed us to discriminate between phages lytic against two different bacterial species and genus. As seen in the study of Guriérrez et al. (2011), RAPD PCR is a great approach for typing phages infecting strains belonging to the same species or different species within the same genus or a different genus. This technique was used in previous studies to characterize newly isolated phages against enteropathogenic, enterohemorrhagic, enterotoxigenic, and Shiga-toxin-producing strains of E. coli (Dini and De Urraza, 2010; Kakou-Ngazoa et al., 2020). It was also used for typing phages infecting P. aeruginosa (Azizian et al., 2015; Kumari et al., 2009; Li et al., 2010).
The dendrogram was deduced using the complete linkage clustering method. Usually, molecular typing dendrograms in ecology are constructed based on average similarities among objects or on centroids of clusters (Gutiérrez et al., 2011). By contrast, complete linkage clustering demands, for a group to agglomerate with another group, that all objects be related at the given similarity. The result clustering offers a much more robust and contrasting analysis. The clustering obtained with the dendrogram in this study was not surprising because of the phylogenetic relations among phages. Phages isolated with the same species were clustered together. As previously described, phages infecting a specific bacterial host are most similarly and closely related (Hatfull, 2008). This technique was very useful in assessing the diversity of phages isolated in both urban sewage and the Ebrié Lagoon.
Twenty-eight genetically distinct phages lytic against P. aeruginosa and S. aureus were isolated from water samples in this study. RAPD PCR was useful in performing the comparison between viral strains and did not require earlier genome characterization. These phages and subsequent cocktails could be useful for biocontrol against clinically and industrially relevant bacteria.
The authors have not declared any conflict of interests.
Ms. Addablah was financially supported by a scholarship from the Ministry of Higher Education and Scientific Research, Lomé, Togo.
REFERENCES
|
Abatángelo V, Peressutti BN, Boncompain CA, Amadio AA, Carrasco S, Suárez CA, Morbidoni HR (2017). Broad-range lytic bacteriophages that kill Staphylococcus aureus local field strains. PLOS ONE 12(7):e0181671.
Crossref
|
|
|
|
Abedon ST (2008). Bacteriophage ecology: Population growth, evolution, and impact of bacterial viruses. Cambridge University Press P 15.
Crossref
|
|
|
|
|
Ács N, Gambino M, Brøndsted L (2020). Bacteriophage Enumeration and Detection Methods. Frontiers in Microbiology 11:2662.
Crossref
|
|
|
|
|
Alharbi NM, Ziadi MM (2021). Wastewater as a fertility source for novel bacteriophages against multi-drug resistant bacteria. Saudi Journal of Biological Sciences, 28(8):4358?4364.
Crossref
|
|
|
|
|
Azizian R, Nasser A, Askari H, Taheri Kalani M, Sadeghifard N, Pakzad I, Amini R, Mozaffari NAS, Azizi Jalilian F (2015). Sewage as a rich source of phage study against Pseudomonas aeruginosa PAO. Biologicals 43(4):238-241.
Crossref
|
|
|
|
|
Coulibaly-Kalpy J, Kouamé YC-K, Ouattara Koffi N, Kouamé K, Amon Lydie N, Ehuié P, Yéo K, Bamba A, Gourène G, Dosso-Bretin M (2016). Potential Pathogenic Bacteria of Wastewater Collectors from Abidjan (Côte d'Ivoire). International Journal of Current Microbiology and Applied Sciences 5(5):358-369.
Crossref
|
|
|
|
|
De Leeuw M, Baron M, Ben DO, Kushmaro A (2020). Molecular Insights into Bacteriophage Evolution toward Its Host. Viruses 12(10):1132.
Crossref
|
|
|
|
|
Dini C, De Urraza PJ (2010). Isolation and selection of coliphages as potential biocontrol agents of enterohemorrhagic and Shiga toxin-producing E. coli (EHEC and STEC) in cattle?: Selection of coliphages as biocontrol agents. Journal of Applied Microbiology 109(3):873-887.
Crossref
|
|
|
|
|
Dongo K, Niamke B, Adje A, Britton B, Nama L, Anoh K, Adima A, Atta K (2013). Impacts des effluents liquides industriels sur l'environnement urbain d'Abidjan-Côte D'Ivoire. International Journal of Biological and Chemical Sciences 7(1):404.
Crossref
|
|
|
|
|
Essoh C, Blouin Y, Loukou G, Cablanmian A, Lathro S, Kutter E, Thien HV, Vergnaud G, Pourcel C (2013). The Susceptibility of Pseudomonas aeruginosa Strains from Cystic Fibrosis Patients to Bacteriophages. PLoS One 8(4):e60575.
Crossref
|
|
|
|
|
Essoh C, Latino L, Midoux C, Blouin Y, Loukou G, Nguetta S-PA, Lathro S, Cablanmian A, Kouassi AK, Vergnaud G, Pourcel C (2015). Investigation of a Large Collection of Pseudomonas aeruginosa Bacteriophages Collected from a Single Environmental Source in Abidjan, Côte d'Ivoire. Plos One 10(6):e0130548.
Crossref
|
|
|
|
|
Guessennd N, Ouattara M, Ouattara N, Nevry R, Gbanon V, Tiekoura K, Dosso M, Ger B (2013). Étude des bactéries multirésistantes des effluents hospitaliers d'un centre hospitalier et universitaire (CHU) de la ville d'Abidjan (Côte d'Ivoire). Journal of Applied Biosciences, 69(0):5456.
Crossref
|
|
|
|
|
Gutiérrez D, Martín-Platero AM, Rodríguez A, Martínez-Bueno M, García P, Martínez B (2011). Typing of bacteriophages by randomly amplified polymorphic DNA (RAPD)-PCR to assess genetic diversity?: Bacteriophage typing by RAPD-PCR. FEMS Microbiology Letters 322(1):90?97.
Crossref
|
|
|
|
|
Hamdi S, Rousseau GM, Labrie SJ, Tremblay DM, Kourda RS, Ben Slama K, Moineau S (2017). Characterization of two polyvalent phages infecting Enterobacteriaceae. Scientific Reports 7(1):40349.
Crossref
|
|
|
|
|
Harada LK, Silva EC, Campos WF, Del Fiol FS, Vila M, D?browska K, Krylov VN, Balcão VM (2018). Biotechnological applications of bacteriophages?: State of the art. Microbiological Research 212(213):38?58.
Crossref
|
|
|
|
|
Hatfull GF (2008). Bacteriophage genomics. Current Opinion in Microbiology 11(5):447-453.
Crossref
|
|
|
|
|
Heras J, Domínguez C, Mata E, Pascual V, Lozano C, Torres C, Zarazaga M (2015). GelJ - a tool for analyzing DNA fingerprint gel images. BMC Bioinformatics 16(1):270.
Crossref
|
|
|
|
|
Jako?i?n? D, Moodley A (2018). A Rapid Bacteriophage DNA Extraction Method. Methods and Protocols 1(3):27.
Crossref
|
|
|
|
|
Kakou-Ngazoa ES, Addablah AA, Krylova K, Saraka D, Kouassi KS, Coulibaly ND, Sina KM, Aoussi S, Dozois C, Dosso M (2020). First novel phages from rodents with lytic activity on clinical Enterobacteriaceae strains?: Initiation for phage therapy in West Africa. African Journal of Microbiology Research 14(6):280-285.
Crossref
|
|
|
|
|
Kifelew LG, Warner MS, Morales S, Thomas N, Gordon DL, Mitchell JG, Speck PG (2020). Efficacy of Lytic Phage Cocktails on Staphylococcus aureus and Pseudomonas aeruginosa in Mixed-Species Planktonic Cultures and Biofilms. Viruses 12(5):559.
Crossref
|
|
|
|
|
Koskella B, Meaden S (2013). Understanding Bacteriophage Specificity in Natural Microbial Communities. Viruses 5(3):806-823.
Crossref
|
|
|
|
|
Kumari S, Harjai K, Chhibber S (2009). Characterization of Pseudomonas aeruginosa PAO Specific Bacteriophages Isolated from Sewage Samples. American Journal of Biomedical Sciences 1(2):91-102.
Crossref
|
|
|
|
|
Li L, Yang H, Lin S, Jia S (2010). Classification of 17 newly isolated virulent bacteriophages of Pseudomonas aeruginosa. Canadian Journal of Microbiology 56(11):925-933.
Crossref
|
|
|
|
|
Mattila S, Ruotsalainen P, Jalasvuori M (2015). On-Demand Isolation of Bacteriophages against Drug-Resistant Bacteria for Personalized Phage Therapy. Frontiers in Microbiology 6:1271.
Crossref
|
|
|
|
|
Oliveira H, Sillankorva S, Merabishvili M, Kluskens LD, Azeredo J (2015). Unexploited opportunities for phage therapy. Frontiers in Pharmacology 6:180.
Crossref
|
|
|
|
|
Salem M, Virtanen S, Korkeala H, Skurnik M (2015). Isolation and characterization of Yersinia- specific bacteriophages from pig stools in Finland. Journal of Applied Microbiology 118(3):599-608.
Crossref
|
|
|
|
|
Santajit S, Indrawattana N (2016). Mechanisms of Antimicrobial Resistance in ESKAPE Pathogens. BioMed Research International pp. 1-8.
Crossref
|
|
|
|
|
Scheren PAGM, Kroeze C, Janssen FJJG, Hordijk L, Ptasinski KJ (2004). Integrated water pollution assessment of the Ebrié Lagoon, Ivory Coast, West Africa. Journal of Marine Systems 44(1?2):1-17.
Crossref
|
|
|
|
|
Son B, Yun J, Lim J-A, Shin H, Heu S, Ryu S (2012). Characterization of LysB4, an endolysin from the Bacillus cereus-infecting bacteriophage B4. BMC Microbiology 12(1):33.
Crossref
|
|
|
|
|
Song J, Ruan H, Chen L, Jin Y, Zheng J, Wu R, Sun D (2021). Potential of bacteriophages as disinfectants to control of Staphylococcus aureus biofilms. BMC Microbiology 21(1):57.
Crossref
|
|
|
|
|
Van Twest R, Kropinski AM (2009). Bacteriophage enrichment from water and soil. In: J. M. Walker, Bacteriophages?: Methods and protocols, Volume 1?: Isolation, characterization, and interactions (Vol. 501). Humana Press.
Crossref
|
|
|
|
|
Wang K, Chen Y, Salido MM, Kohli GS, Kong J, Liang H, Yao Z, Xie Y, Wu H, Cai S, Drautz-Moses DI, Darling AE, Schuster SC, Yang L, Ding Y (2017). The rapid in vivo evolution of Pseudomonas aeruginosa in ventilator-associated pneumonia patients leads to attenuated virulence. Open Biology 7(9):170029.
Crossref
|
|
|
|
|
Wang Z, Zheng P, Ji W, Fu Q, Wang H, Yan Y, Sun J (2016). SLPW?: A Virulent Bacteriophage Targeting Methicillin-Resistant Staphylococcus aureus In vitro and In vivo. Frontiers in Microbiology 7:934.
Crossref
|
|
|
|
|
Weber-D?browska B, Jo?czyk-Matysiak E, ?aczek M, ?obocka M, ?usiak-Szelachowska M, Górski A (2016). Bacteriophage Procurement for Therapeutic Purposes. Frontiers in Microbiology 7:1177.
Crossref
|
|