Full Length Research Paper

ABSTRACT
Among the enzymes of the cellulolytic complex, β-glucosidases are noteworthy due to the possibility of their application in different industrial processes, such as production of biofuels, winemaking, and development of functional foods. This study aimed to evaluate the production and characterization of β-glucosidase from the filamentous fungus Gongronella butleri, recently isolated from Cerrado soil and cultivated in agro-industrial residue substrates. The highest production of β-glucosidase, about 215.4 U/g of dry substrate (or 21.5 U/mL), was obtained by cultivation of the microorganism on wheat bran with 55% of the initial moisture, for 96 h at 30°C. This β-glucosidase showed higher catalytic activity at pH 4.5, and a temperature of 65°C. The original enzymatic activity was recovered in a pH range of 3.0-7.5 after 24 h of incubation. The enzyme retained 80% of its catalytic activity when incubated for 1 h at 50°C. The enzyme was strongly inhibited by glucose, an effect that was completely reversed by increasing substrate concentration in the reaction mixture, which is typical for competitive inhibition. High catalytic activity was observed in solutions containing up to 20% ethanol, allowing the application of this enzyme in processes with high alcohol concentrations (for example beverages and biofuels). The significant production of β-glucosidase by the selected strain, along with these enzyme characteristics, highlights the biotechnological potential of the fungus G. butleri.
Key words: Microbial enzyme, biofuels, agro-industrial residues, cellulases, hemicellulases.
INTRODUCTION
Agro-industrial residues are usually discarded in the environment, yet their composition allows the use of such materials as nutrient sources in fermentation processes. In this context, solid-state fermentation is essential in the use of residues as substrates for the synthesis of various compounds of industrial interest, in addition to being beneficial to the environment (Deswal et al., 2011; Pereira et al., 2015a).
Lignocellulosic biomass is the most abundant renewable source found in nature, and is mainly composed of cellulose, hemicellulose and lignin (Pauly and Keegstra, 2010). Cellulose is a linear homopolysaccharide that constitutes of glucose molecules linked through β-glycosidic bonds of the type (1 → 4) (Huber et al., 2006; Thakur and Thakur, 2014). For efficient conversion of cellulose into fermentable sugars, an enzymatic system constituted of at least three groups of enzymes is necessary: (1) endoglucanases (EC 3.2.1.4), which internally hydrolyze cellulose chains, resulting in a rapid reduction in the degree of polymerization; (2) exoglucanases (EC 3.2.1.91), which hydrolyze glycosidic bonds at the ends of chains, releasing principally cellobiose; and (3) β-glucosidases (EC 3.2.1.21), which finalize hydrolysis, converting cellobiose to glucose. The glucose from lignocellulosic biomass can be converted into ethanol through fermentation by microorganisms, which has aroused great interest in biofuel industries (Bansal et al., 2012).
β-Glucosidase has the ability to increase the overall yield of fermentable sugars (Rani et al., 2014) while reducing the cellobiose inhibitory effect on other cellulolytic enzymes, promoting continuity of enzymatic hydrolysis process (Rani et al., 2014; Borges et al., 2014). These enzymes have many applications in industrial processes, including the conversion of isoflavone glycosides into aglycones, which have antioxidant activities and are more easily absorbed by the human intestine. Studies have shown that the use of these aglycones confer health benefits, including the prevention of certain types of cancer and a reduction in the risk of cardiovascular disease, osteoporosis, menopausal symptoms and diabetes, among others (Leite et al., 2007; Singhania et al., 2013). The addition of microbial β-glucosidase in winemaking processes promotes the release of volatile terpenes (deglycosylated by the action of the enzyme), contributing to the aromatic composition of the wine (Alves-Prado et al., 2011).
The high cost of enzymes is a major factor limiting the use of these biocatalysts on a large scale. Increasing productivity while reducing the cost of production are crucial factors for the industrial application of an enzyme (Romero et al., 2007; Oliveira et al., 2015). In this context, this study aimed to optimize the parameters in solid-state fermentation of the filamentous fungus Gongronella butleri, aimed at producing β-glucosidase. The biochemical characteristics of β-glucosidase and the catalytic profile of enzymatic extract produced were also assessed.
MATERIALS AND METHODS
Microorganism
In this study, the filamentous fungus mesophilic Gongronella butleri was used. The fungus was isolated from soil samples collected from the Cerrado biome, located in Dourados-MS (Brazilian Midwest-22°10’49.2’’S 54°56’57.4W). Soil samples were serially diluted (10-1-10-4) in a solution of sodium chloride (0.9%) and plated on Sabouraud Dextrose Agar (dextrose 4%, mycological peptone 1% and agar 1.5%), aiming to obtain purified strains. The selected strain was identified by Brazilian Collection of Microorganisms of Industry and Environment (Coleção Brasileira de Microrganismos de Indústria e Meio Ambiente – CBMAI), at UNICAMP, Campinas, SP. The fungus was cultivated at 28°C in Sabouraud Dextrose Agar and maintained at 4°C.
β-Glucosidase production by solid-state fermentation
Inoculum
The fungus was cultivated in 250 mL Erlenmeyer flasks containing 40 mL of Sabouraud Dextrose Agar, and incubated for 48 h at 28°C. Microorganism suspension was obtained by scraping the surface of the culture medium using 25 mL of nutrient solution (0.1% ammonium sulfate, 0.1% magnesium sulfate heptahydrate and 0.1% ammonium nitrate, w/v) (Merheb-Dini et al., 2009). The fungus was inoculated in the agro-industrial residues by transfer of 5 mL of the microbial suspension (105 spores/g of dry substrate). The inoculum volume was considered to calculate the initial moisture in solid-state fermentation processes.
Solid-state fermentation
Several agro-industrial residues were tested for β-glucosidase production: wheat bran, soy bran, rice peel, corn cobs and corn straw. Prior to inoculation with microorganism, all substrates were washed with distilled water and dried at 50°C for 48 h. Fermentation was performed in 250 mL Erlenmeyer flasks containing 5 g of substrate sterilized at 121°C for 20 min. Initially, all substrates were moistened to 60% with nutrient solution (previously described). After inoculation of the microorganism, Erlenmeyer flasks were incubated at 28°C for 96 h. The substrate that showed the best β-glucosidase production by the fungus was selected for the evaluation of other fermentation parameters (e.g., moisture, temperature and time of cultivation) whose optimized values were adopted in subsequent assays. All the assays were performed in duplicate. The results were expressed as U/g refer to enzymes units per gram of dry substrate.
Enzyme extraction
Extraction of the enzyme from the fermented substrate was carried out by adding 50 mL of distilled water, and then shaking constantly at 150 rpm for 1 h. This mixture was then filtered through synthetic tissue (nylon) and centrifuged at 1500 ×g for 5 min at 5°C. The supernatant was used for the enzymatic assays.
Determination of β-glucosidase activity
β-Glucosidase activity was determined with 50 µL of enzymatic extract, 250 µL of 0.1 M sodium acetate buffer (pH 4.5), and 250 µL of 4 mM p-nitrophenyl β-D-glucopyranoside (pNPβG, Sigma), incubated for 10 min at 50°C. The enzymatic reaction was stopped by addition of 2 mL of 2 M sodium carbonate, and the liberated product was spectrophotometrically quantified at 410 nm using a 4-nitrophenol standard curve. One unit of β-glucosidase was defined as the amount of enzyme that releases 1 µmol of nitrophenol per minute of reaction (Palma-Fernandez et al., 2002).
Protein determination
Protein content was measured, following the method of Hartree (1972), using bovine serum albumin (BSA) as standard.
Biochemical characterization of β-glucosidase
Effect of pH and temperature
The optimum pH was determined by measuring the enzyme activity in 0.1 M citrate-phosphate buffer solution at 50°C and pH values of 3.0-8.0. The optimum temperature was determined by measuring the enzyme activity at temperatures from 35 to 70°C, at the optimal pH of the enzyme. The pH stability was determined by incubating the enzyme for 24 h at 25°C at different pH values (3.0-10.5). The buffers used (0.1 M) were McIlvine buffer (pH 3.0-8.0), Tris-HCl (pH 8.0-8.5) and Glycine-NaOH (pH 8.5-10.5). Thermostability was studied by incubating the enzyme for 1 h at different temperatures (35-70°C) at McIlvine buffer pH 4,5. After incubation, aliquots were withdrawn and cooled on ice bath. Residual activities were measured under optimal conditions of the enzyme.
Effect of ethanol and glucose on β-glucosidase activity
Enzymatic activities were quantified with the addition of glucose or ethanol at different concentrations in the reaction mixture (0-35 mM glucose or 0-30% ethanol). Assays were performed under optimal conditions of pH and temperature.
Catalytic potential of the enzymatic extract
Carboxymethylcellulase (CMCase) and xylanase activities were determined by adding 100 µL of enzyme extract in 900 µL of 0.1 M sodium acetate buffer (pH 4.5) containing 3% carboxymethyl-cellulose (Sigma C5678) and 0.5% xylan (Sigma Birch -Wood), respectively. After 10 min of reaction, the reducing sugar released was quantified by the DNS method (3,5-dinitrosalicylic acid) (Miller, 1959) using glucose or xylose standard curve, respectively. β-Xylosidase activity was measured with the synthetic substrate p-nitrophenyl β-D-xylopyranoside (4 mM; Sigma), following the methodology previously described for β-glucosidase. Assays were performed in 0.1 M sodium acetate buffer (pH 4.5) at 50°C. One unit of enzymatic activity was defined as the amount of enzyme capable of producing 1 µmol of product per minute of reaction.
RESULTS AND DISCUSSION
β-Glucosidase production by solid-state fermentation
Different agro-industrial residues were tested for β-glucosidase production by the fungus G. butleri, for which other fermentation parameters were fixed, including moisture (60%), temperature (28°C) and cultivation time (96 h). The highest enzyme production (213.8 U/g dry substrate) and specific activity (6.39 U/mg of protein) was obtained using wheat bran as substrate. Other residues evaluated in this study showed no significant production when compared with wheat bran (Table 1). A previous study confirmed the potential for β-glucosidase production by microorganisms of the genus Gongronella, when grown in wheat bran (Fang et al., 2014).
The increased production of the enzyme on wheat bran can be attributed to the fact that this residue is rich in proteins, cellulose, inorganic salts, nitrogen and carbon. Wheat bran acts as a source of nutrients and support for mycelial adhesion of the filamentous fungi, being similar to their natural habitat (Abdella et al., 2014). The nutritional complexity of this residue favors the growth of many microorganisms, and production of several enzymes of industrial interest (Haque et al., 2002). Considering the results shown in Table 1, wheat bran was selected as substrate for subsequent optimization of parameters that affect enzymatic production.
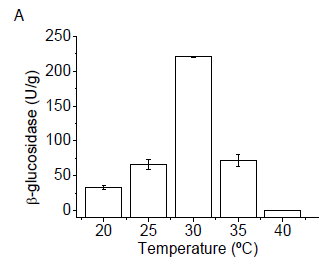
Among the evaluated temperatures, the optimal production of β-glucosidase by the fungus G. butleri was obtained in cultivations at 30°C, reaching about 211.7 U/g (Figure 1A). The optimum temperature for enzyme production does not always correspond to the temperature of the natural habitat of the microorganism (Kheng and Omar, 2005). High or very low temperatures can slow down the growth of a microorganism, and thereby the formation of product. The low thermal conductivity of agro-industrial residues used in solid-state fermentation processes can hinder the dissipation of metabolic heat generated by microbial growth (Pandey et al., 2003). Thus, analysis of the cultivation temperature is essential to delineate a bioprocess to produce enzymes, considering that the biological catalysts are denatured at high temperatures.
The fungus G. butleri was also grown on wheat bran with different moisture levels. As seen in Figure 1B, the optimal production of β-glucosidase was obtained in cultivations containing 55 or 60% initial moisture (w/v). In cultures containing 80% moisture, enzyme production by the microorganism was near zero (Figure 1B), indicating that higher levels of moisture were unfavorable for enzyme production.
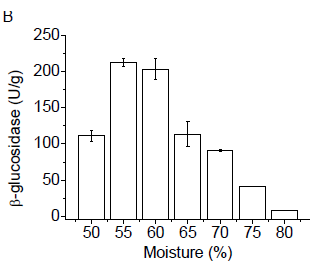
Moisture content has key role in the success of solid-state fermentation processes, and may vary according to the substrate and the microorganism used (Kalogeris et al., 2003). The moisture of the medium must contribute to the growth of the microorganism, which does not occur at very low levels. However, excess moisture reduces the porosity of the medium and hampers gas exchange, resulting in the reduction of microbial activity and conse-quently the production of enzymes (Bansal et al., 2012).
Cultivation time was the last parameter evaluated. Production of β-glucosidase was highest at 96 h of cultivation, reaching about 215.4 U/g dry substrate (or 21.5 U/mL). After this period, enzymatic activity fell considerably (Figure 1C); the decline in enzyme production after 120 h of cultivation can be related to the reduced amount of nutrients and metabolic changes of fungus, resulting in the denaturation of enzymes (Bon et al., 2008).
Higher enzyme production was observed with only 96 h of cultivation, confirming the reduced time to obtain enzymes of microbial origin, when compared with plant and animal enzymes, which highlight the trend of the use of microbial enzymes in industrial processes (Oliveira et al., 2006). When compared with other fungal strains, the optimum cultivation time for the production of β-glucosidase by the fungus G. butleri is considerably lower. For example, Pereira et al. (2015b) reported maximum enzyme production after 192 h of solid-state cultivation of the fungus Thermomucor indicae-seudaticae N31. Leite et al. (2008) had higher production of β-glucosidase at 120 h of solid-state cultivation by the yeast Aureobasidium pullulans.
Production of β-glucosidase by the fungus G. butleri was extremely high, about 215.4 U/g of dry substrate, as compared to previously reported data (Table 2). Considering the low incidence of studies using this fungal species for the production of industrial enzymes, the β-glucosidase produced by G. butleri was biochemically characterized and, the catalytic profile of enzymatic extract was valued.
Biochemical characterization of β-glucosidase
Effect of pH and temperature
β-Glucosidase produced by the fungus G. butleri showed optimal activity at pH 4.5 and 65°C (Figure 2A and B). However, the half-life (t1/2) of the enzyme was approximately 36 and 12 min., at 60 and 65° C, respectively (Figure 2E), suggesting the enzyme employment at 60°C. The enzyme retained its original activity after 24 h of incubation over a pH range of 3.0-7.5 (Figure 2C). After incubation for 1 h at 50°C, about 80% of the catalytic potential was recovered. When incubated at 55°C for the same period, the enzyme maintained57% of its original activity (Figure 2D).
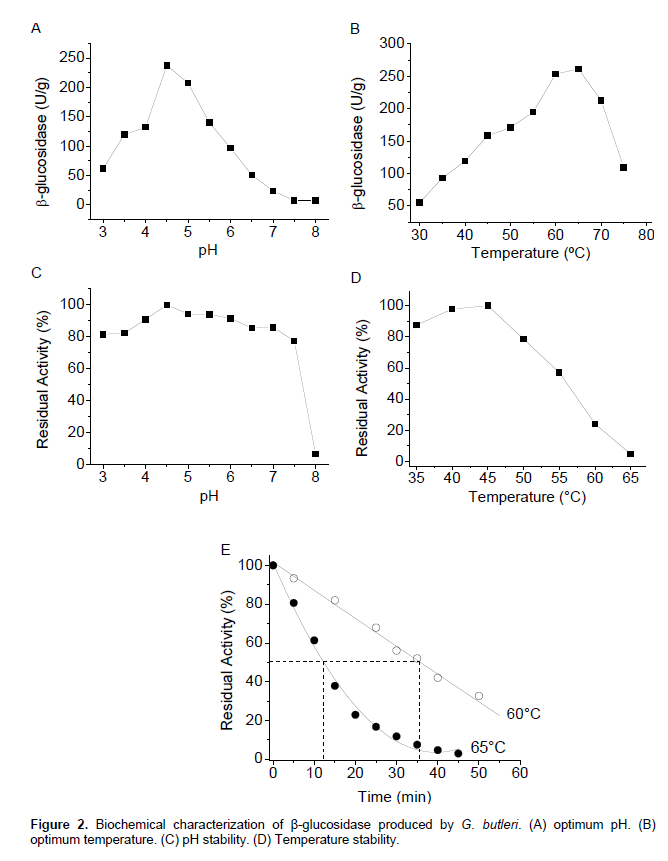
The characteristics of the β-glucosidase produced by the fungus G. butleri are similar to those described in the literature for β-glucosidase produced by mesophilic and thermophilic fungi. Leite et al. (2008) reported optimal pH values of 4.5 and 4.0 for β-glucosidase produced by the fungi A. pullulans and Thermoascus aurantiacus, respectively. Lin et al. (1999) obtained the same optimum temperature for β-glucosidase produced by the fungus Thermomyces lanuginosus SSBP. Garcia et al. (2015) reported optimal activity at pH 5.5 and 65°C for β-glucosidase produced by the fungus Lichtheimia ramosa; further, the enzyme retained its catalytic activity after 1 h at 55°C. Camassola et al. (2004) showed that the highest activity of β-glucosidase produced by Penicillium echinulatum occurred between 55 and 60°C. Yun et al. (2001) reported the production of extracellular β-glucosidase by the fungus Trichoderma harzianum, and demonstrated that the enzyme was stable at temperatures below 55°C for 15 min, keeping only 36% of the initial activity after 15 min at 60°C. Delabona et al. (2013) described the stability of β-glucosidase produced by the fungus Aspergillus fumigatus P40M2 at temperatures of 40-60°C and pH levels 3.0-5.5. The β-glucosidase from Penicillium funiculosum NCL1 showed optimum activity at pH 4.0 and 5.0, optimum temperature of 60°C, and exhibited a half-life of 1 h at 60°C (Ramani et al., 2012).
Effect of ethanol and glucose on β-glucosidase activity
As shown in Figure 3A, ethanol concentrations near 5% potentiated the enzymatic activity as compared to the control, increasing levels to approximately 40%. The enzyme showed catalytic activity similar to the control in solutions containing 15% ethanol, yet when the ethanol concentration was increased to 25%, only 62% of the original enzymatic activity was recovered (Figure 3A).
β-Glucosidases can be applied in industrial processes that contain alcohol (e.g., production of biofuels and beverages), which makes ethanol tolerance an extremely valuable feature for this group of enzymes (Sun and Cheng, 2002). The characteristics described in this study allow us to infer that the β-glucosidase produced by G. butleri can be applied in processes which have alcoholic fermentation as their central metabolic route, considering that the final ethanol concentration in fermented broths obtained by traditional processes is about 10%. Alcoholic concentrations greater than this are generally harmful to the fermenters organisms (Gu et al., 2001).
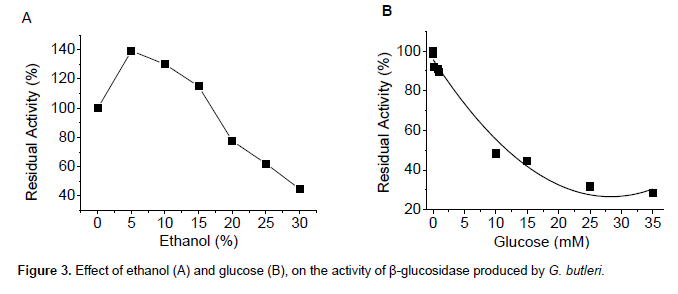
The enzyme was strongly inhibited by glucose, showing only 50% of its catalytic activity in solutions containing 10 mM glucose (Figure 3B). However, when the concentration of substrate (pNPβG) was increased to the same concentration of the inhibitor (glucose), inhibition of the enzyme was completely reversed (Table 3), indicating that the inhibitor and the substrate compete for the same binding site, typical characteristic of competitive inhibition (Leite et al., 2008).

Most microbial β-glucosidases are inhibited by glucose, which may be competitive or non-competitive (Leite et al, 2008; Sonia et al., 2008). Competitive inhibition can be controlled by periodic addition of substrate to keep it at a concentration exceeding the levels of the inhibitor (product of the enzymatic action); alternatively, the product formed by the action of the enzyme can be removed to maintain it at concentrations below the substrate (Bon et al., 2008; Garcia et al., 2015). The characteristics described for the β-glucosidase of fungus G. butleri (ethanol tolerance and competitive inhibition of glucose) enable its application in simultaneous saccharification and fermentation processes, with the aim of obtaining ethanol from vegetal biomass. In this type of process, the glucose liberated by enzymatic hydrolysis of cellulose is simultaneously converted to ethanol by microbial fermentation, minimizing inhibition of the enzymes of the cellulolytic complex (Scott et al., 2013).
Catalytic potential of the enzymatic extract
The catalytic potential of enzymatic extract obtained under optimal conditions of cultivation was evaluated. Among the enzymes of the cellulolytic and hemicellulolytic complexes that were evaluated, high xylanase production by the fungus G. butleri was noted, reaching about 4303 U/g dry substrate (or 430.3 U/mL) (Table 4). The xylanase values obtained in this study are considerably high when compared with those previously reported. For example, Maciel et al. (2008) reported the production of xylanaseby the fungus A. niger LPB 326 in solid-state fermentation, to be about 3099 IU/g of substrate. Rezende et al. (2002) obtained 288 U/mL of xylanase by solid-state fermentation of the fungus T. harzianum in sugar cane bagasse and Da-Silva et al. (2005) produced 107 U/mL xylanase by the fungus T. aurantiacus using corncob as substrate. Sadaf and Khare (2014) reported xylanase production of 1025 U/g of substrate by the fungus Sporotrichum thermophile by solid-state fermentation.
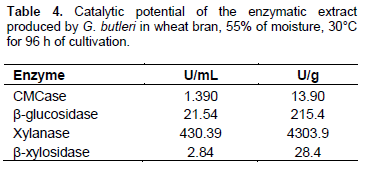
Significant values for CMCase and β-xylosidase were not obtained in optimized cultivation conditions (Table 4). The catalytic profile of enzymatic extract described in this work is extremely desirable for application within the cellulose and paper industries. Enzymatic extract with high concentrations of xylanase and reduced cellulase activity can be used for bleaching Kraft pulp, in order to obtain white paper. The presence of cellulases can reduce the quality of paper because they degrade cellulose fibers. Hydrolysis of xylan facilitates the removal of residual lignin from pulp, thereby reducing the use of conventional chemical agents that contribute to the environmental damage caused by the effluents of the paper industry (Xin and He, 2013).
CONCLUSIONS
The results obtained in this study highlight the fungus G. butleri as an excellent β-glucosidase producer, especially when cultivated on wheat bran. The characteristics of this β-glucosidase enable the application of this enzyme in different industrial processes, such as hydrolysis of cellulose for production of second-generation ethanol, or in the food and beverage industry. The enzymatic extracts obtained under optimal growing conditions showed high xylanase activity with reduced cellulolytic potential, making them highly applicable for improvement of cellulose pulp bleaching processes.
CONFLICT OF INTEREST
The authors declare that there is no conflict of interests.
ACKNOWLEDGEMENTS
The authors gratefully acknowledge the financial support of the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Apoio ao Desenvolvimento do Ensino, Ciência e Tecnologia do Estado de Mato Grosso do Sul (FUNDECT) and Coordenação de Aperfeiçoamento Pessoal de Nível Superior (CAPES).
REFERENCES
|
Abdella A, Mazeed TES, Yang ST, El-Baz, AF (2014). Production of β-Glucosidase by Aspergillus niger on Wheat Bran and Glycerol in Submerged Culture: Factorial Experimental Design and Process Optimization. Curr. Biotechnol. 3(2):197-206. |
|
|
Alves-Prado HF, Leite RSR, Bocchini DA, Gomes E, Da-Silva R (2011). Cellulolytic enzymes isolated from Brazilian areas: production, characterization and applications. In: Adam E. Golan. (Org.). Cellulase: Types and Action, Mechanism and Uses. 1ed.Hauppauge, NY: Nova Science Publishers, Cap 6:178-206. |
|
|
Bansal N, Tewari R, Soni R, Soni SK (2012). Production of cellulases from Aspergillus niger NS-2 in solid state fermentation on agricultural and kitchen waste residues. Waste Manage. 32:1341-1346. |
|
|
Bon EPS, Gírio F, Pereira-Junior N (2008). Enzimas na Produção de Etanol. Enzimas em Biotecnologia: Produção, Aplicações e Mercado. Rio de Janeiro: Interciência, pp. 241-272. |
|
|
Borges DG, Baraldo-Junior A, Farinas CS, Giordano RLC, Tardioli PW (2014). Enhanced saccharification of sugarcane bagasse using soluble cellulase supplemented with immobilized b-glucosidase. Bioresour. Technol. 167:206-213. |
|
|
Camassola M, Bittencourt LR, Shenem NT, Andreaus J, Dillon AJP (2004). Characterization of the cellulase complex of Penicillium echinulatum. Biocatal. Biotransformation 22:391-396. |
|
|
Da-Silva R, Lago ES, Merheb CW, Macchione MM, Park YK, Gomes E (2005). Production of xylanase and CMCase on solid state fermentation in different residues by Thermoascus aurantiacus miehe. Braz. J. Microbiol. 36(3):235-241. |
|
|
Delabona PDS, Pirota RDPB, Codima CA, Tremacoldi CR, Rodrigues A, Farinas CS (2013). Effect of initial moisture content on two Amazon rainforest Aspergillus strains cultivated on agro-industrial residues: Biomass-degrading enzymes production and characterization. Ind. Crop. Prod. 42:236-242. |
|
|
Deswal D, Khasa YP, Kuhad RC (2011). Optimization of cellulose production by a brown rot fungus Fomitopsis sp. RCK2010 under solid state fermentation. Bioresour. Technol. 102(10):6065-6072. |
|
|
Fang W, Song R, Zhang X, Zhang X, Zhang X, Wang X, Fang Z, Xiao Y (2014). Characterization of a novel β-glucosidase from Gongronella sp. W5 and its application in the hydrolysis of soybean isoflavone glycosides. J. Agric. Food Chem. 62(48):11688-11695. |
|
|
Gao J, Weng H, Zhu D, Yuan M, Guan F, Xi Y (2008). Production and characterization of cellulolytic enzymes from the thermoacidophilic fungal Aspergillus terreus M11 under solid-state cultivation of corn stover. Bioresour. Technol. 99(16):7623-7629. |
|
|
Garcia NFL, Santos FRS, Gonçalves FA, Paz MF, Fonseca GG, Leite RSR (2015). Production of β-glucosidase on solid state fermentation by Lichtheimia ramosa in agroindustrial waste: characterization and catalytic properties of the enzyme extract. Electron. J. Biotechnol. 18(4):314-319. |
|
|
Gonçalves FA, Leite RSR, Rodrigues A, Argando-a EJS, Fonseca GG (2013). Isolation, identification and characterization of a novel high level β-glucosidase-producing Lichtheimia ramosa strain. Biocatal. Agric. Biotechnol. 2(4):377-384. |
|
|
Gu Y, Qiao M, Zhou Q, Zhou Z, Chen G (2001). Hyperproduction of Alcohol Using Yeast Fermentation in Highly Concentrated Molasses Medium. Tsinghua Sci. Technol. 6(3):225-230. |
|
|
Haque MA, Shams-Ud-Din M.; Haque, A (2002). The effect of aqueous extracted wheat bran on the baking quality of biscuit. Int. J. Food Sci. Technol. 37(4):453-462. |
|
|
Hartree EF (1972). Determination of protein: a modification of the Lowry method that gives a linear photometric response. Anal. Biochem. 48(2):422-427. |
|
|
Huber GW, Iborra S, Corma A (2006). Synthesis of Transportation Fuels from Biomass: Chemistry, Catalysts, and Engineering. Chem. Rev. 106(9):4044-4098. |
|
|
Kalogeris E, Iniotaki F, Topakas E, Christakopoulos P, Kekos D, Macris BJ (2003). Performance of an intermittent agitation rotating drum type bioreactor for solid-state fermentation of wheat straw. Bioresour. Technol. 86(3):207-213. |
|
|
Kang SW, Park YS, Lee JS, Hong SI, Kim SW (2004). Production of cellulases and hemicellulases by Aspergillus niger KK2 from lignocellulosic biomass. Bioresour. Technol. 9(2):153-156. |
|
|
Kheng PP, Omar IC (2005). Xylanase production by a local fungal isolate, Aspergillus niger USM AI 1 via solid state fermentation using palm kernel cake (PKC) as substrate. Songklanakarin J. Sci. Technol. 27(2):325-336. |
|
|
Leite RSR, Alves-Prado HF, Cabral H, Pagnocca FC, Gomes E, Da-Silva R (2008). Production and characteristics comparison of crude β-glucosidases produced by microorganisms Thermoascus aurantiacus e Aureobasidium pullulans in agricultural wastes. Enzyme Microb. Technol. 43(6):391-395. |
|
|
Leite RSR, Bocchini DA, Martins ES, Silva D, Gomes E, Da-Silva R (2007). Production of cellulolytic and hemicellulolytic enzymes from Aureobasidium pulluans on solid state fermentation. Appl. Biochem. Biotechnol. 137(1):281-288. |
|
|
Lin J, Pillay B, Singh S (1999). Purification and biochemical characteristics of beta-D-xylanase from a thermophilic fungus, Thermomyces lanuginosus - SSBP. Biotechnol. Appl. Biochem. 30:81-87. |
|
|
Maciel GM, Vandenberghe LPS, Haminiuk CWI, Fendrich RC, Della-Bianca BE, Brandalize TQS, Pandey A, Soccol CR (2008). Xylanase production by Aspergillus niger LPB 326 in solid-state fermentation using statistical experimental designs. Food Technol. Biotechnol. 46(2):183-189. |
|
|
Merheb-Dini C, Cabral H, Leite RSR, Zanphorlin LM, Okamoto DN, Rodriguez GOB, Juliano L, Arantes EC, Gomes E, Da-Silva R (2009). Biochemical and functional characterization of a metalloprotease from the thermophilic fungus Thermoascus aurantiacus. J. Agric. Food Chem. 57(19):9210-9217. |
|
|
Miller GL (1959). Use of Dinitrosalicylic Acid Reagent for Determination of Reducing Sugar. Anal. Chem. 31(3):426-428. |
|
|
Oliveira APA, Silvestre MA, Alves-Prado HF, Rodrigues A, Paz MF, Fonseca GG, Leite RSR (2015). Bioprospecting of yeasts for amylase production in solid state fermentation and evaluation of the catalytic properties of enzymatic extracts. Afr. J. Biotechnol. 14(14):1215-1223. |
|
|
Oliveira NA, Oliveira LA, Andrade JS, Chagas-Júnior AF (2006). Enzimas hidrolíticas extracelulares de isolados de Rizóbia nativos da Amazônia Central, Amazonas, Brasil. Ciênc. Tecnol. Aliment. 26(4):853-860. |
|
|
Palma-Fernandez ERD, Gomes E, Da-Silva R (2002). Purification and Characterization of two β-Glucosidases from Thermophilic Fungus Thermoascus aurantiacus. Folia Microbiol. 47(6):685-690. |
|
|
Pandey A (2003). Solid-State Fermentation. Biochem. Eng. J. 13:81-84. |
|
|
Pauly M, Keegstra K (2010). Plant cell wall polymers as precursors for biofuels. Curr. Opin. Plant Biol. 13(3):304-311. |
|
|
Pereira JC, Leite RSR, Alves-Prado HF, Bocchini-Martins DA, Gomes E, Da-Silva R (2015b). Production and characterization of β-glucosidase obtained by the solid-state cultivation of the thermophilic fungus Thermomucor-indicae seudaticae N31. Appl. Biochem. Biotechnol. 175(2):723-732. |
|
|
Pereira JC, Marques NP, Rodrigues A, Oliveira TB, Boscolo M, Da-Silva R, Gomes E, Bocchini-Martins DA (2015a). Thermophilic fungi as new sources for production of cellulases and xylanases with potential use in sugarcane bagasse saccharification. J. Appl. Microbiol. 118(4):928-939. |
|
|
Ramani G, Meera B, Vanitha C, Rao M, Gunasekaran P (2012). Production, purification,and characterization of a β-glucosidase of Penicillium funiculosum NCL1. Appl. Biochem. Biotechnol. 167(5):959-972. |
|
|
Rani V, Mohanram S, Tiwari R, Nain L, Arora A (2014). Beta-Glucosidase: Key Enzyme in Determining Efficiency of Cellulase and Biomass Hydrolysis. J. Bioprocess Biotech. 5:197. |
|
|
Rezende MI, Barbosa AM, Vasconcelos AFD, Endo AS (2002). Xylanase production by Trichoderma harzianum rifai by solid state fermentation on sugarcane bagasse. Braz. J. Microbiol. 33:67-72. |
|
|
Romero E, Bautista J, García-Martínez AM, Cremades O, Parrado J (2007). Bioconversion of corn distiller's dried grains with solubles (CDDGS) to extracellular proteases and peptones. Process Biochem. 42(11):1492-1497. |
|
|
Sadaf A, Khare SK (2014). Production of Sporotrichum thermophile xylanase by solid state fermentation utilizing deoiled Jatropha curcas seed cake and its application in xylooligosachharide synthesis. Bioresour. Technol. 153:126-130. |
|
|
Scott F, Quintero J, Morales M, Conejeros R, Cardona C, Aroca G (2013). Process design and sustainability in the production of bioethanol from lignocellulosic materials. Electron. J. Biotechnol. 16(3):1-16. |
|
|
Silva CAA, Lacerda MPF, Leite RSR, Fonseca GG (2013). Production of enzymes from Lichtheimia ramosa using Brazilian savannah fruit wastes as substrate on solid state bioprocesses. Electron. J. Biotechnol. 16(5):1-9. |
|
|
Singhania RR, Patel AK, Sukumaran RK, Larroche C, Pandey A (2013). Role and significance of beta-glucosidases in the hydrolysis of cellulose for bioethanol production. Bioresour. Technol. 127:500-507. |
|
|
Sonia KG, Chadha BS, Badhan AK, Saini HS, Bhat MK (2008). Identification of glucose tolerant acid active β-glucosidases from thermophilic and thermotolerant fungi. World J. Microbiol. Biotechnol. 24:599-604. |
|
|
Sun Y, Cheng J (2002). Hydrolysis of lignocellulosic materials for ethanol production: a review. Bioresour. Technol. 83:1-11. |
|
|
Thakur VK, Thakur MK (2014). Processing and characterization of natural cellulose fibers/thermoset polymer composites. Carbohydr. Polym. 109:102-117. |
|
|
Xin F, Geng A (2010). Horticultural Waste as the Substrate for Cellulase and Hemicellulase Production by Trichoderma reesei Under Solid-State Fermentation. Appl. Biochem. Biotechnol. 162:295-306. |
|
|
Xin F, He J (2013). Characterization of a thermostable xylanase from a newly isolated Kluyvera species and its application for biobutanol production. Bioresour. Technol. 135:309-315. |
|
|
Yun SI, Jeong CS, Chung DK, Choi HS (2001). Purification and some properties of a β-Glucosidase form Trichoderma harzianum type C-4. Biosci. Biotechnol. Biochem. 65:2028-2032. |
|
Copyright © 2024 Author(s) retain the copyright of this article.
This article is published under the terms of the Creative Commons Attribution License 4.0


